Emelt szintű biológia érettségi előkészítő tanfolyamok
Tisztelt Érdeklődők!
A 2026. évi október-novemberi, ill. a 2027. évi május-júniusi érettségi vizsgákra felkészítő tanfolyamokra április 15-től nyílik lehetőség jelentkezni.
Jelentkezni e-mailben lehet: bioszfera.com@gmail.com
2026. évi október-novemberi emelt szintű biológia érettségi vizsgára felkészítő nyári online tanfolyam
A képzés júliusban és augusztusban heti 2x2 órában történik, hétfőn és csütörtökön 10:00-11:30-ig, szeptembertől pedig az órák szombat délelőtt lesznek 10:00-11:30-ig.
A tanfolyam kezdési időpontja június 29.
Az óradíj 1600 Ft.
Jelentkezési lap letöltése (pdf)
Részletes tájékoztató letöltése (pdf)
A 2027. évi május-júniusi emelt szintű biológia érettségi vizsgára felkészítő jelenléti tanfolyam
A képzés összesen 120 óra, 30 héten keresztül tart, heti 2x2 órában, a tantermi órák kedden és csütörtökön 16:30-18:00-ig lesznek a Városmajori Gimnáziumban.
A tanfolyam kezdési időpontja szeptember 8.
Az óradíj 1400 Ft.
Jelentkezési lap letöltése (pdf)
Részletes tájékoztató letöltése (pdf)
A 2027. évi május-júniusi emelt szintű biológia érettségi vizsgára felkészítő online tanfolyam
A képzés összesen 120 óra, 30 héten keresztül tart, heti 2x2 órában, az órák szerdán és pénteken 16:30-18:00-ig lesznek.
A tanfolyam kezdési időpontja szeptember 9.
Az óradíj 1400 Ft.
Az interaktív online képzés mind formailag, mind tematikáját tekintve megegyezik a jelenléti oktatással.
Jelentkezési lap letöltése (pdf)
Részletes tájékoztató letöltése (pdf)
Emelt szintű feladatmegoldó online tanfolyam
A képzés összesen 56 óra, 28 héten keresztül tart, heti 1x2 órában, hétfőnként 17:30-19:00-ig.
A tanfolyam kezdési időpontja szeptember 28.
Az óradíj 1500 Ft.
Jelentkezési lap letöltése (pdf)
Részletes tájékoztató letöltése (pdf)
A Voovo az előkészítő tanfolyami órákkal együtt minden tudást megadnak, ami a sikeres emelt szintű biológia érettségihez kell.
Gyere az előkészítő órákra és hozd ki a maximumot a Voovo appal.
A felkészítés garanciája a tanfolyamot vezető tanár 34 éves tanítási és sikeres érettségire felkészítő tapasztalata, illetve vizsgáztatói gyakorlata. A tanfolyamot vezető tanár Vizkievicz András, aki a kétszintű érettségi bevezetése óta, 20 éve folyamatosan ellát
- írásbelivel kapcsolatosan javító szaktanári,
- javításvezetői,
- felüljavítói,
- szóbelin kérdező szaktanári,
- vizsgaelnöki feladatokat,
- továbbá társszerzője az Oktatási Hivatal által kiadott, Gyűjtemény a Biológia emelt szintű oktatásához tankönyvnek.
- A korábbi érettségi rendszerben pedig a felvételi feladatsorok készítésében közreműködött, mint feladatkészítő tanár.
1991-ben végeztem az ELTE TTK biológia-kémia szakán. Az egyetem negyedik és ötödik évében az egyetem Biokémia Tanszékén géntechnológia témakörében dolgoztam, s szakdolgozatomat is ebből a témából írtam. 1991 óta első munkahelyemen, a Városmajori Gimnáziumban dolgozom.
2011-ben a Budapest XII. kerületi Hegyvidéki Önkormányzat a szakmai tevékenységem elismeréseként a Hegyvidék Ifjúságáért Díjat adományozta.
2018-ban az Emberi Erőforrások Minisztériuma a "kiemelkedően eredményes és áldozatos tehetségsegítő munka elismerésképpen" a "Bonis Bona - A Nemzet Tehetségeiért" díjat adományozta.
Az előterjesztésben az alábbiak szerepeltek:
A természettudományok megszerettetése, a biológiai kutatások iránti fogékonyság megteremtése a specialitása. Tanítványainak többsége az orvosi pályán vagy kutatóként dolgozik, valamint a természettudományos szakterületeken is doktori fokozatot szerzők többsége az ő tanítványai. Az iskolát a Semmelweis Egyetem a Tanár úr munkássága alapján kérte fel partnerintézményi együttműködésre. A sikeres magas számú természettudományos pályára való felkészítés mellett, versenyeredményekkel is büszkélkedhetnek diákjai.
A természettudományos tantárgyak mindenki által elérhetővé tételére álmodta meg a bioszféra digitális tananyagot, melynek fejlesztésben tehetséges diákjaival közösen alkotnak, s amely az iskola honlapjáról elérhető. A portál folyamatos fejlesztés alatt van, így egyre több a többi tanuló számára is letölthető tanulmányi információk száma. Tanárideálként is kiemelkedő egyéniség, aki hihetetlen népszerűsége, végtelen sok energiával végzett munkája mellett szerény, kedves tanárember tud maradni.
Vizkievicz András 30 éve a Városmajori Gimnáziumban tanít biológiát és kémiát. Az iskola biológia tagozatos tantervi programjának meghatározó szakmai kidolgozója. A biológia tagozatra jelentkező tanulók létszáma az ő munkássága alapján iskolánk kiemelt, 10-szeres túljelentkezéssel működő képzési iránya. Éveken keresztül munkaközösség vezetőként segítette a képzés hatékonyságát, de a SE felvételi, közép és emelt szintű érettségi felkészítő tanáraként is kiemelkedő eredményeket mondhat magáénak. Az iskolában ő alakította ki a biológia emelt szintre felkészítő fakultáció rendszerét, szakterületén tapasztalható végtelen precizitása, a tantárgy iránti elkötelezettsége, a megbízhatósága, a szakmai feladatteljesítési szerénysége és tisztessége pótolhatatlanná teszi őt. Varázsos egyénisége miatt a diákok a biológia emelt szintű képzésre vagy fakultációs képzésre is szinte csak az ő neve miatt jelentkeznek. Óráinak összeállítása, módszertana, valamint a szakma iránt érzett elkötelezettsége példaértékű.
2025-ben a Közép-Budai Tankerületi Központ a Tankerület Kiváló Pedagógusa címben részesített.
Mit nyújt az előkészítő tanfolyam számodra?
Az előkészítő tematikája, metodikája hosszú évek – több, mint 3 évtizedes tanítási, felkészítési, továbbá vizsgáztatói, kérdező tanári, javítói, felüljavítói, javításvezetői, vizsgaelnöki gyakorlat - során kristályosodott ki.
Hangsúlyoznom kell, hogy a tanfolyamon való részvétel nem helyettesíti a lexikális anyag megtanulását, a rendszeres otthoni készülést, a feladatok gyakorlását.
Miután Winston Churchillt a második világháború második évében, 1940 májusában miniszterelnökké választották, ezekkel a cseppet sem megnyugtató szavakkal szólt a tömegekhez: "Nem ígérhetek mást, csak vért, erőfeszítést, verítéket és könnyeket!"
Továbbá,” ez nem a könnyed kényelem ideje. Ez a merészség és kitartás ideje.”
Valahogy így van ez az érettségire készüléssel is.
Az előkészítő utat mutat, irányt ad, a tanfolyam szeptembertől egészen április közepéig rendszerezi a tanulásod, a tananyag beosztása, egymásra épülése segíti az egyes anyagrészek megértését, megvilágítja az egyes fejezetek kapcsolatát. A tananyag feldolgozása a bioszfera.com oldalon, a NAT 2024 emelt szintű jegyzetek menüben található tematika alapján történik. A tanfolyam anyaga teljes mértékben illeszkedik az érettségi követelményrendszerhez.
A sikeres érettségi vizsgának minimum 3 feltétele van:
- a rendszeres részvétel az órákon,
- folyamatos, lelkiismeretes készülés,
- és az elsajátított elmélet alkalmazása a feladatmegoldások során, más szóval a gyakorlás.
- az ismeretek elsajátítása,
- az elsajátított ismeret megértése,
- az elsajátított és megértett ismeretek alkalmazása problémák megoldásában.
- megtanuljuk az emelt szintű követelményrendszer által meghatározott elméleti tananyagot,
- hozzávetőleg 100 érettségi feladatot oldunk meg,
- rendszeresen ellenőrizzük a felkészülés eredményességét,
- 2 alkalommal próbaérettségi megírására kerül sor,
- megvilágítjuk az ok-okozati összefüggéseket,
- felfedezzük a különböző biológiai jelenségek közötti kapcsolatokat,
- elemzünk szövegeket, ábrákat, adatokat, grafikonokat, biológiai kísérleteket,
- értelmezések alapján következtetéseket fogalmazunk meg,
- fejlesztjük a természettudományos gondolkodást, logikát,
- elsajátítjuk annak a képességét, hogy a vizsgázó önállóan és logikus gondolatmenetbe illesztve, összefüggően és a nyelvhelyesség szabályainak megfelelően legyen képes megfogalmazni mondanivalóját.
Az elmélyült tudásnak 3 szintje van:
A biológia tananyag feldolgozása a bioszfera.com oldalon, a Emelt szintű jegyzetek menüben található fejezetek alapján történik. A tanítás célja az érettségi követelményrendszerben megfogalmazott kompetenciák elsajátítása.
-
Az órákon:
A 120 órás felkészítő tanfolyamon át kell vennünk a teljes biológia tananyagot emelt szinten, amit egyébként a középiskolai biológia oktatás során 4 éven át, összesen minimum 342 órában tanultatok. Ezért az idő rövidsége miatt a megértést nem igénylő, kizárólag lexikális ismereteket magába foglaló fejezeteket mellőzzük, pl. állattan, etológia, bőr, ezen témakörök tartalmi részének az elsajátítása egyéni feladat. Ugyanakkor különös hangsúlyt kapnak a megértést igénylő, bonyolultabb összefüggéseket tartalmazó fejezetek, mint pl. emberi szervezettan, élettan, biokémia, sejtbiológia, genetika, populációgenetika, molekuláris genetika, biotechnológia, ökológia stb.
Az órák menete
A 120 órás képzés hetente kétszer történik 30 héten keresztül, 60 alkalommal, összesen 120 órában. Egy óra időtartama 45 perc, egy alkalommal 2 óra lesz, összesen 90 percben. Az órák 16:30-kor kezdődnek, de már 16 órától meg lehet kezdeni az óra előtti feladat kidolgozását.
Az óra elején az általatok feltett kérdésekre válaszolok, majd az adott tananyag elméletét vesszük át. Az óra egy másik részében pedig az adott anyagrésszel kapcsolatos problémás érettségi feladatokat beszéljük meg. Az év során, az órákon több, mint 100 emelt szintű érettségi feladatot oldunk meg.
Minden óra előtt (nem kötelező jelleggel) – 60 alkalommal - az aktuális tananyaggal kapcsolatban, ellenőrzésképpen az előző órák anyagából feladatot oldunk meg egyénileg, melynek %-os eredményét feljegyezzük, így minden témakörből lesz mérés, értékelés, melynek összesített eredménye a sok éves tapasztalat alapján előrevetíti az írásbeli vizsga várható eredményét. Ezen túlmenően az év során 2 próbaérettségit írunk.
Fontos az órákra való folyamatos készülés. Ehhez segítséget adnak a bioszfera.com-on elérhető Emelt szintű jegyzetek, melyek tartalma teljes mértékben az érettségi követelményrendszerre lett igazítva, persze azon túlmutatva.
A tanfolyam költsége
A tanfolyamon való részvétel a tanfolyami díj befizetésével biztosított.
A képzés költségét a következő számlaszámra kell utalni: 50420898-10004902
A befizetést igazoló számlát kata-s vállalkozóként a résztvevő diák nevére állítom ki.
120 órás tanfolyam
Az óradíj 1400 Ft. A tanfolyami díj rendezése történhet egy összegben, de van lehetőség negyedéves, ill. féléves részletfizetésre is.
A tanfolyam teljes költsége 120x1400 Ft, azaz 168 000 Ft.
Az első három negyedév költsége, negyedévenként 39 200 Ft (28x1400 Ft), a kicsit hosszabb negyedik negyedév díja 50 400 Ft (36x1400 Ft).
Fizetési ütemezés
A negyedévek befejeztével, legkésőbb:
2. negyedév, október 31-ig,
3. negyedév, 2026 január 5-ig,
4. negyedév, 2026 február 20-ig.
50 órás nyári tanfolyam
Az óradíj 1600 Ft. A tanfolyam teljes költsége 50x1600 Ft, azaz 80 000 Ft.
Feladatmegoldó tanfolyam
Az óradíj 1500 Ft. A tanfolyami díj rendezése történhet egy összegben, de van lehetőség féléves részletfizetésre is.
A tanfolyam teljes költsége 56x1500 Ft, azaz 84 000 Ft.
Érdeklődni lehet Vizkievicz András vezető tanárnál, email-ben, telefonon vagy személyesen a Városmajori Gimnáziumban.
A felkészüléshez ajánlott irodalom
- A 2020-as NAT által meghatározott, 2022 január 1-től alkalmazandó, 2024-től pedig kizárólagosan érvénybe lépő érettségi követelményrendszer szerint összeállított jegyzetek.
- Gyűjtemény a Biológia emelt szintű oktatásához tankönyv, melynek tananyagfejlesztőként társszerzője vagyok (Tankönyvkatalógus).
- Gőz József debreceni biológiatanár által összeállított középszintű és emelt szintű írásbeli feladatok tematikus gyűjteménye BioTéka
Ajánlások, a tanfolyamon résztvevő diákok értékelései
Kedves Tanár úr!
Szeretném megköszönni Önnek a felkészítést és azt a sok energiát, amit adott nekünk az októberi érettségire való felkészítés során. Nagyon boldog vagyok, ma megtudtam a biosz emeltem eredményét, 95% lett. A tanár úr óráin kezdtem el készülni életemben először a biosz emeltre, így összesen 4 hónapot töltöttem készüléssel teljes munka mellett, az eredmények magukért beszélnek. A tanár úr jegyzetei és órái minden kérdésre választ adtak, olyannyira, hogy a szóbelin megkértek próbáljak vázlatosabban fogalmazni mert láthatóan “túl nagy a tudásom”. Azok a kiegészítő információk, amik a jegyzékben találhatók töltötték meg számomra élettel és úgy érzem valódi tudást sajátítottam el.
Nagyon hálás vagyok Önnek! Köszönöm!
Tisztelt Vizkievicz András!
Az előző tanévben csatlakoztam két Bioszférán meghirdetett kurzusához, mely órák segítségével 85%-os emelt biológia érettségit sikerült teljesítenem, illetve megismernem a biológia csodálatos világát, hogy mennyi érdekes dolog környez minket és mennyi minden van, amit még nem tudunk.
Ez úton szeretném megköszönni munkáját, amivel létrehozta a Bioszféra oldalát, amellyel szert tehettem a teljes biológia tudásomra. Ezek mellett tiszteletre méltó, hogy az oldal működtetése mellett csoportos online órákat tart (még betegen is). Hatalmas segítséget nyújt azok számára, akik tanulni szeretnének, de nincs olyan anyagi helyzetük, hogy tudjanak fizetni saját magántanárt vagy esetleg szükségük van a tanuláshoz arra is, hogy szóban hallják többször a tananyagot. Hatalmas segítséget nyújtott az órákon több százszor elhangzott fogalmak említésével, mellyel szinte tudtam mikor mire kérdeznek rá az írásbelin és a szóbelin egyaránt.
A nyár folyamán is meg szerettem volna már írni ezt az e-mailt, de most így egyetem kezdés után döbbentem rá ismét az órákon ülve, hogy milyen szerencsés vagyok, hogy jelentkeztem a kurzusokra, mert jelenetnősebb nagyobb tudásra tehettem szert, mint társaim.
Azóta is minden biológiát tanuló ismerősömnek ajánlom az oldalt és az online kurzusokat is.
Tisztelt Tanár úr!
Online feladatmegoldó kurzusra jártam Önhöz, illetve honlapjának, és jegyzeteinek rendszeres olvasója vagyok. Nagyon hálás vagyok a segítségéért, a türelméért az órákon és végül, de nem utolsó sorban az önzetlenségéért, hogy ingyen elérhetővé tette a jegyzeteit. Az Ön segítségével 94%-os emelt biológia érettségit írtam. Nagyon szépen köszönöm!
Tisztelt Vizkievicz Tanár Úr!
Ismeretlenül is szeretném megköszönni alaposságát, szakértelmét és önzetlenségét, amiért összeállította és elérhetővé tette a jegyzeteit! A kidolgozott témakörök nem csak a felkészüléshez nyújtottak pótolhatatlan segítséget, de az unalmas (és sokszor haszontalan) fakultációs órák után visszaadták a lelkesedésemet. Minden nehézség ellenére öröm volt ezekből a jegyzetekből tanulni! (Az emelt biológia érettségim 88%-os lett, szinte biztosan felvesznek a Semmelweis Egyetem általános orvos szakára.)
Nagyon hálás vagyok Önnek!
Kedves Vizkievicz András!
Szeretném megköszönni a tanfolyamos órákat és a bioszférás jegyzeteket, ezek segítségével sikerült 89%-os emelt szintű biosz érettségit tennem, ami valószínűleg elegendő lesz arra, hogy a kémiával együtt felvegyenek az orvosi egyetemre. Hálás vagyok a precizitásáért és azért, hogy mindig lehetett kérdezni. Jó szívvel fogom a jegyzeteit és a tanfolyamot ajánlani olyan ismerőseimnek és barátaimnak, akik szintén emelt szintű biosz érettségire készülnek.
Kedves Vizkievicz Tanár Úr!
Szeretném megköszönni üzenetben az emelt biológia érettségire felkészítést. A feladatmegoldó csoportba jártam, és a bioszféra jegyzetekből tanultam. Hihetetlenül sok segítséget adtak, igazi kincs az interneten! Összességében 92%-ra sikerült az érettségi, így sikerülhet bekerülni az állatorvosi egyetemre.
Nagyon köszönöm és további sikeres munkát kívánok!
Tisztelt Vizkievicz András!
Azért írom Önnek ezt a rövid levelet, mert köszönettel tartozom. Elhatároztam, hogy egyetem mellett felkészülök emelt biológiára, ráadásul mindezt 4 hónappal érettségi előtt. Ehhez az Ön jegyzeteit használtam, mert azt találtam ezek a legjobbak bármilyen más tankönyvvel szemben. A mai nap jöttem ki az eredményhirdetőről egy 91%-os tanúsítvánnyal a kezemben.
Rendkívül hálás vagyok a munkájáért, és azért is, hogy publikussá tette ezt a fáradtságosan összegyűjtött anyagot.
Köszönöm a segítségét!
Kedves Tanár úr!
Egyik osztálytársammal közösen találtunk rá a Tanár úr órájára, ő orvosi pályára szeretne menni. Emlékszem, amikor az első órájáról jöttünk haza, teljesen le voltunk nyűgözve, hogy 90 percen keresztül, megszakítás nélkül képesek voltunk figyelni a Tanár úrra, hihetetlen, hogy hogy fenn tudta tartani a figyelmünket, az óra izgalmas és pörgős volt. Nyilván nem minden alkalommal tudtam 100%-osan odafigyelni, viszont nagyon szerettem az órákat, nem egyszer kifejezetten vártam, hogy jöhessek és tanulhassak. Utoljára nem is tudom mikor álltam hozzá így az iskolához. Azon is gondolkoztam egy ideig, hogy inkább kihagyok egy évet, mert annyira kiábrándultam az oktatási rendszerből, úgy voltam vele, inkább pihenek egy évet, minthogy teljesen kiégjek egyetemen. Viszont, kifejezetten ebben az utolsó hétben, amikor délelőttönként a Tanár úr órájára jártam be és csak utána mentem az iskolába, rájöttem, hogy szeretek tanulni, csak nem az én iskolában, és most nyilván nem is azt tanulom, ami érdekel, miután nincs is biológia órám, de ha egyetemre bekerülök, nagyon remélem, hogy ott lesz 1-2 olyan jó tanárom, mint amilyen a Tanár úr volt, mert akkor nem lesz panaszra okom. Szeretném megköszönni, hogy felkészített az érettségire, újra megszerettette velem a biológiát és minden órán nagyon kedves is volt, sose féltem kérdezni vagy éreztem magam butának. A Tanár úr biztosított arról is, hogy vannak még jó tanárok, akik szeretik, amit csinálnak és jól is csinálják azt.
Messze a legnagyobb tudással rendelkező tanár, akivel találkoztam az évek során. Hihetetlenül nagy elkötelezettséget érez a munkája iránt. Emellett rendkívül segítőkész és fáradhatatlan. Mindig figyelembe vette a diákok igényeit, így akár többször is átrágtuk magunkat egy-egy nehezebben megérthető részen. Ennek ellenére tartottuk az ütemet, így nem érzem, hogy bárhol elcsúsztunk volna és kevesebbet kaptunk volna. Nagyon sokat segítettek a kisebb számonkérések, mert ezáltal tudtam, melyik témakör az, amivel többet kell foglalkoznom, annak ellenére, hogy azt hittem, elegendő a tudásom. A jegyzeteknél jobbat el sem tudok képzelni:D Minden információt megtaláltam bennük. Logikus a felépítése is, így a hosszas kereséseket is átugorhattam azzal, hogy csak megnéztem a címeket. Én idén megismétlem a biológia érettségit, mivel tavaly sajnos nem volt elegendő a tudásom. Ebben az évben, mikor Vizkievicz Tanár Úrtól tanulhattam, úgy érzem sokkal inkább uralom a biosz, mint valaha.
A Tanár Úr minden órán érthetően, átláthatóan adta át a biológia tananyagot, mindig készségesen válaszolt a kérdéseinkre, az órákon örömmé tette a biológia tanulást. Sokat segített, hogy sosem éreztem azt, hogy szégyen lenne kérdezni és hogy hallottam a Tanár Úr hangján a lelkesedést a tantárgy iránt és a kedvességet felénk
Bár én már kijártam a gimit, ilyen jó tanárt még sehol sem találtam!:) Még a biológia kevésbé szerethető részeit is megkedveltette velem. Szaktudása pedig messze meghaladja egy átlagos középiskolai tanárét, az összes komplikált kérdésünkre tudja a választ:)
Szeretném megköszönni az odaadó és kemény munkáját. Az órák sebessége, üteme megfelelő volt, sohasem éreztem, hogy rohannánk vagy nem lenne idő a felmerülő kérdésekre. Mindig türelmesen elmagyarázta, akár többször is, a nehezebb témarészeket. Szemléletes ábráival és színes rajzaival követhetővé és érthetővé tette a leckéket. A vázlatait nagyon szerettem, mert egy-egy anyagrészek más-más témakörökkel is kapcsoltban voltak és azokat ugyanúgy megemlítette, illetve újból leírta azokat. Sokkal összetettebben tudtam tanulni és még ismételtem is.
A Tanár Úr nagyon lelkes és segítőkész volt végig. Az órákon mindig rávilágított a fontosabb összefüggésekre, érthetően és türelmesen magyarázott. A lényegesebb részek gyakori ismétlésével és kihangsúlyozásával sok információ rögzült bennem az órákon csupán hallás után. Én nagyon hiányos tudással kezdtem a felkészítőt, több olyan témakör is volt, amiről itt hallottam először. Ez az egy év során rengeteget fejlődtem, a tanfolyam végére pedig sikerül egy átfogó és biztos tudást kapnom, amivel magabiztosan és optimistán vághatok neki az érettséginek.
Abszolút meg vagyok elégedve a Tanár Úrral! Derült kedvével mindig meg tudott minket nevettetni, még ha szárazabb is volt az adott téma. Bíztatott minket, hogy bátran tegyük fel a kérdéseinket valamint a nehezebb anyagokat alaposan elmagyarázta. Percre pontosan az ütemtervet követtük így április közepére már végeztünk is a biológiával az által, nekem volt is időm többször is átismételni a már megtanult anyagokat. Az óra eleji dolgozatok segítettek, hogy megerősítést kapjak a már tanult témakörökkel kapcsolatban, így úgy gondolom, hogy elegendő volt a tanultak számonkérése és azok részletes átbeszélése. Valamint, mindig vissza-vissza tértek a már tanult témák az órákon, hiszen a biológia egy olyan tantárgy, ahol minden mindennel összefügg, így futólag még a régebbi anyagokat is át volt időnk átnézni. Minden információ benne volt minden jegyzetben, így sose kellett más oldalakon keresgélnem a témákkal kapcsolatban megválaszolatlan kérdéseimet. Viszont, a jegyzetekben lévő linkek nem érhetőek el így sose tudtam azokba belenézni. Elérhetővé tenném azokat minden bioszférás diáknak már a tanfolyam elején. Persze az én előre haladásom tőlem függ, viszont a Tanár Úr számonkérései és órái mindig sok kedvet és motivációt adtak, hogy leüljek és bioszozzak. Ez a tanfolyam segített megszerettetni velem a tanulást. Túl is szárnyalta az előzetes elképzeléseimet ez a tanfolyam. Tavaly nem vettek fel, így elég csüggedt voltam emiatt, hiszen nem tudtam kellőképp felkészülni az érettségire. Viszont a Tanár Úrral való munka sok motivációt adott az évben, és most érzem igazán, hogy megérett a sok munka gyümölcse. Nagyon hálás vagyok, hogy egy ilyen ember készített fel engem újra a biológia érettségire.
További értékelések olvashatók itt.
Korábbi fakultációs csoportok diákjainak ajánlásai olvashatók itt.
A tanfolyamot vezető tanár Vizkievicz András bemutatkozása olvasható itt.
Elérhetőségek
vizkieviczandras@gmail.com, telefon: 06308151140, Városmajori Gimnázium 1122 Városmajor u. 71.
Tisztelt Látogató!
Ez az oldal azzal a szándékkal jött létre, hogy segítséget nyújtson azoknak a középiskolás diákoknak, akik valamilyen okból az átlagosnál nagyobb érdeklődést mutatnak a biológia tudománya iránt. A fokozott figyelem lehet csak egyszerű érdeklődés, de fakadhat továbbtanulási szándékból is. Számukra készültek azok a jegyzetek, amelyek segítenek felkészülni az emelt szintű biológia érettségi vizsgára.
Kellemes böngészést és sikeres tanulást kívánok!
Vizkievicz András
Bemutatkozás
1991-ben végeztem az ELTE TTK biológia-kémia szakán. Az egyetem negyedik és ötödik évében az egyetem Biokémia Tanszékén géntechnológia témakörében dolgoztam, s szakdolgozatomat is ebből a témából írtam. 1991 óta első munkahelyemen, a Városmajori Gimnáziumban dolgozom.
2011-ben a Budapest XII. kerületi Hegyvidéki Önkormányzat a szakmai tevékenységem elismeréseként a Hegyvidék Ifjúságáért Díjat adományozta.
Az indoklásban ez állt: "Vizkievicz András a Városmajori Gimnázium tanárának ez az első munkahelye, melyhez hűséggel kötődik. Az iskolában működő biológia tagozat elindításában és felfejlesztésében, tantervének kidolgozásában jelentős szerepet vállalt. Éveken keresztül munkaközösség vezetőként segítette az iskolai szakmai munkát. A biológia emelt szintű képzés legsikeresebb tanára, versenyeken kiváló eredménnyel szerepelnek diákjai. A természettudományok megszerettetése, a biológiai kutatások iránti fogékonyság megteremtése a specialitása. Diákok biológia emelt szintű képzésre vagy fakultációra, s utóbbiakban emelt szintű képzésre is szinte az ő neve miatt jelentkeznek, magas színvonalú munkájának a híre az iskola biológia oktatásának védjegyévé vált. Tanár úr órái a legnépszerűbb órák a gimnáziumban, szemléltető módszere, átfogó, érthető, világos előadói stílusa minden elismerést megszerez számára. Az ő tanítványait a Semmelweis Egyetemen kiemelt figyelemmel fogadják."
Az Emberi Erőforrások Minisztériuma a "kiemelkedően eredményes és áldozatos tehetségsegítő munka elismerésképpen" a "Bonis Bona - A Nemzet Tehetségeiért" díjat adományozta.
Az előterjesztésben az alábbiak szerepeltek:
A természettudományok megszerettetése, a biológiai kutatások iránti fogékonyság megteremtése a specialitása. Tanítványainak többsége az orvosi pályán vagy kutatóként dolgozik, valamint a természettudományos szakterületeken is doktori fokozatot szerzők többsége az ő tanítványai. Az iskolát a Semmelweis Egyetem a Tanár úr munkássága alapján kérte fel partnerintézményi együttműködésre. A sikeres magas számú természettudományos pályára való felkészítés mellett, versenyeredményekkel is büszkélkedhetnek diákjai. A természettudományos tantárgyak mindenki által elérhetővé tételére álmodta meg a bioszféra digitális tananyagot, melynek fejlesztésben tehetséges diákjaival közösen alkotnak, s amely az iskola honlapjáról elérhető. A portál folyamatos fejlesztés alatt van, így egyre több a többi tanuló számára is letölthető tanulmányi információk száma. Tanárideálként is kiemelkedő egyéniség, aki hihetetlen népszerűsége, végtelen sok energiával végzett munkája mellett szerény, kedves tanárember tud maradni.
Vizkievicz András 30 éve a Városmajori Gimnáziumban tanít biológiát és kémiát. Az iskola biológia tagozatos tantervi programjának meghatározó szakmai kidolgozója. A biológia tagozatra jelentkező tanulók létszáma az ő munkássága alapján iskolánk kiemelt, 10-szeres túljelentkezéssel működő képzési iránya. Éveken keresztül munkaközösség vezetőként segítette a képzés hatékonyságát, de a SE felvételi, közép és emelt szintű érettségi felkészítő tanáraként is kiemelkedő eredményeket mondhat magáénak. Az iskolában ő alakította ki a biológia emelt szintre felkészítő fakultáció rendszerét, szakterületén tapasztalható végtelen precizitása, a tantárgy iránti elkötelezettsége, a megbízhatósága, a szakmai feladatteljesítési szerénysége és tisztessége pótolhatatlanná teszi őt. Varázsos egyénisége miatt a diákok a biológia emelt szintű képzésre vagy fakultációs képzésre is szinte csak az ő neve miatt jelentkeznek. Óráinak összeállítása, módszertana, valamint a szakma iránt érzett elkötelezettsége példaértékű.
- Szakértői feladatok ellátása a közép- és az emelt szintű írásbeli feladatokkal kapcsolatosan
- Az Oktatási Hivatal megbízásából a Gyűjtemény a Biológia emelt szintű oktatásához tankönyv tananyagfejlesztője
- Biológia tantárgyból a közös írásbeli érettségi/egységes írásbeli felvételi tétel-összeállító bizottsági tagság
- Biológia tantárgyból a közös írásbeli érettségi/egységes írásbeli felvételi eljárásban feladatkészítő tanár
- Emelt szintű érettségi vizsga biológia tantárgyi bizottságának elnöke
- Általános érettségi vizsgaelnök
- A Semmelweis Egyetem megbízásából emelt szintű érettségi vizsgára felkészítő tanár
- Szóbeli felvételi eljárás során szakmai kérdezői feladatok ellátása a Semmelweis Egyetemen
- A Városmajori Gimnázium biológia-kémia munkaközösségének vezetője
- Tehetségpártolók Baráti Körének alapító tagja
- A Semmelweis Egyetem partneriskolai programjának összekötője
- Szent István Egyetem: Tudományos Diákköri Konferencia zsűritagság
- Kis Bolyai-díj kuratóriumi tagság






Érettségi 2026
Ezen az oldalon a 2020-as NAT által meghatározott, 2022 január 1-től alkalmazandó, 2024-től pedig kizárólagosan érvénybe lépő kétszintű biológia érettségivel kapcsolatos hasznos linkek találhatók.
- A biológia vizsgatárgy 2024. május-júniusi vizsgaidőszaktól érvényes vizsgakövetelményei és a vizsga leírása
- Érettségi pontszámító kalkulátor
- Érettségi pontszámítás 2024
- Mintafeladatsor 2024
- Mintafeladatsorok 2017
- Emelt szóbeli mintatételek 2017
- Emelt szóbeli mintatétel 2024
- Tájékoztató az emelt szintű szóbeli vizsgához, az A) feladatok címe
- Korábbi érettségi időszakok feladatsorai biológiából
- 100/1997. (VI. 13.) Korm. rendelet az érettségi vizsga vizsgaszabályzatának kiadásáról
Az egyetemek által adható intézményi pontok.
- Semmelweis Egyetem
- Szegedi Tudományegyetem
- Pécsi Tudományegyetem
- Debreceni Tudományegyetem
- Állatorvostudományi Egyetem
- ELTE TTK
Biológia szóbeli érettségi tételek kidolgozott A) feladatai
A szóbeli tételeket Vizkievicz András dolgozta ki.
A kidolgozott biológia szóbeli tételek teljes mértékben magukba foglalják mindazokat az aktuális követelményrendszer által elvárt vizsgakövetelményeket - kulcsfogalmakat, gondolkodási műveleteket -, melyek az adott témakörhöz tartoznak. Az oldalon a böngészést nagyban segíti a kulcsszavakra történő keresés ctrl+f használatával.
Tekintve, hogy az oldal minden tanulni vágyó diák számára költségmentesen elérhető, a tételek anyagának bármilyen kereskedelmi célű felhasználásához semmilyen módon nem járulok hozzá. Sikeres felkészülést kívánok!
Tájékoztató az emelt szintű szóbeli vizsgához, az A) feladatok címei (pdf)
Szóbeli vizsga általános szabályai
Az emelt szintű szóbeli vizsga központi tételsor alapján zajlik. Feleléskor a kifejtés sorrendjét a vizsgázó választja meg. A vizsgáztatónak lehetővé kell tennie, hogy a vizsgázó gondolatmenetét önállóan fejtse ki, majd – amennyiben a feladat ez – álláspontját is megfogalmazza és megvédje.
A szóbeli tételsor tartalmi jellemzői
A szóbeli vizsgán a vizsgázó tárgyi tudásáról, kifejezőkészségéről, problémaérzékenységéről tesz bizonyságot.
A tételsor jellemzői
A tételsornak legalább 20 tételt kell tartalmaznia. A tételsornak a követelményrendszer minden fő témakörét érintenie kell. Valamennyi tételhez két feladat – A) és B) – tartozik. Egy tétel két feladata nem vonatkozhat azonos témára.
A tételek jellemzői
A) feladat: A közzétett címeknek megfelelő feladat kifejtése megadott szempontok alapján.
B) feladat: Az érettségi követelményekben dőlt betűvel jelzett követelmények: biológiai problémát tartalmazó feladat megoldása, illetve értelmezése, elemzése, értékelése, tervezése megadott szempontok alapján.
A szóbeli vizsgarész értékelése
Az A) feladat értékelésénél 20 pont, a B) feladatnál 25 pont adható a tartalomra, és összesen 5 pont az A) és B) feladatok kifejtésének módjára. A központi értékelési útmutató rögzíti az egyes tételek kifejtésének elvárt összetevőit és az ezekre adható, a 20, illetve 25 pont felosztásával kialakított maximális részpontszámokat.
tetelek.png) |
1. tétel: Az élet általános jellemzői |
Az élőlények életjelenségeket mutatnak. Életjelenségeken olyan folyamatokat értünk, amelyek csak az élő szervezetekre jellemzőek. Ezek teszik lehetővé a változó környezetben az élőlények életben maradását és a környezethez való alkalmazkodását.
A legfontosabb életjelenségek
- Az anyagcsere, melynek során az élőlények a környezetből anyagokat és energiát vesznek fel, amiket beépítenek, ill. átalakítanak. A felvett anyagok, ill. az egyes saját vegyületeik lebontásakor felszabaduló energia felhasználódik az életműködések fenntartására. Ezzel párhuzamosan történik a felesleges bomlástermékek leadása.
- Egyediség, viszonylagos elhatárolódás, ami nem elszigetelődést jelent, hiszen az élő rendszerek a környezetükkel állandó anyag- és energiaforgalmat bonyolítanak le, azaz nyílt anyagi rendszerek.
- Homeosztázis, amely a belső környezet szabályozott állandósága. A homeosztázis a szervezet egységes működésének – belső egység - az eredménye, melyet a szabályozás tesz lehetővé.
- Szabályozottság, vagyis a változó külső és belső körülményekhez az élő szervezetek működésük rugalmas megváltoztatásával képesek alkalmazkodni. A szabályozás alapja az ingerlékenység.
- Az ingerlékenység, melynek során az élőlények a környezetükből különféle ingereket fognak fel, melyekre működésük megváltoztatásával reagálnak.
- A mozgás.
- A szaporodás.
- A növekedés.
- Az öröklődés – kódolt információhordozás és -átadás - és az öröklődő változékonyság.
- Az öröklődés során a szülői tulajdonságok továbbadódnak az utódokba.
- Az öröklődő változékonyság azt jelenti, hogy az öröklött tulajdonságok nem állandók, nemzedékről-nemzedékre változhatnak, ez lehetővé teszi, hogy az utódok tulajdonságai némileg eltérjenek a szülőkétől. Az élőlények változatossága azt eredményezi, hogy az egyes egyedeknek különbözők az esélyei az életben maradásra és a szaporodásra. Az élőlények változatossága teremti meg az alapját az élővilág törzsfejlődésének, az evolúciónak.
- Halandóság, mivel csak az élő képes meghalni.
- Evolúció, melynek során az élővilág többnyire fokozatosan és szüntelen fejlődik. A törzsfejlődés hátterében olyan folyamatos változások sorozata áll, melynek során a fajok öröklődő jellegei nemzedékről nemzedékre változnak.
Az élet szénalapúsága
Az élőlények elkülönülése a környezetüktől több szinten is megnyilvánul. Egyik legszembeötlőbb eltérés az egyes kémiai elemek előfordulási gyakoriságában megmutatkozó különbség.
Az elemek előfordulási gyakoriságában az egyik legnyilvánvalóbb különbség a szén esetében mutatkozik meg. Ennek az a magyarázata, hogy az élőlények testének felépítésében nagyrészt szerves vegyületek vesznek részt, mint pl. nagyobb csoportjaik a zsírok, a szénhidrátok, a fehérjék, a nukleinsavak. A szerves vegyületeknek pedig definíciószerűen alapvetően a szén- és hidrogéntartalmú vegyületeket tekintjük, mivel minden szerves molekula alapvázát szénatomok láncolata alkotja.
A szén az egyetlen elem, melynek atomjai szinte korlátlan számban képesek egymással összekapcsolódva különféle stabil óriásmolekulákat létrehozni, mivel a kisméretű szénatomtörzs körül az erős kötőelektronpárok szimmetrikusan, többnyire tetraéderesen helyezkednek el.
Az élővilág egységességét jól tükrözi az is, hogy a legkülönbözőbb élőlények kémiai összetétele (az elemek és vegyületek szintjén is) rendkívüli hasonlóságot mutat.
A biológiai rendszerek anyagi rendszerek. Az anyagi rendszereket általában anyag- és energiaáramlás jellemzi és működésük fenntartásához sokszor energiára van szükség. A Föld globális rendszerének működéséhez a meghatározó külső energiaforrás a Nap sugárzó energiája.
Az élő rendszerek, mint nyílt anyagi rendszerek a környezetükből anyagot és energiát vesznek fel, ill. adnak le.
Az élő rendszerek energiaforgalma
A sejtek – és általában az élő rendszerek – működésükhöz, életfolyamataik fenntartásához folyamatosan energiát igényelnek.
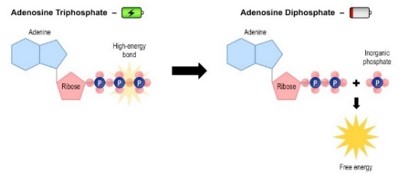
Az élő szervezetekben a legjelentősebb a kémiai energia. A kémiai energia kémiai kötések hasadásához szükséges, illetve kialakulásukkor felszabaduló energiamennyiség.
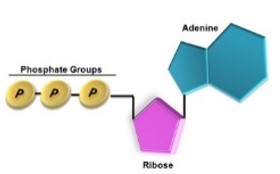
A biológiai energia a kémiai energia egy speciális fajtája, amely a sejtekben nagy energiájú kémiai kötésekkel kapcsolatos. A nagy energiájú kötések energiát tároló vegyületekben találhatók meg. Ilyen minden élő szervezetben megtalálható energiatároló és -szállító vegyület az ATP.
 |
 |
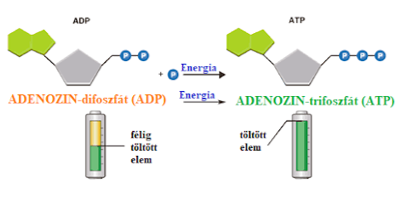 |
A sejtek ATP-készlete állandó, energiaigényes folyamatokban energiát szolgáltatva bomlik, energiatermelő folyamatokban pedig a felszabaduló energiát megkötve regenerálódik.
Az élőlények a környezetüktől elkülönülő, de azzal kölcsönhatást folytató rendszerek.
Az élő rendszerek az elkülönülés okán környezetükkel állandó egyensúlytalanságot tartanak fenn, hiszen
- egyrészt belső környezetük tényezőinek értékei – kémhatás, ionkoncentrációk, hőmérséklet stb. – jelentősen eltérnek a környezetben tapasztalható értékektől,
- másrészt, mint már korábban volt róla szó anyagi összetételben – elemek előfordulási gyakorisága, szerves vegyületek – is jelentős különbség figyelhető meg.
- Továbbá egy érdekes különbség a rendezetlenségben nyilvánul meg, melynek mértékét az entrópiával fejezzük ki. Általában elmondható, hogy az élettelen természetben minden folyamat a rendezetlenség mértékének növekedése irányába hat, aminek oka az energiaminimumra való törekvés, ami egy általános elv az univerzumban. Az élő rendszerek rendezettsége és így energiatartalma jóval nagyobb, mint az élettelen környezeté.
Ez az egyensúlytalanság az élő állapot egyik legalapvetőbb jellemvonása. Ahhoz, hogy az élő rendszer folyamatosan fenn tudja tartani az egyensúlytalanságot, energiára van szükség, hiszen minden életjelenség végső soron munkavégzés. Az élet megszűnésével ez az egyensúlytalanság, a rendezettség is megszűnik, átadja helyét az élettelen világban fennálló egyensúlynak, rendezetlenségnek.
Sokszor olvashatjuk, hogy az élőlények egyensúlyban vannak a környezetükkel, ami ebben az esetben azt jelenti, hogy a természetben az élőlények hosszútávon stabilan együtt tudnak létezni környezetükkel.
Az intermedier anyagcsere
Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az anyagcsere során az élőlények a környezetből anyagokat és energiát vesznek fel, ill. adnak le.
Az élőlények a felvett anyagokat átalakítják:
- beépítik, ill. felépítő folyamataikban használják fel, vagy
- energianyerés céljából lebontják, majd
- a felesleges, ill. a fel nem használható anyagokat eltávolítják.
A felvétel és a leadás közötti átalakító folyamatok összességét intermedier - köztes - anyagcserének nevezzük, melynek köszönhetően az élőlények a szüntelen változó környezetben fenntartják egyediségüket, viszonylagos állandóságukat, rendezettségüket, működésük rugalmas változásával.
A sejt, mint működési alapegység
A sejt az élővilág legkisebb, önálló életre képes, életjelenségeket mutató szerveződési egysége. Az élőlények, mint biológiai rendszerek egyik meghatározó alaptulajdonsága a sejtes szerveződés.
Legelsőként Robert Hook (1635– 1703) vizsgált sejteket saját készítésű mikroszkópjának segítségével. Majd a XIX. században fogalmazódott meg a sejtelmélet, miszerint minden élőlény egy vagy több sejtből áll, s minden sejt már létező korábbi sejt kettéosztódásából keletkezik.
Az élővilág rendkívüli sokszínűségének, az élőlények szinte végtelen változatosságának ellenére, a különféle élő rendszerek felépítése és működése alapjaiban számos közös sajátossággal bír. Ez nem meglepő, hiszen a mai élőlények egy hosszú törzsfejlődési folyamat eredményeként jöttek létre, aminek kiindulópontja egyetlen közös ős volt. Tehát a ma élő összes élőlény erre a kb. 3,8 milliárd éve létezett ún. egyetemes közös ősre (Last Universal Common Ancestor, LUCA) vezethető vissza.
Az eltérő életformák - mint pl. növényi, állati, gomba, baktérium - egyedeit felépítő sejtek különbségeik mellett számos alapvető jellegeikben lényegi hasonlóságot mutatnak, mind felépítésükben, mind működésükben.
- Minden sejt sejthártyával határolódik el a környezetétől, ill. a sejthártya segítségével bonyolít le anyagforgalmat a környezetével.
- Közös sajátság a sejtplazma, az anyagcsere-folyamatok színtere.
- A molekuláris biológia fejlődése nyomán az is világossá vált, hogy az élőlények kémiai összetétele, alapvető anyagcsere folyamatai, a kémiai energia tárolásának módja is azonos.
- Minden élőlény genetikai információt tároló anyaga DNS, ill. az információt kódoló, továbbító és kifejező rendszerek is alapjaiban hasonlóan szerveződnek.
2. tétel: Az élőlények felépítésének fizikai, kémiai vizsgáló módszerei |
A biológiai kutatások módszerei
A biológiai kutatásokban alkalmazott módszereket a kutatás tárgya alapján választjuk ki. Ilyenek pl.
Megfigyelés, melynek során a biológiai jelenséget anélkül vizsgáljuk, hogy bármilyen módon beavatkoznánk, megváltoztatnánk a körülményeket.
Leírás, mikor megfigyeléseink tapasztalatait rögzítjük jegyzőkönyv formájában.
Összehasonlítás hasonló vagy éppen eltérő jelenségekkel, melynek során megállapítjuk és következtetéseket vonunk le a hasonlóságokból és a különbségekből.
A kísérlet olyan kutatási eljárás, melynek során a kísérletező végez valamilyen tevékenységet, majd megfigyeli annak következményeit. A kísérletnek megismételhetőnek kell lennie és azonos eredményre kell vezetnie ahhoz, hogy az eredményt tudományos eredménynek nevezhessük.
A kísérletek kapcsán meg kell határoznunk a kísérleti változók fogalmát:
- Független változó: az a tényező, melynek hatását vizsgáljuk (az eredményt grafikonon ábrázolva értékeit az x tengelyen tüntetjük fel).
- Függő változó: azon tényező, melyet a független változó módosít, melynek a változását mérjük (értékeit az y tengelyről olvashatjuk le).
- A rögzített változókat (egyéb körülményeket, melyek befolyásolhatják a kísérlet eredményeit) állandó értéken tartjuk, hogy egyértelmű legyen az összefüggés a független, ill. a függő változó között.
Kísérletezéskor a kutató szabályozza a feltételeket, tervszerűen avatkozik be a jelenségbe. A megfigyeléstől a kísérlet abban különbözik, hogy a vizsgálat során a körülményeket, változókat mi határozzuk meg. A kísérletek megtervezésének, végrehajtásának fontos feltételei;
- a változók pontos mérhetősége,
- a megismételhetőség,
- kontrollvizsgálat, ellenőrzés, melynek során a jelenséget beavatkozás nélkül figyeljük meg, majd vizsgálati eredményeinket ezzel a tapasztalattal vetjük össze.
Modellalkotás során a modell segítségével a vizsgált valóságos rendszer lényegi tulajdonságait ismerjük meg. A modell a valóság egyszerűsített leképezése. A modellt összevetjük a forrásául szolgáló valóságos jelenséggel, majd következtetéseket fogalmazunk meg. A modellalkotásnak rendkívül sokféle formája létezik, az egyszerűbb anatómiai modellektől, mint pl. az emlősök légzőműködéseit bemutató Donders-féle modelltől, a modern számítógépes rendszerekig.
Mérés
A mérés során egy adott mennyiség értékét határozzuk meg úgy, hogy a mért értéket a mértékegységhez viszonyítjuk. A mérés során azt állapítjuk meg, hogy a mérendő mennyiség hányszorosa a választott mértékegységnek. Tervszerűen végrehajtott művelet, kivitelezéséhez mérőeszközt használunk.
Fénymikroszkóp
Összetett optikai rendszer. Főbb részei:
- talpazat,
- a tubus,
- a revolverfoglalat,
- mozgatható tárgyasztal,
- kezelőgombok,
- megvilágító rendszer.
A tubus a szemlencse (okulár) és a tárgylencse (objektív) befoglalására szolgál. A tárgyasztal az objektív alatt található, amely a preparátum elhelyezését teszi lehetővé.
Az optikai rendszer nagyítása az objektív és az okulár nagyításának szorzatával egyenlő. A fénymikroszkóp nagyítása maximum 1500-szoros, elméleti felbontóképességük 0,2 mikrométer.
Nagyítás alatt egy kép méretének megnövelését értjük, jele N. A nagyítás a képméret (K) és a tárgyméret (T) hányadosa. N=K/T
A felbontóképesség az a legkisebb távolság, amelynél az elhelyezkedő tárgypontok még különálló képpontokként képződnek le. Az emberi szem felbontóképessége 0,2 mm.
Elektronmikroszkóp
A fénymikroszkópok látható fényt (400-800 nm), az elektronmikroszkópok ugyanakkor jóval kisebb hullámhosszú, mágnesekkel fókuszált elektronsugarat használnak képalkotásra.
Az elektronmikroszkóppal 100 000-szeres nagyságrendű nagyítás és 0,5 nm-es felbontóképesség érhető el, amivel már nem csak a sejtek, hanem a sejtalkotók finomszerkezete is tanulmányozható.
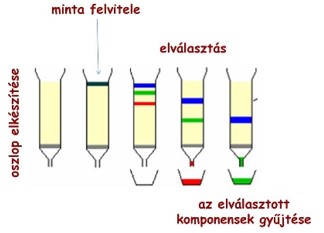
Kromatográfia
Gáz- vagy folyadékelegyek szétválasztására szolgáló vegyi eljárás. Az eljárás során az elválasztandó alkotórészek két fázis között oszlanak meg;
- az egyik fázis álló,
- a másik mozgó fázis (minta), tulajdonképpen a szétválasztandó keverék, amely átáramlik az álló fázison.
Az álló fázis többnyire szilárd anyag.
A mozgó fázis lehet
- folyadék vagy
- gáz, ami egyben folyadék- vagy gázkromatográfiát jelöl.
Az álló fázisként szolgáló finomszemcsés adszorbens - mely fontos sajátsága a nagy fajlagos felület -, lehet például Al2O3, aktív szén, szűrőpapír (cellulóz). Ez alkotja a kromatográfiás oszlopot. Erre viszik fel a szétválasztandó elegyet, ami mint mozgó fázis, mozog az álló fázison keresztül.
Az álló fázis felületén a mozgó fázis komponensei különböző erősséggel megkötődnek, adszorbeálódnak. Az egyes alkatrészek aszerint választódnak el, hogy mennyire erősen kötődnek az álló fázisra. Azok a komponensek, amelyek erősebben adszorbeálódnak, kisebb sebességgel vándorolnak, mint azok, amelyek kevésbé adszorbeálódnak, ennek megfelelően a folyadékmozgás irányában az alkotórészek elválasztódnak egymástól és gyűrű formájú mozgó zónákba összpontosulnak.
 |
 |
 |
 |
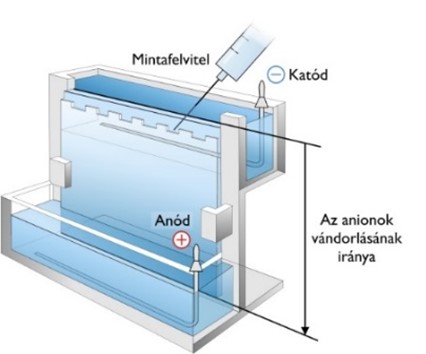
Gélelektroforézis
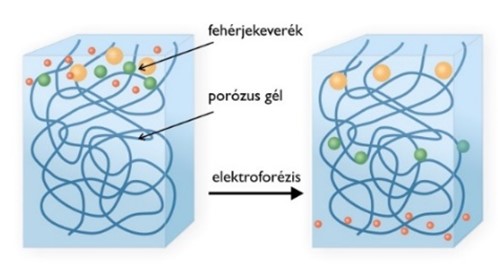
A gélelektroforézist leggyakrabban nukleinsavak és fehérjék méret szerinti elválasztására használják. Az eljárás segítségével a makromolekulákat három fizikai tulajdonságuk, a méretük, alakjuk és a töltésük alapján tudjuk elválasztani.
A gélelektroforézis alapelve, hogy a töltéssel rendelkező molekulák elektromos térben, össztöltésüknek megfelelően, az ellentétes töltésű elektróda felé vándorolnak. A vándorlás sebessége többek között függ a molekula töltésétől, tömegétől és alakjától.
A gél egy térhálós szerkezetű anyag, mely a molekulák méretétől, alakjától függően lassítja azok mozgását. Ha a molekula mérete kicsi a pórusokhoz képest, gyorsan mozog, ha nagy, a molekula szinte mozdulatlan marad a gélben.
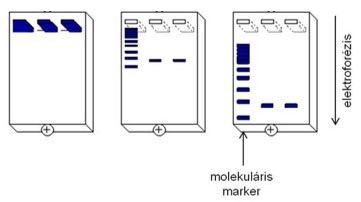
Méret szerinti elválasztás esetén az ismeretlen mintáink mellett mindig kell futtatnunk egy ismert tömegű/hosszúságú fehérjéket/nukleinsavakat tartalmazó ún. molekulamarkert.
A megfelelő ideig történt futtatás után a feszültséget kikapcsoljuk és a gélt megfestjük, pl. a nukleinsavak esetében valamilyen, a nukleinsavakhoz kötődni képes fluoreszcens festék oldatával, majd az elválasztott nukleinsavakat a fluoreszcens festéket gerjeszteni képes fénnyel (pl. UV-fénnyel) láthatóvá tehetjük.
Centrifugálás
Elegyek komponenseit szétválaszthatjuk centrifugálással is, melynek során a keverék alkotórészei a centrifugális erő hatására tömegük, ill. sűrűségük szerint elkülönülnek.
A centrifuga egy álló és egy forgó rotorból áll. A rotor fogadja be a mintákat tartalmazó centrifugacsöveket. A centrifugális erő - melyet a tengely körül forgó rendszerrel lehet előállítani – hatására a nagyobb tömegű, sűrűségű anyagok a centrifugacsövek alján, míg a kisebb tömegű részecskék a csövek felső részén rétegződnek.
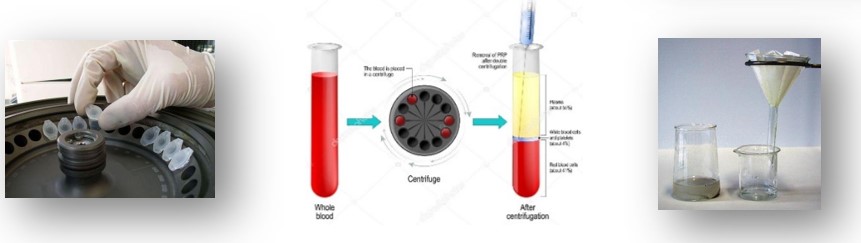 |
Szűrés
A szűrés egy fizikai elválasztási eljárás, melynek során
- szilárd anyagokat különítünk el folyadéktól, ill.
- különböző szemcseméretű részecskéket választunk szét egymástól.
A szűrő egy olyan porózus réteg – pl. szűrőpapír -, ami bizonyos szemcseméretű anyagokat átenged, másokat visszatart.
Diagnosztikai vizsgálatok
Az orvosi eljárás első szakasza, amely
- a beteg panasza okának feltárására, állapotának tisztázására,
- valamely betegség fennállásának igazolására vagy kizárására irányul.
A diagnosztikai vizsgálatok a diagnózis meghatározását teszik lehetővé, mely megállapítás az adott betegség fennállását valószínűsíti.
A diagnosztikai vizsgálatok közé tartoznak
- fizikális vizsgálat,
- képalkotó diagnosztika (pl. röntgen, CT, ultrahang, MR),
- labordiagnosztika,
- patológia (szövettani vizsgálat),
- egyéb eszközös vizsgálat, mint pl. EKG, EEG, endoszkóp.
EKG, EEG
Az emberi test folyadékterei, mivel ionokat tartalmaznak, jól vezetik az elektromosságot, ezért a szív működésekor (EKG), ill. az agyi működés (EEG) során keletkezett elektromos jeleket mérni lehet a test felszínén is. Amennyiben a feszültségváltozásokat az idő függvényében ábrázoljuk, jellegzetes görbéket kapunk.
A különféle képalkotó diagnosztikai eszközökkel feltárás nélkül vizsgálható az emberi test.
Endoszkópos vizsgálat
Segítségével a páciens belső szerveiről, testüregeiről kapható betekintés a testnyílásokon keresztül bevezetett kamera segítségével.
Ultrahangvizsgálat
Elsősorban lágyrészek vizsgálatára használatos. Fájdalom, bódítás nélkül lehetséges a belső szerveket működés közben megfigyelni. A vizsgálófej ultrahangot bocsájt ki, mely a belső szervekben részben elnyelődik, részben visszaverődik. Az egyes szövetek, ill. a különféle szervek eltérő felületei más-más módon verik vissza a kibocsájtott ultrahangot, melyet a műszer érzékelve képpé alakít.
 |
 |
 |
 |
Röntgenvizsgálat
Alapvető vizsgálati módszer a mellkasi – tüdő - betegségek, csonttörések, ortopédiai, fogászati problémák, egyes hasi panaszok diagnosztikájában. A röntgenkészülék nagy energiájú elektromágneses sugarakat – ún. röntgensugarakat - bocsájt ki. A sugárzás bejutva a testbe, a különféle szövetekben eltérő mértékben nyelődik el, vagy másképpen az egyes szövetek különböző mértékben engedik át a sugarakat. Ezért a test mögött elhelyezett röntgenfilmen sötétebb és világosabb területek láthatók. A nagyobb sűrűségű részek, pl. csontok, jobban elnyelik a sugarakat, ezért a fotólemezen világos sávként tűnnek fel, míg a lágyrészek, melyek inkább átengedik a sugárzást, sötét foltokként jelennek meg.
CT
A computer tomográfia több síkban röntgen-rétegfelvételeket készít a vizsgált területről, amiből számítógépes eljárással akár háromdimenziós képek is létrehozhatók.
MR (mágneses rezonancia)
Az MR-készülék által generált rendkívül erős mágneses térben egyes atomok, pl. a hidrogénatomok, elektromágneses rádióhullámokkal történő gerjesztés után alapállapotba visszakerülve szintén elektromágneses hullámokat bocsátanak ki, képpontokat képezve, melyeket egy nagy teljesítményű számítógép részletes felbontású, háromdimenziós képpé alakít.
Az eszköz alkalmas rándulás, ficam, törés, gyulladások, agyvérzés, sclerosis multiplex, epilepszia, gerincferdülés, gerincsérv, csontritkulás, érbetegségek és fejlődési rendellenességek diagnosztikájára.
  |
 |
 |
 |
3. tétel: Az ozmózis |
Diffúzió
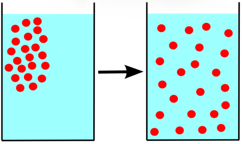
Koncentrációkülönbség hatására a nagyobb koncentrációjú hely felől a kisebb felé irányuló, spontán végbemenő anyagtranszportot diffúziónak nevezzük. A diffúzió eredményeként a rendszerben a diffundáló anyag koncentrációja kiegyenlítődik, a részecskék a rendelkezésre álló teret egyenletesen kitöltik. Elsődlegesen gázokban és oldatokban van jelentősége.
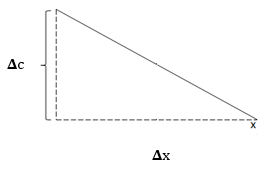
A diffúzió irányát a koncentrációgradiens határozza meg, azaz a koncentráció egységnyi távolságon belüli változása (𝚫𝒄/𝚫𝒙). A grafikon a koncentráció változását jeleníti meg a távolság függvényében.
A diffúziót a részecskék rendezetlen hőmozgása – ún. Brown-féle mozgás – teszi lehetővé. A diffúzió sebessége a diffundáló molekula méretével és a közeg sűrűségével fordítottan, a hőmérséklettel pedig egyenesen arányos.
 |
 |
 |
A diffúziós idő a távolság négyzetével arányos. A diffúzió mikrométeres távolságon gyors, míg centiméterek esetén lassú folyamat.
A diffúziónak óriási a jelentősége az élővilágban, az anyagáramlás alapvető módja, a passzív transzportfolyamatok alapja. Diffúzióval
- lép be pl. az oxigén a tüdő légteréből a hajszálerekbe, a hajszálerekből a szövetekbe, a diffúz légzésű állatoknál a kültakarón át a testbe,
- az akciós potenciálok lezajlásának hátterében szintén a különféle ionok – pl. Na+, K+ – passzív transzportfolyamatai állnak.
- Továbbá diffúzióval – pontosabban ozmózissal, lásd alább – mozog a víz a különböző határfelületeken keresztül, pl. ilyen a növények vízfelvétele a talajból, a vesében a víz visszaszívódása, a bélcsőben a víz felszívódása stb.
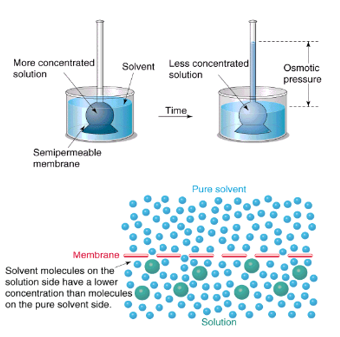
Ozmózis
Az ozmózis folyamatában az oldószer – a víz – féligáteresztő hártyán – ún. szemipermeábilis hártyán – keresztül diffundál a hígabb oldatrész felől a töményebb oldatrész felé. A féligáteresztő hártyák olyan pórusos membránok, amelyek az oldott anyag nagyobb molekuláit nem, csak a kisebb méretű oldószer molekuláit engedik át. Amennyiben az ilyen hártyák két oldalán különböző koncentrációjú oldatok találhatók, akkor az oldószer részecskéi a hígabb oldat felől – ahol a víz koncentrációja nagy – a töményebb oldat felé – ahol a víz koncentrációja kisebb, hiszen az oldatban oldott anyag is van – diffundálnak.
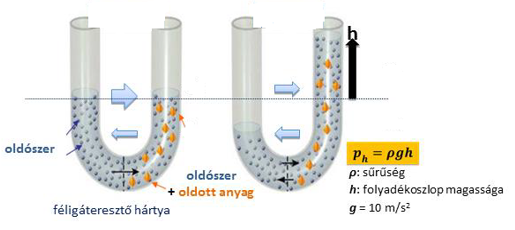
Kísérlet
Egy üvegedénybe desztillált vizet öntünk, majd ebbe belehelyezünk egy üvegcső végére kötözött, cukoroldatot – szacharóz – tartalmazó féligáteresztő tulajdonságú celofánzsákot.
A kísérlet kezdetén az üvegkádban levő víz szintje és az üvegcsőben található cukoroldat vízszintje megegyezik. Néhány óra múlva azt tapasztaljuk, hogy az üvegcsőben levő oldat szintje megemelkedik.
 |
 |
A jelenség hátterében az ozmózis áll.
- A rendszerben koncentrációkülönbség van mind a cukorra, mind a vízre nézve.
- A cukor diffúzióját, nagy molekula mérete miatt, a féligáteresztő hártya megakadályozza.
- A pohárban tiszta víz van, ezért annak térfogategységeiben több vízmolekula van, mint a zsákban lévő cukoroldat térfogategységeiben. Emiatt a diffúzió a vízmolekulákat a zsák belseje felé hajtja.
- Kifelé is lépnek vízmolekulák, de kisebb sebességgel, mint befelé.
- Ha a vízbelépés sebessége v1, a kilépésé pedig v2, akkor a folyamat elején v1 > v2, ezért a zsák folyadékszintje emelkedni fog, de csak addig, ameddig a folyadékoszlop hidrosztatikai nyomása egyenlővé nem válik az ún. ozmózisnyomással.
- Ennek az az oka, hogy az emelkedő folyadékoszlop nyomása fokozza a víz kilépési sebességét, s így v2 végül addig nő, amíg v1-gyel egyenlő lesz.
- Amikor v1=v2, dinamikus egyensúlyi állapot áll be.
Ozmózisnyomásnak nevezzük azt a nyomást, amelyet a tiszta oldószerrel féligáteresztő hártyán át kapcsolatban lévő oldatra kell kifejteni ahhoz, hogy a dinamikus egyensúly beálljon.
Az ozmózisnyomás az oldat koncentrációjával egyenesen arányos. Az oldatban lévő ozmotikusan aktív - amelyek nem jutnak szabadon át a féligáteresztő hártyán - oldott részecskék számát az ozmolalitással fejezzük ki. Tehát minél nagyobb az ozmolalitás, annál nagyobb az ozmotikus nyomás. Az ozmózisnyomás kialakításában az összes ozmotikusan aktív anyag részt vesz.
Az onkotikus (kolloidozmotikus) nyomás az ozmotikus nyomás azon része, amelyet a nem diffundáló kolloidok (főleg fehérjék, a vérben az albuminok) hoznak létre.Fordított – reverz – ozmózisról akkor beszélünk, ha nagyobb külső nyomást alkalmazunk, mint az ozmózisnyomás. Ilyenkor ugyanis oldószer fog kipréselődni a hártyán a hígabb oldat felé. Ez a jelenség az oka annak, hogy a hajszálerek artériás szakaszán (fehérjementes) vérplazma préselődik ki, itt ugyanis a vérnyomás meghaladja a vérplazma vérfehérjék által kialakított ozmózisnyomást (lásd később).
Az élő szervezetben különféle határfelületek rendelkeznek féligáteresztő sajátságokkal, mint pl. a sejtmembránok vagy a kapillárisok fala. E felületek két oldalán a különféle méretű molekulák koncentrációkülönbségét az okozza, hogy az ozmotikusan aktív szerves vegyületek a kolloid mérettartományba sorolhatók, s emiatt ezekre nézve e határfelületek féligáteresztő tulajdonságúak. A kolloid részecskék mérete 1-1000 nm-ig (IUPAC) terjed.
Folyékony közegben, amennyiben a szétoszlatott – diszpergált - részecskék mérete az 1-1000 nm mérettartományba esik, akkor kolloid rendszerről van szó. Tehát a kolloid rendszer nem anyagi minőséget jelent, hanem egy adott mérettartomány határozza meg. A diszpergáló közeg egyébként nemcsak folyadék, hanem gáz (köd, füst), ill. szilárd halmazállapotú is lehet, azonban ennek biológiai jelentősége nincs.
A kolloidok biológiai jelentősége abban áll, hogy a kolloidrészecskék fajlagos felülete igen nagy, ezért jó adszorbensek. Pl. a talajokban található talajkolloidok felületükön ásványi anyagokat képesek megkötni, ami a növények táplálkozása szempontjából fontos.
A kolloid rendszereket többféle szempontból osztályozhatjuk.
- A szétoszlatott anyag halmazállapota szerint.
- Emulzióról beszélünk akkor, ha folyadékban folyadékot oszlatunk szét, vagy
- szuszpenziónak nevezzük a rendszert, amennyiben a szétoszlatott részecskék eredetileg szilárd halmazállapotúak.
- A kolloid rendszereket csoportosíthatjuk a szétoszlatott részecskék minősége alapján.
- Az ún. asszociációs kolloidokban amfipatikus molekulák hozzák létre a kolloid részecskéket, az ún. micellákat,
- a makromolekuláris kolloidokban pedig nagy molekulájú szerves vegyületek (pl. fehérjék, nukleinsavak, poliszacharidok) képeznek kolloid rendszert.
- A kolloidok osztályozása történhet az eloszlatott részecskék közt ható erő szerint.
- A szolokban a kolloid részecskék között ható vonzóerők kisebbek a hőmozgás szétszóró erejénél, ezért a részecskék hidrátburkukkal szabadon elmozdulhatnak. Az ilyen szol állapotú rendszerek folyékony halmazállapotúak, mint pl. tojásfehérje, vér, tej stb.
- A gél állapotú kolloid rendszerekben a részecskék egymáshoz kapcsolódnak, haladó mozgást nem végeznek, így halmazállapotuk szilárdnak mondható, többé-kevésbé alakállandóak, ilyen pl. a főtt tojás, alvadt vér, tejföl.
- A kolloid oldatokat osztályozhatjuk az ozmózisnyomásuk szerint.
- Az izotóniás oldatoknak az ozmózisnyomása megegyezik a vérplazma ozmózisnyomásával, azaz izoozmotikusak. Ilyennek tekintjük a mindennapi orvosi gyakorlatban használatos fiziológiás sóoldatot, mely 0,9%-os NaCl-oldat koncentrációjának felel meg.
- A hipertóniás oldatok ozmózisnyomása nagyobb – hiperozmotikusak –,
- a hipotóniás oldatoknak az ozmózisnyomása pedig kisebb – hipoozmotikusak – a vérplazmáénál.
A sejtek meghatározott alakja az anyagcsere-folyamatok zavartalan lejátszódása miatt fontos, mivel a sejtek belső szerkezete, a sejtszervecskék optimális elrendeződése csak meghatározott sejtalak mellett valósul meg. A sejtek alakját az ozmózis révén alapvetően befolyásolják a testfolyadékok koncentrációviszonyai.
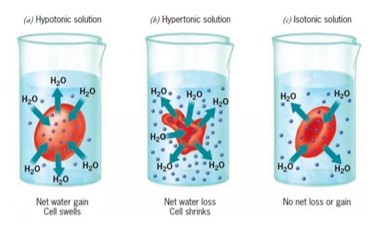
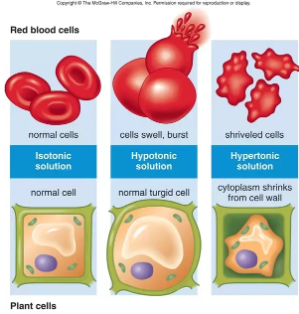
Ha kísérletképpen vörösvértesteket
- desztillált vízbe vagy hipotóniás oldatba teszünk – amelynek ozmózisnyomása jóval kisebb a sejtplazmáénál – a sejtekbe nagy mennyiségű víz áramlik, amelynek következtében a sejtek legömbölyödnek, majd kipukkadnak (hemolízis).
- Ellenkező esetben, ha a sejtet a sejtplazmánál töményebb – hipertóniás – oldatba helyezzük, a sejtek vizet veszítenek és összezsugorodnak.
- Izoozmotikus oldatokban nem lép fel ozmózis.
 |
 |
Élettani példák az ozmózisra
A növények életműködéseiben fontos szerepe van az ozmózisnak, a jelenség szerepet játszik
- a talajból történő vízfelvételben,
- a gázcserenyílások nyitásában-záródásában,
- a szerves anyagok áramlásában a háncs rostaelemeiben.
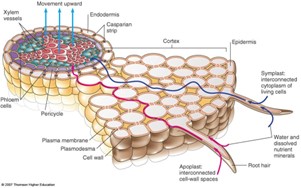
A víz és a sók felvételének módja
A gyökérszőrök sejthártyája féligáteresztő sajátosságú, azon a víz passzívan, ozmózissal beáramolhat, mivel a gyökérszőr sejtjeinek a citoplazmája töményebb, mint a talajoldat, így a sejtplazma egyfajta szívóerőt gyakorol a gyökérsejtek körüli vízmolekulákra. A beáramlott víz növeli a sejten belüli nyomást, turgor állapotot létrehozva, ezért a víz átpréselődik a szomszédos alapszöveti sejtekbe, majd onnan a farész vízszállító csöveibe (gyökérnyomást kialakítva).
A szükséges ionokat a gyökérszőrök sejthártyája válogatja ki a növény szükségleteinek megfelelően, majd a koncentráció viszonyoktól függően (esetleg passzív de) főleg aktív transzporttal a sejt belsejébe juttatja. Az energiaigényes transzporttal az ionok a nagyobb koncentráció irányába is szállítódhatnak.
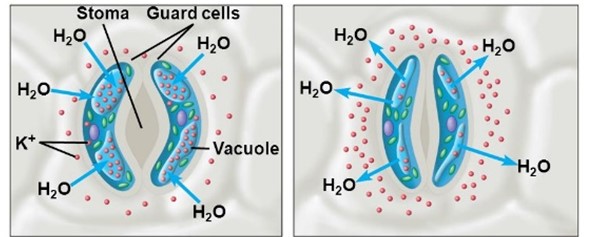
A gázcserenyílások működése
A zárósejtek speciális felépítésük révén a légrést tágítani vagy szűkíteni képesek. Az aktív sztómamozgás a zárósejtek vízzel való telitettségétől függ. Ha a zárósejtek turgora növekedik, a külső vékony falak erőteljesen megnyúlnak, kifelé görbülnek, amelynek következtében az egész zárósejt az epidermisz síkjában meggörbül, s a légrést körülvevő sejtfalak eltávolodnak egymástól, mire a légrés kinyílik.
- Amennyiben a zárósejtek K+-t és vizet vesznek fel és megduzzadnak, turgoruk nő, a légrés kinyílik,
- ellenkező esetben a vízvesztés következtében a zárósejtek összezáródnak.
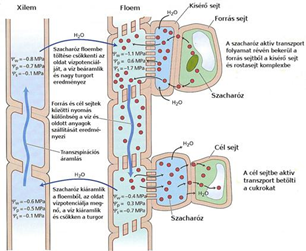
A szerves anyagok szállítása
Münch 1926-ban leírt nyomás-áramlási modellje szerint a forrás és a célállomás közötti eltérő ozmózisnyomás alapján működik a szállítás.
Eszerint a
- a levelek fotoszintézist végző sejtjeiből, aktív transzport szállítja a cukrot a rostacsövekbe,
- ahol emiatt nő az ozmózisnyomás.
- Ennek következtében a vízszállító csövekből víz áramlik a rostacsövekbe, ahol a beáramló víz miatt növekszik a turgornyomás.
- A felhasználás helyén, például a raktározószövetekben, a rostacsövekből szállítja aktív transzport a cukrot a környező sejtekbe,
- ami miatt csökken a rostacsövekben az oldat ozmotikus nyomása.
- Ennek hatására a víz kiáramlik a rostacsövekből, ami miatt csökken a turgornyomás. A rostacsövekben tehát az oldat a nagyobb nyomású hely felől a kisebb nyomású hely felé áramlik.
 |
 |
 |
Végezzük el a következő vizsgálatot. Vöröshagyma húsos alleveléből készítsünk nyúzatot, öt percre helyezzük 10%-os KCl-oldatba, majd mikroszkóp alatt vizsgáljuk a változást!
A látottakat az ozmózissal értelmezzük.
- Amennyiben növényi szöveteket magas koncentrációjú, ún. hipertóniás közegbe helyezünk, a sejtek vizet veszítenek, aminek következtében a sejt zsugorodik és a sejthártya elválik a merev sejtfaltól. A jelenséget plazmolízisnek nevezzük. A folyamat egy dinamikus egyensúlyi állapot elérése után megáll, hiszen a folyamat során a sejtben a vízvesztés miatt nő a koncentráció. Amikor a sejthártyán kívüli tér, ill. a sejten belüli tér ozmózisnyomása kiegyenlítődik, akkor a víz ki- és beáramlásának sebessége megegyezik, beáll az egyensúlyi állapot. A növények hervadása is akkor következik be, ha a sejtek a vízhiány miatt megváltozott koncentrációviszonyok mellett jelentősebb vizet veszítenek.
- Abban az esetben, ha a nyúzatot desztillált vízbe (hipotóniás közegbe) helyezzük, ozmózissal víz lép be a sejtplazmába (majd a sejtnedv-vakuólumba), a sejt vízzel telítődik, megduzzad, a citoplazma nyomása megemelkedik, nekifeszül a sejtfalnak, ami ellenáll a sejt térfogatnövekedésének. Ekkor a citoplazma sejtfalra gyakorolt nyomását turgornyomásnak nevezzük. A turgornak nagy szerepe van a növényi test alakjának fenntartásában, az anyagok szállításában, a gázcserenyílások működésének a szabályozásában, így a gázcserében, ill. a párologtatásban.
Az állati szervezetekben, és így az emberi szervezetben zajló vízmozgás irányának meghatározásában is óriási szerepe van az ozmotikusan aktív anyagoknak.
- A vese nefronjaiban a szűrletből jelentős vízvisszaszívás történik, 1 nap alatt közel 180 liter, ami annak köszönhető, hogy az elvezetőcsatornák, ill. a gyűjtőcsövek olyan szöveti környezetben futnak, amelynek az ozmotikus nyomása jelentősen meghaladja a szűrlet ozmotikus nyomását. A nagyobb ozmotikus nyomás kialakításában szerepet játszó ozmotikusan aktív anyagok pl. részben a szűrőműködés miatt megnövekedett koncentrációjú vérfehérjék, részben a sóvisszaszívás miatt megnövekedett koncentrációjú sók, elsősorban NaCl.
- A tápcsatornában is a víz passzív transzporttal szívódik fel, annak köszönhetően, hogy a béltartalom koncentrációja többnyire kisebb a bélhámsejtek citoplazmájának koncentrációjánál.
- A szövetek alapállományát képező szövetnedv a hajszálerek területén jön létre, mivel a vérerek közül kizárólag a kapillárisok fala átjárható a víz és a benne oldott kis molekulájú anyagok számára. A kapillárisok fala szűrőként működik, féligáteresztő, mivel átbocsátja a vizet, az ionokat, kisebb szerves molekulákat, pl. glükózt, aminosavakat, azonban visszatartja a nagyobb méretű vérfehérjéket és az alakos elemeket. A szűrés során lényegében fehérjementes vérplazma (szövetközti folyadék) jön létre.
A hajszálerek területén a folyadék mozgásának az irányát ellentétes hatások eredője szabja meg.
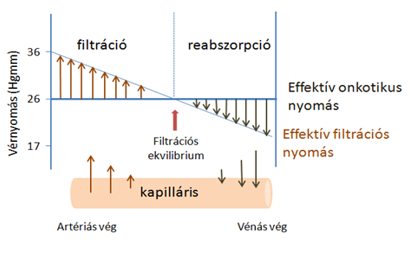
- Egyrészt az erekben uralkodó vérnyomás, amely a vérplazmát kifelé préseli,
- másrészt ennek ellene hat a plazmafehérjék kolloidozmotikus nyomása – ozmotikus szívóereje. A kapilláris artériás végén a filtráció miatt a plazmafehérjék koncentrációja és így az onkotikus nyomás kismértékben nő (1-2 Hgmm), de a változás csekély, ezért az élettani leírásokban gyakran állandónak tekintik.
 |
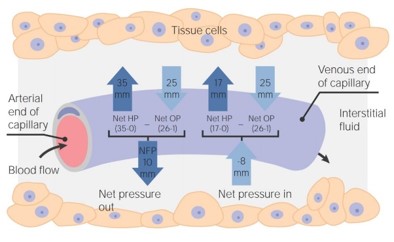 |
- A kapillárisok artériás szakaszán a vérnyomás nagyobb a kolloidozmotikus nyomásnál, ezért a vérplazma fehérjementes szűrlete kilép az érpályából (reverz ozmózis). A kapillárisokban a vénás oldal felé haladva a folyadék kilépése következtében a vér nyomása csökken.
- Amikor a vérnyomás egyenlővé válik a kolloidozmotikus nyomással, a folyadék kiszűrődése megszűnik.
- A kapillárisok vénás szakaszán a vérnyomás tovább csökken és a kolloidozmotikus nyomás alá esik. Ennek következtében a vénás szakaszon a vérfehérjék kolloidozmotikus szívóhatásának köszönhetően a szövetnedv egy része visszalép az érpályába, elszállítva a szövetekben keletkezett bomlásterméket. A szövetnedv oldott vérfehérjéket nem nagyon tartalmaz, ezért kolloidozmotikus nyomása alacsony.
Az ozmózis alkalmazása az orvosi gyakorlatban
- A különféle hatóanyagok, gyógyszerek szervezetbe juttatása történhet infúzió, ill. injekció útján is, melyek elkészítése során az összetevőket izotóniás – a vérplazmával megegyező ozmotikus koncentrációjú - fiziológiás sóoldatban oldják fel. Ez az eljárás az izotóniás környezetnek köszönhetően kivédi a szervezet sejtjeinek ozmotikus károsodását.
- A vér és a szövetnedv folyadék egyensúlyának zavara duzzanat, ödéma kialakulását eredményezi a szövetekben felhalmozódó felesleges szövetnedv miatt. Az ödéma kezelésére alkalmazható pl. dextrán oldat vagy keserűsó oldat (MgSO4), az eljárás során a duzzanat környezetében hipertóniás közeget teremtenek, ami vízelvonó hatású, ennek következtében a duzzanat csökken.
- A keserűsó alkalmazása hatékony lehet a székrekedés kezelésében A keserűsó kevéssé szívódik fel a tápcsatornából, s így a vastagbélben megnöveli a béltartalom ozmotikus koncentrációját, ez csökkenti a vízfelszívódás mértékét, ami viszont a béltartalom hígulását eredményezi a könnyebb üríthetőség érdekében.
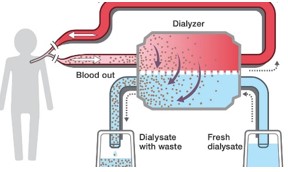
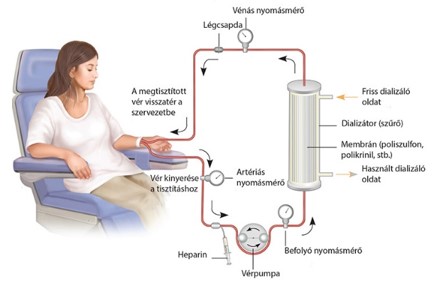
- A veseelégtelenségben szenvedő betegeknél alkalmazott művesekezelés szintén az ozmózis jelenségén alapszik. Az eljárást hemodialízisnek nevezzük. A beteg vérét egy szemipermeábilis csövön keresztül dializálógépbe vezetik, ahol a toxikus anyagokban gazdag vért tartalmazó csövek körül ún. steril dializáló folyadék áramlik. A berendezésben a mérgező salakanyagok a koncentrációkülönbségnek megfelelően kijutnak a vérből a féligáteresztő hártyán keresztül a dializáló oldatba, ugyanakkor a plazmafehérjék és a vér alakos elemei a vérben maradnak. Az ilyen módon megtisztított vért visszavezetik a testbe.
 |
 |
4. tétel: Az enzimek felépítése és működése |
Az enzimek felépítése és működése
A sejtekben lezajló biokémiai reakciók kötések bontásával, illetve létesítésével kapcsolatosak. A kémiai kötések felbontásához, a kémiai reakciók lejátszódásához energia szükséges, amit laboratóriumi körülmények között a hőmérséklet emelésével biztosítunk.
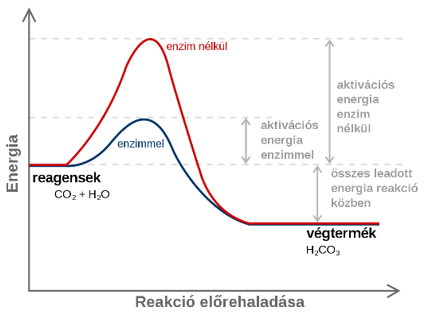
Ugyanakkor az élő szervezetek működésének körülményei között – hőmérsékleten, nyomáson – a kellő energia nem áll rendelkezésre a kötések felszakításához, a biokémiai reakciók maguktól nem mennek végbe. Ennek az az oka, hogy az élő rendszerekben a biomolekulák nem rendelkeznek annyi energiával – ún. aktiválási energiával –, amely a kötések felbontásához szükséges.
A reakciók végbemenetelét az ún. aktiválási gát akadályozza. Ennek ellenére a különféle reakciók zavartalanul lejátszódnak, mivel az élőlényekben biokatalizátorok, ún. enzimek működnek.
Általában a katalizátorok olyan anyagok, amelyek a kémiai reakciókat gyorsítják (a reakciósebességet növelik).
A biokatalizátorok úgy növelik a reakciósebességet, hogy a reakció számára egy másik, kisebb aktiválási energiával járó utat nyitnak meg.
Az enzimek működésének tehát az a jelentősége, hogy a reakciókat oly mértékben felgyorsítják, hogy azok a testünk hőmérsékletén is megfelelő sebességgel mennek végbe.
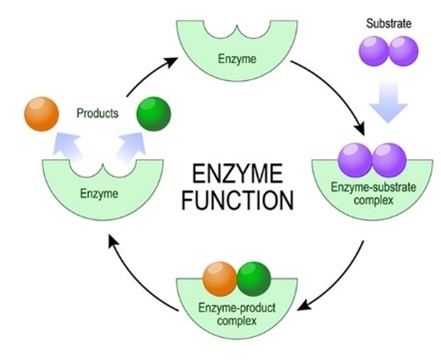
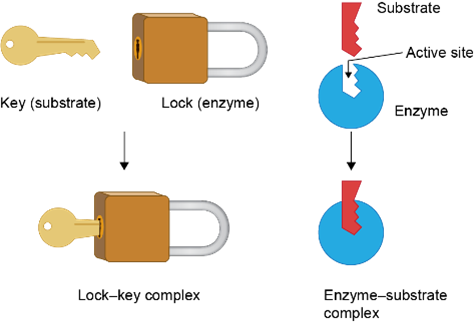
Az enzimek elsősorban globuláris fehérjék. Az enzimek felületén azt a helyet, ahol a katalizátorhatás történik, aktív centrumnak, vagy kötőzsebnek nevezzük. Az enzimek reakcióspecifikusak, fajlagosak, azaz a különböző reakciókat más-más enzimek katalizálják, mivel a kötőzseb felépítése specifikus, szerkezete kiegészítő – komplementer – annak az anyagnak – szubsztrátnak – a szerkezetével, amit átalakít (kulcs-zár modell).
 |
 |
 |
A biokémiai reakciók hőmérséklet-, ill. kémhatás-érzékenysége is az aktív centrum bonyolult szerkezetével függ össze, mivel már a hőmérséklet, ill. a kémhatás kismértékű megváltozásának hatására a kötőzseb elveszti a szubsztrátéval komplementer szerkezetét. Az enzimek optimális működéséhez tehát meghatározott környezeti feltételek – hőmérséklet, kémhatás, ionkoncentráció – szükségesek. Közismert például, hogy a magas láz életveszéllyel jár, mivel 41 °C felett annyira megváltozik az enzimek térszerkezete (még nem csapódnak ki!), hogy nem képesek ellátni biológiai feladatukat, ami miatt az egyes biokémiai folyamatok leállnak.
Az enzimek működésének hatékonyságát az enzimaktivitással jellemezzük. Az enzimaktivitás megmutatja, hogy adott idő alatt, adott mennyiségű enzim, adott feltételek mellett mennyi szubsztrátot képes átalakítani.
Az enzimaktivitás mértékét az élő szervezetekben különféle szabályozó mechanizmusok befolyásolhatják, hiszen amennyiben nincs szükség az enzim által katalizált reakcióra, annak további lefolyása még káros is lehet. Az enzimaktivitás mértékét csökkentő anyagokat inhibitoroknak nevezzük. Az inhibitoroknak többféle típusát ismerjük.
Egyik csoportjuk a versengő (kompetitív) inhibitorok, melyek a szubsztráttal való hasonló szerkezetük révén képesek az enzim aktív centrumába bekötődni, elfoglalva azt. Amennyiben az inhibitor jelen van, az enzim már képtelen a szubsztrát megkötésére, inaktívvá válik. Az enzimaktivitás mértékét a szubsztrát-, ill. az inhibitorkoncentráció viszonyai határozzák meg, hiszen ezek versengenek az aktív centrumban való megkötődés lehetőségéért.
Allosztérikus gátlás esetén az inhibitor az enzim térszerkezetét, azon keresztül is az aktív centrum szerkezetét változtatja meg úgy, hogy az enzim nem lesz képes megkötni a szubsztrátot.
Az enzimek csak átmenetileg vesznek részt a reakcióban, maradandó változást nem szenvednek, újra felhasználhatók.
Az egyszerű enzimek csak aminosavakból épülnek fel, ugyanakkor egyes, ún. összetett enzimek működéséhez gyakran szükséges valamilyen nem fehérje rész, ún. kofaktor.
A kofaktorokat az enzimhez kötődés erőssége alapján csoportosítjuk.
- A nem fehérjerész lehet ún. prosztetikus csoport, mely erősen kötődik a polipeptidlánchoz, eltávolítása a biológiai funkció megszűnésével jár. A prosztetikus csoport többnyire valamilyen fémion – pl. Cu2+, Zn2+, Fe2+ –, amely az enzim aktív centrumában a megfelelő térszerkezet kialakításához szükséges.
- Egyes enzimek működéséhez pedig ún. koenzimek szükségesek. A koenzimek olyan nem fehérjetermészetű vegyületek – kofaktorok –, többnyire nukleotidszármazékok, melyek lazán, reverzibilis módon kötődnek az enzimekhez. A koenzimek általában szállítómolekulák, a szubsztrátok közötti csoportátviteli reakciókban szerepelnek, mint pl. NAD+, NADP+, melyek hidrogénatomok átvitelében játszanak szerepet. A koenzimek felépítésében gyakran vesznek részt vitaminjellegű csoportok, ezért a vitaminhiány gátolja a koenzimek felépülését, ezen keresztül egyes biokémiai folyamatok lejátszódását. A NAD+ felépítésében pl. a B3-vitamin vesz részt.
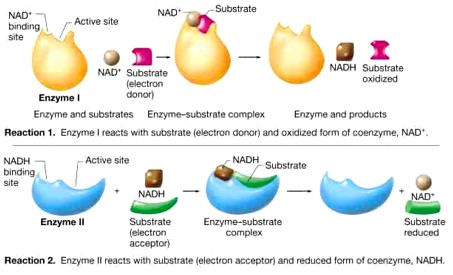 |
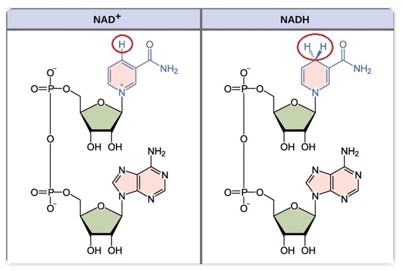 |
Enzimhiányon alapuló emberi betegségek
Számos betegség kialakulásának a hátterében az anyagcserében szerepet játszó valamilyen enzim hiánya vagy elégtelen működése áll. Ilyen többek között a tejcukor-érzékenység és a fenilketonuria.
Tejcukor-érzékenység (laktózintolerancia)
A táplálékintolerancia egy olyan, többnyire az emésztőszervrendszert érintő tünetegyüttes, amelyet egyes ételek adott összetevője vált ki. A tüneteket az anyag emésztési, ill. felszívódási zavara okozhatja. Nem érinti az immunrendszert, szemben a táplálékallergiával.
Tejcukor-érzékenység esetén a tejcukor (laktóz) vált ki hasi panaszokat, puffadtságot, hasmenést, émelygést. A tünetek hátterében a tejcukrot bontó enzim, a laktáz hiánya vagy elégtelen működése áll. A vékonybélben a tejcukor nem tud monoszacharidokká bomlani, így nem szívódik fel és a vastagbélbe kerülve különféle problémákat okoz.
A vastagbélbe kerülő laktóz
- erősen növeli a béltartalom ozmotikus koncentrációját, szívóerejét, miáltal csökkenti a vízfelszívódás mértékét,
- továbbá a vastagbélben zajló bakteriális erjedés következtében különféle szerves savak és gázok – H2, CO2 – keletkeznek, melyek erőteljesen fokozzák a bélperisztaltikát.
Mindezek eredményeképpen a széklet jelentősen hígul, ami elsődleges oka a hasmenésnek. A kialakuló tünetek mértékét a laktózbevitel csökkentésével lehet mérsékelni pl. laktózmentes tejtermékek fogyasztásával.
A fenilketonuria lásd még 6.1. és 6.2. fejezet
A fenilketonuria autoszómás, recesszíven öröklődő, enzimhiányon alapuló szervezetszintű anyagcserezavar. A tüneteket az okozza, hogy
- egyrészt a betegek mája a fenilalanin-hidroxiláz enzim hiánya miatt nem tudja lebontani a – nagyobb mennyiségben mérgező hatású, egyébként esszenciális – fenilalanin aminosavat, ezért az veszélyes mértékben felszaporodik a vérben.
- Másfelől a felhalmozódó fenilalanint a szervezet alternatív anyagcsereutakon próbálja átalakítani, melynek során szintén különféle mérgező anyagcseretermékek keletkeznek.
A túlzott mennyiségben jelen lévő fenilalanin és egyéb anyagcseretermékek pedig idegrendszeri károsodást, értelmi fogyatékosságot okoznak.
Kezelése a fenilalanin bevitelének kontrollálásával, fehérjeszegény diétával történik. A fenilketonuriásnak tilos húsféléket, tejtermékeket, hüvelyeseket, tojást, szóját ennie.
Néhány példa a sejtek energiaátalakításával kapcsolatos enzimek működésére
Egyes enzimek, pl. ún. ATP-ázok (ATP bontó enzimek) képesek az ATP nagy energiájú kötéseiben rejlő kémiai energiát különféle folyamatokban hasznosítani.
Na+-K+- pumpa (ATP-áz) lásd még 2.3. fejezet
Az állati sejtek legfontosabb ionpumpája. Az enzim működése során 1 ATP hidrolízisével 3 Na+-t pumpál ki a sejtből és 2 K+-t visz be a sejtbe, ezzel létrehozva a sejthártya két oldalán az ionkoncentrációk különbségét. Működése tehát energiaigényes, mivel az ionmozgások a koncentrációgradienssel ellentétesek.
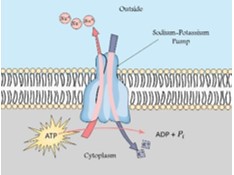 |
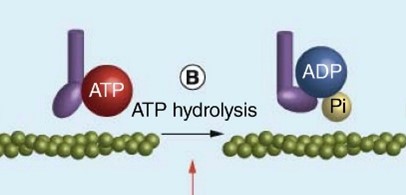 |
Miozin lásd még 4.3. fejezet
Az izomszövetekben jellemzően megtalálható miozin fehérje szintén ATP bontó enzimhatással rendelkezik, tekintve, hogy az izomösszehúzódás energiaigényes folyamat. Az összehúzódás során az ATP bontásakor felszabaduló energia a miozin szerkezetváltozásához szükséges, ami elengedhetetlen a fehérjefonalak egymás melletti elcsúszásához.
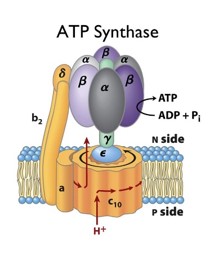
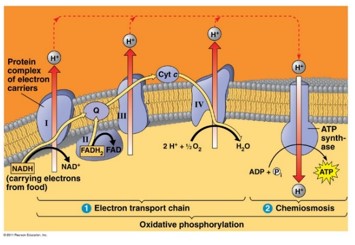
ATP-szintázok lásd még 2.3. fejezet
Az ATP-szintázok olyan enzimek, amelyek bizonyos folyamatokban felszabaduló energiát képesek ATP előállítására fordítani. Az ATP egyébként a lebontó anyagcsere során keletkezik, pl. a glikolízisben (szubsztrát szintű foszforiláció), ill. a terminális oxidációban (oxidatív foszforiláció).
A terminális oxidáció egy elektronszállító rendszer működésével valósul meg, mely a mitokondriumok belső membránjában található.
- A lebontó anyagcserében a szubsztrátokról leváló hidrogének NADH molekulák közvetítésével kerülnek a mitokondrium belső membránjához.
- Az elektronszállító rendszer elején a protonok és az elektronok szállítása különválik.
- Az elektronok bekerülnek az elektronszállító rendszerbe, ahol áramlásuk eredményeképpen a felszabaduló energia protonok aktív transzportjára fordítódik a mitokondrium belső teréből a külső kamrába.
- Ennek eredményeképpen a külső kamrában jelentősen megnő a protonkoncentráció, így a membrán két oldalán protonkoncentráció különbség alakul ki.
- A különbség kiegyenlítődését az alapállomány felé néző enzimkomplex, az ATP-szintáz végzi. Az enzimkomplexen a protonok átáramlásakor felszabaduló energia ATP szintézisre fordítódik (oxidatív foszforiláció).
 |
 |
5. tétel: A víz biológiai szempontból fontos jellemzői |
Alapfogalmak, alapismeretek
Az oldatok két vagy több komponensből álló összetett homogén anyagi rendszerek. A homogén rendszerekben nincsenek fázishatárok, azaz szabad szemmel vagy fénymikroszkóppal látható határfelületek. A komponensek a halmaz kémiailag különböző összetevői. Az oldatok minimum két komponensből állnak,
- a nagyobb mennyiségben jelenlevő, többnyire folyékony összetevő az oldószer,
- a másik alkotórész az oldott anyag.
Az élő rendszerek működése szempontjából a folyékony halmazállapotú oldatoknak van jelentősége, ahol az oldószer a víz, az oldott anyagok lehetnek ionok, gázok és különféle szerves vegyületek.
A vizes oldatokban az oldószer molekulái mozoghatnak szabadon, de kötődhetnek az oldott anyag részecskéihez is, mint pl. az ionok, ill. különféle poláris molekulák felületén található hidrátburok vízmolekulái. A hidrátburokban adszorpciósan kötött víz nehezen eltávolítható a részecskék felületéről, a különféle élettani folyamatokban nem hasznosítható.
Az oldatok összetételét, az oldott anyag mennyiségét, többek között a koncentrációval lehet kifejezni. Biológiában leggyakrabban az oldatok összetételének a jellemzésére az anyagmennyiség-koncentrációt használjuk, ami az oldott anyag anyagmennyiségének és az oldat térfogatának hányadosa, jele c, mértékegysége mol/dm3.
Az oldódás során az oldószer és az oldott anyag részecskéi elkeverednek egymással. Az elegyedés lehet
- pusztán fizikai folyamat, amikor a részecskék között kémiai reakció nem történik, ilyen pl. az oxigéngáz oldódása vízben,
- de az oldódást kísérheti kémiai változás is, mint pl. amikor a szén-dioxid gáz vízben oldódva szénsavvá alakul.
A vízmolekulák dipólusos szerkezetűek, s emiatt a vizet poláris oldószernek tekintjük. A vízmolekulák között H-kötések találhatók, ezért a víznek, mint folyadéknak magas az olvadás- és forráspontja, nagy a hőkapacitása, nagy párolgáshő jellemzi, továbbá fagyásakor a halmaz térfogata nő. A felsorolt tulajdonságoknak az élő rendszerek működésében van jelentősége.
Az oldott anyagok gyakran nem képesek korlátlan mértékben elegyedni az oldószerrel, csak maximum a telítettség eléréséig, az ún. oldhatóságuknak megfelelően. Az adott anyag oldhatóságát a telített oldatának összetételével jellemezzük. A telített oldatban az oldószer nem tud több oldott anyagot feloldani, az oldódás és a meginduló kiválás folyamata ún. dinamikus egyensúlyt alakít ki. Általános értelemben dinamikus egyensúlyban az ellentétes irányú folyamatok – jelen esetben az oldódás és a kiválás – sebessége megegyezik.
A vizes oldatok kémhatását a hidrogénionok (H+ (H3O+)) és a hidroxidionok (OH-) koncentrációjának viszonya szabja meg.
- A savas kémhatású oldatokban a hidrogénionok koncentrációja nagyobb, mint a hidroxidionok koncentrációja,
- a semleges kémhatású oldatokban a hidrogénionok koncentrációja megegyezik a hidroxidionok koncentrációjával,
- a lúgos kémhatásúoldatokban pedig a hidroxidionok koncentrációja nagyobb, mint a hidrogénionok koncentrációja.
Az oldatok kémhatását számszerűen a pH-érték megadásával lehet jellemezni. A pH a H+-koncentráció hatványkitevőjéből adható meg (H+-koncentráció tízes alapú logaritmusának mínusz egyszerese). A pH-skála 0–tól 14-ig terjed.
- A semleges oldatok pH-ja 7.
- Ha a pH értéke kisebb, mint 7, akkor az oldat savas, pl. a 0,1 mol/dm3-es sósav pH-ja 1, mivel [H+] = 10-1 mol/dm3.
- Ha a pH érték nagyobb, mint 7, akkor az oldat lúgos.
Meghatározó élettani folyamatok hátterében, a vízzel kapcsolatosan két fontos fizikai-kémiai folyamat áll, a diffúzió és az ozmózis (lásd 3. tétel).
 |
 |
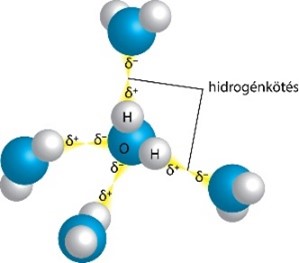 |
A víz
Az élőlények számára a legjelentősebb szervetlen vegyület a víz.
A vízmolekula poláris, dipólusos szerkezetű, az oxigénatomon elektronfelesleg – negatív töltéstúlsúly –, a hidrogénatomokon pedig elektronhiány – pozitív töltéstúlsúly - alakul ki, tekintve, hogy az oxigén nagyobb elektronvonzó képessége – elektronegativitása – folytán a kötő elektronpárokat maga felé vonzza.
A poláris vízmolekulákat H-kötés tartja össze, mely az egyik oxigénatom nemkötő elektronpárja és a másik vízmolekula hidrogénatomja között létesül. A folyékony víznek, mint anyagi halmaznak a sajátos fizikai tulajdonságai – melyek a földi élet szempontjából különös jelentőséggel bírnak - a molekulák közötti H-kötésre vezethetők vissza, mint pl.
- a molekulatömegéhez képest magas olvadás- és forráspont,
- a nagy hőkapacitás,
- a nagy párolgáshő,
- továbbá, hogy fagyásakor a halmaz térfogata nő.
A víznek molekulatömegéhez képest magas az olvadáspontja (0 oC) és forráspontja (100 oC), aminek köszönhetően a természetben mindhárom halmazállapotban előfordul. Figyelembe véve az életfolyamatok működési hőmérséklet-tartományát, a földi élet folyékony víz nélkül nem jöhetett volna létre.
Hőkapacitásnak nevezzük az adott tömegű anyag hőmérsékletének 1 °C-al való emeléséhez szükséges hőt. A különböző minőségű anyagoknak eltérő a hőkapacitása, azaz ugyanakkora hőmérséklet-változás létrejöttéhez különböző hőmennyiségre van szükségük.
A víznek nagy a hőkapacitása, ezért nagyon sok hőt képes elnyelni, tárolni és szállítani. Ennek jelentősége abban áll
- egyrészt, hogy klímameghatározó szerepe van, mivel a nagyobb víztömegek lassabban és kevésbé melegszenek fel, valamint lassabban és kevésbé hűlnek le, mint a szárazföldi területek, ill. a víz a nyáron elnyelt hőmennyiséget télen bocsátja ki, tehát télen fűt, nyáron hűt (óceáni éghajlat),
- másrészt pedig, hogy az élőlények testhőmérséklete a magas víztartalmuk miatt kevésbé ingadozik, akár a külső hőmérsékletváltozás okán, akár a hőtermelő (exoterm) anyagcsere folyamatok végett.
A párolgáshő egységnyi tömegű folyadék állandó hőmérsékleten történő elpárologtatásához szükséges energia. Nagy párolgáshőjének köszönhetően a víz sok hőt von el a környezetétől párolgása során, aminek többek között az élőlények – növények, állatok egyaránt – hőszabályozásában van jelentősége.
 |
 |
 |
A víz sűrűsége + 4°C-nál a legnagyobb, további hűlésekor sűrűsége már csökken, így fagyáskor térfogata nő, aminek megfelelően a tavak nem alulról, hanem felülről fagynak be.
Télen a természetes édesvizek befagyás előtt teljes mennyiségükben + 4°C-ra hűlnek le, s csak majd ezt követően fagy meg a legfelső vízréteg, ami a vízi élővilág számára rendkívül kedvező.
A víznek nagy a felületi feszültsége, ami miatt lehetőség szerint a legkisebb fajlagos felületű (tömeghez viszonyított felület) gömb alakot veszi fel, ill. a vízszintes folyadék felülete rugalmas hártyához hasonlóan viselkedik.
A vízzel kapcsolatos jelenség az ún. kapilláris hatás (hajszálcsövesség). Amennyiben egy folyadék és egy szilárd anyag részecskéi között nagyobb a vonzóerő (adhézió), mint a folyadék molekulái közötti kohézió, kapilláris emelkedés következik be, a víz keskeny térben a nehézségi erő ellenében is képes mozogni, a folyadék úgymond nedvesíti az edény falát.
A víz felületi feszültségének sokrétű a biológiai jelentősége:
- egyes rovarok képesek a víz felszínén futkározni, annak ellenére, hogy a sűrűségük nagyobb a vízénél (nem süllyednek el).
- A hajszálcsövességnek, ill. a vízmolekulák között ható kohéziós erőnek fontos szerepe van a növények vízszállító edényeiben a víz szállításában, ill. a talajban a felfelé irányuló vízmozgásban.
A víz biológiai szerepe
Kitűnő poláris oldószer
- Poláris vegyületeket hidratációval,
- ionrácsos vegyületeket elektrolitos disszociációval,
- amfipatikus vegyületeket micellaképződéssel oldja.
Általában mindazon vegyületek jól oldódnak vízben, amelyek H-kötésre képesek.
Reakciópartner
- Hidrolízis során egy nagyobb molekula víz belépésével kisebb molekulákra bomlik. Makromolekulák monomerekre bomlásában van jelentősége, pl. az emésztésben.
- Kondenzáció során kisebb molekulák víz kilépésével nagyobb molekulákká egyesülnek. Monomerek makromolekulákká összekapcsolódásában jelentős, pl. fehérjeszintézisben.
Reakcióközeg, mivel az anyagcsere-folyamtok vizes oldatokban játszódnak le.
Szállítóközeg, vízben oldott anyagokat szállítja (pl. vér).
Szerkezetmeghatározó
- A sejtek alakjának,
- növényi sejtek turgorának,
- lágyszárú növények, puhatestűek testalakjának meghatározója.
6. tétel: A szénhidrátok biológiai szempontból fontos jellemzői |
A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek.
A szénhidrátok biológiai jelentősége:
- a sejtek elsődleges energiaforrásai (glükóz),
- tartaléktápanyagok (keményítő, glikogén),
- vázanyagok (cellulóz, kitin, pektin),
- más vegyületekkel összekapcsolódva fontos makromolekulák – nukleinsavak – építőkövei (ribóz, dezoxiribóz).
Többféleképpen csoportosíthatók.
Méret alapján:
- egyszerű szénhidrátok, ill. monoszacharidok, amelyek savas hidrolízissel tovább nem bonthatók,
- összetett szénhidrátok, savas hidrolízissel monoszacharidokra bonthatók,
- diszacharidok, két monoszacharidból,
- poliszacharidok, sok száz, ill. ezer egyszerű szénhidrátból épülnek fel.
Funkciós csoport alapján:
- aldózok (a) aldehidcsoportot,
- ketózok (b) ketocsoportot tartalmaznak.
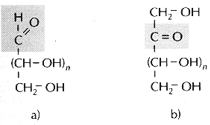 |
Monoszacharidok
Általában
- édes ízű,
- fehér, kristályos,
- nem hidrolizálható,
- vízben jól oldódó anyagok.
- Összetett szénhidrátok emésztésének végtermékei.
További csoportosításuk szénatomszám szerint történik.
Triózok (C3)
A legegyszerűbb monoszacharidok, ilyen pl. a glicerinaldehid, mely az anyagcserében fontos köztes termék.
Pentózok (C5)
- Ribóz (RNS-ben, ATP-ben, koenzimekben)
- Dezoxiribóz (DNS-ben)
A kétféle pentóz között csupán egyetlen oxigénatom a különbség. Mint az elnevezés mutatja, a dezoxiribóz 2. szénatomjához nem kapcsolódik oxigén, csupán hidrogén.
A pentózok jelentősége:
- DNS, RNS építőkövei,
- az anyagcserében köztestermékek.
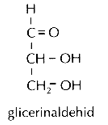 |
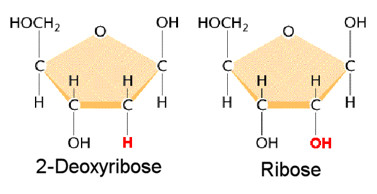 |
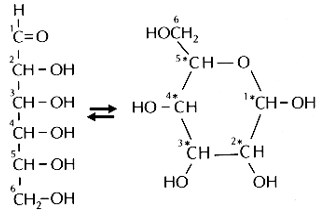 |
Hexózok (C6)
Glükóz, szőlőcukor C6H12O6
A legfontosabb monoszacharid:
- a legelterjedtebb, legnagyobb mennyiségben fordul elő,
- fotoszintézis sötét szakaszának végterméke,
- a vérben a szénhidrátok szállítása ebben a formában történik,
- makromolekulák - keményítő, cellulóz stb. -monomerje,
- a sejtek elsődleges, közvetlen energiaforrása, a vérben literenként kb. 1g glükóz van.
- Édes ízű, vízben kitűnően oldódó anyag.
- Összetett szénhidrátok emésztésének a végterméke, tovább nem emészthető.
Vizes oldatban a molekulák kb. 1%-a nyílt láncú, 99%-a gyűrűs szerkezetű. Gyűrűs állapotban a glükóznak kettő stabil izomerje létezik.
- Béta-glükóz (63%),
- alfa-glükóz (36%).
A béta-glükóz
A gyűrűs állapotú székkonformációban a C-atomokhoz kapcsolódó összes -OH-csoport ekvatoriális állású. Ez a konformáció a lehető legstabilabb glükózizomer.
Az alfa-glükóz
A gyűrűs állapotú székkonformációban az első szénatomon az -OH-csoport axiális állású, a többi C- atomon ekvatoriális.
A kétféle izomer vizes oldatban a nyitott formán keresztül átalakul egymásba, egyensúlyt fenntartva.
A glükóz kimutatása
A szénhidrátokat a reakcióképes aldehid-csoporton keresztül lehet kimutatni. A reakció lényege, hogy az aldehidcsoport megfelelő körülmények között karboxil-csoporttá oxidálódik, miközben a reagens anyagok – pl. ezüstionok - színváltozás kíséretében redukálódnak.
Ezüsttükör-próba (Tollens próba)
Az ezüstionokat lúgos közegben az aldehidcsoport fém ezüstté redukálja, amely kiválva az oldatból bevonatot képez az edény falán.
 |
 |
 |
Fruktóz, gyümölcscukor
Főleg gyümölcsökben (elnevezés), mézben előforduló, legédesebb ízű monoszacharid. Összegképlete C6H12O6, megegyezik a glükózéval, konstitúciós izomerek, aldehidcsoport helyett ketocsoportot tartalmaz nyílt láncú formában.
Diszacharidok
Két monoszacharid kondenzációja révén jönnek létre. A monoszacharidok között éterkötés (glikozidkötés) található.
A mono- és a diszacharidokat cukroknak nevezzük, többnyire élelmiszerekben fordulnak elő, pl. glükóz, fruktóz, maltóz, szacharóz, laktóz.
Maltóz, malátacukor
Fehér színű, vízben jól oldódó, édes ízű por. Két alfa-glükózból épül fel. Az egyik α-D-glükóz 1-es szénatomjának hidroxilcsoportja és egy másik α-D-glükóz 4-es szénatomjának hidroxilcsoportja között történik a kondenzáció.
Elsősorban ott fordul elő, ahol előzőleg keményítő bontása folyt, így pl. csírázó magvakban (maláta = csírázó árpa), zöld levelekben, emésztéskor a tápcsatornában. A glikogén bontásakor szintén felszabadul.
Laktóz, tejcukor
A laktóz egy β-D-glükóz és egy β-D-galaktóz kondenzációjával jön létre. Amennyiben a tejcukor a vékonybélben nem tud egyszerű cukrokká bomlani - a laktózt bontó enzim, a laktáz hiánya miatt -, ún. tejcukor-érzékenység alakul ki, mely jellemző tünetei a különféle hasi panaszok, mint pl. puffadtság, hasmenés, émelygés. A panaszok hátterében
- egyrészt az áll, hogy a bélben maradó laktóz erősen növeli a béltartalom ozmotikus szívóerejét, rontva a vízfelszívódás hatékonyságát, ami a széklet hígulását eredményezi,
- másrészt a vastagbélben működő baktériumok működésének köszönhetően különféle szerves savak és gázok keletkeznek, melyek jelentősen fokozzák a bélperisztaltikát.
Szacharóz, répacukor, nádcukor
Fehér színű, vízben jól oldódó, édes ízű por. A növényvilágban az egyik legelterjedtebb, szabadon előforduló diszacharid. Az α-D-glükóz 1-es szénatomján lévő hidroxilcsoport és a β-D-fruktóz 2-es szénatomján lévő hidroxilcsoport között történik a kondenzáció.
Az emberi táplálkozás és az élelmiszer-előállítás szempontjából a legjelentősebb szénhidrát, emberi étrendben a legfontosabb természetes édesítőszer, a közönséges cukor alapanyaga. Neve mutatja, hogy cukorrépából, ill. cukornádból állítják elő.
 |
 |
Poliszacharidok
A poliszacharidok monoszacharidokból kondenzációval felépülő óriásmolekulák, polimerek. Sok száz vagy akár több ezer monoszacharid kapcsolódhat egymáshoz. Savas hidrolízissel általában előbb diszacharidokká, majd monoszacharidokká bonthatók. A legelterjedtebb szénhidrátok. Nem édes ízűek. Keletkezésük:
nC6H12O6 → (C6H10O5)n + (n–1)H2O
Általános képletük:
Feladatuk szerint csoportosítjuk:
- tartalék tápanyagok: keményítő, glikogén,
- szilárdító vázanyagok: cellulóz, kitin.
Keményítő
Hideg vízben nem, meleg vízben kolloidálisan oldódó, nem édes ízű fehér por. A növényekben keletkezik a fotoszintézis eredményeképpen, tartalék tápanyag. A keményítő több száz alfa-glükózmolekulából épül fel.
Emészthető, emésztése már a szájüregben megkezdődik, majd a vékonybélben fejeződik be, a nyál-, ill. a hasnyálamiláz enzim hatására, amely maltózra hidrolizálja.
A keményítő hidrolízis hatására glükózra esik szét.
Ha a kenyeret sokat rágjuk, előbb-utóbb édes ízt fogunk érezni, mivel a liszt keményítőtartalma ugyan nem édes, de a belőle keletkezett maltóz már igen.
Növényi magvakban a keményítő hidrolízisét szintén az amiláz enzim végzi.
A növényekben, a raktározó alapszövet sejtjeiben, fajra jellemző módon szemcsékben jelenik meg. A szemcsék kétféle szerkezetű keményítőből állnak.
 |
 |
 |
Amilóz
Az amilóz egy elágazásmentes, spirálisan feltekeredett lánc. A spirált H-kötések stabilizálják.
Amilopektin
Ágas-bogas szerkezetű.
A keményítő kimutatása
- Por keményítőből melegítéssel készítsünk keményítőoldatot.
- Híg oldatba cseppentsünk 1-2 csepp kálium-jodidos jódoldatot – Lugol-oldatot.
- + próba esetén az oldat megkékül.
- Melegítés hatására elszíntelenedik, visszahűtéskor újra kék lesz.
Magyarázat
Az apoláris jódmolekulák a poláris vízben barna színűek. A jódmolekulák mérete olyan, hogy pont beleférnek az amilóz spiráljába, onnan kiszorítva a vízmolekulákat, apoláris közeget hozva létre. Az apoláris közegben a jód színe kékre változik.
Melegítés hatására a hőmozgás miatt a jódmolekulák kibújnak a spirálból a kék szín elhalványodása kíséretében. Hűtéskor a folyamat ellenkező irányú.
 |
 |
 |
Glikogén
Heterotróf szervezetek - állatok, gombák - tartalék szénhidrátja. Állati szervezetekben elsősorban a májban és az izomban raktározódik. Alfa-glükózokból felépülő, többszörösen elágazó, ágas-bogas szerkezetű. Nem édes ízű, vízben rosszul oldódik. Emészthető, szintén amiláz hatására hidrolizál maltózzá.
Cellulóz
A biomassza tömegének kb. a felét a cellulóz adja! A növényi sejtfal szilárdító anyaga (ezenkívül moszatgombákban fordul elő).
Óriásmolekula, sok ezer béta-glükózmolekulából áll. A cellulózban a glükózmolekulák egy elágazásmentes polimert hoznak létre. A glükózgyűrűk egymáshoz képest felváltva 180 fokot elfordulnak. A cellulózmolekula egyenes, szálas szerkezetű.
Rendkívül stabil, vízben nem oldódik, ami lehetővé teszi biológiai feladatának - szilárdítás - ellátását. Rostos szerkezetű, a láncok egymással párhuzamosan rendeződnek, a glükózmolekulák OH-csoportjai között H-kötések vannak, a láncon belül és a láncok között egyaránt.
A cellulózt a legtöbb élőlény nem képes bontani, így nem emészthető. A cellulózt bontó enzim a celluláz, amely csak baktériumokban, gombákban, ill. egyes egysejtűekben fordul elő. Hidrolízisekor végső soron glükóz keletkezik.
Kitin
N-tartalmú poliszacharid. Ízeltlábúak kutikulájának, gombák sejtfalának szilárdító vázanyaga. Rendkívül ellenálló, vízben nem oldódik, nem emészthető.
 |
 |
7. tétel: A fehérjék biológiai szempontból fontos jellemzői |
A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50%-át adják. Csoportosításuk biológiai feladataik alapján történik, lehetnek:
- vázanyagok, szerkezeti fehérjék: tartó, szilárdító feladatokat látnak el, pl. a legtöbb állati kötőszövetben, a csontban a kollagén, keratin az elszarusodott hámokban, hajban, tollban, hüllők pikkelyeiben (lásd hámszövetek),
- összehúzékony fehérjék: ilyen az aktin, miozin, pl. az izmokban (lásd izomszövetek),
- transzportfehérjék: szállító feladatokat látnak el, pl. a hemoglobin oxigént szállít, ill. a vérben a globulinok zsírokat, vasat, hormonokat szállítanak (lásd vér),
- védőfehérjék, immunfehérjék (antitestek): fertőzésekkel szembeni védekezésben közreműködnek, pl. az immunoglobulinok a vérben (lásd vér, immunitás),
- véralvadásban közreműködő fehérjék, pl. trombin, fibrin stb. (lásd vér),
- szabályozó fehérjék, mint pl. a hormonok: kémiai jelek, szervek, szövetek működését befolyásolják, pl. inzulin (lásd hormonrendszer),
- receptorok a sejtek felszínén, melyek különféle anyagokat ún. ligandumokat – pl. hormonokat - képesek megkötni (lásd biológiai membránok),
- ozmotikus faktorok az albumin a vérben, mely a vér optimális ozmotikus koncentrációjának (ozmotikus nyomásának) kialakításában szerepel (lásd anyagszállítás jegyzet),
- jelölőfehérjék (markerek) a sejtek felszínén elhelyezkedő egyed-, szövet-, ill. fajspecifikus molekulák (MHC), melyeknek a sejtfelismerésben (saját-idegen), így az immunrendszer működésében látnak el alapvető szerepet (lásd vér, immunrendszer),
- tartalék tápanyagok egyes növényi magvakban, gabonafélék szemtermésében ilyen az aleuron, ill. a búza szemtermésében a glutén, vagy a tojásfehérje továbbá a kazein a tejben,
- enzimek: biokatalizátorok, a sejtekben zajló kémiai folyamatok aktiválási energiáját csökkentik, aminek következtében az átalakulások reakciósebessége megnő. Az emberi szervezet működési körülményei között, katalizátorok nélkül az életfolyamatok végtelen lassú sebességgel mennének végbe (lásd 1.2. fejezet).
Az aminosavak
A fehérjék makromolekulák, monomerjeiket aminosavaknak nevezzük. Kémiailag amino-karbonsavak, azaz a molekulában két eltérő jellegű funkciós csoport is megtalálható:
- bázisos aminocsoport,
- savas karboxilcsoport.
Minden aminosav egy azonos, és egy eltérő molekularészletből áll:
- az azonos rész tartalmazza az amino-, és a karboxilcsoportokat,
- az eltérő rész az ún. oldallánc, amely szerkezetileg 20 (21)-féle Az oldallánc lehet poláris, apoláris, savas, bázisos jellegű.
Az aminosavak egy részét a szervezet nem, vagy csak elégtelen mennyiségben képes előállítani, ezeket esszenciális aminosavaknak nevezzük, melyeket a táplálékkal kell felvenni. Emberben 9 ilyen aminosav ismert, mint pl. a fenilalanin, lizin, metionin, valin stb.
Amfoter vegyületek, azaz savként és bázisként egyaránt viselkedhetnek a reakciópartnertől függően, azonban sav-bázis sajátságaikat az oldallánc kémiai természete is befolyásolja.
 |
 |
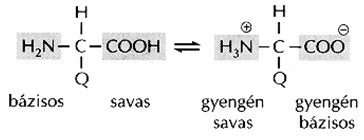 |
Biológiai szempontból legfontosabb reakciójuk a kondenzáció, melynek során az egyik aminosav aminocsoportja, és a másik aminosav karboxilcsoportja között vízkilépéssel, ún. peptidkötés jön létre (amidcsoport). A reakció eredményeként a két aminosavat egy síkalkatú amidcsoport köti össze.
Peptidek
Az aminosavak összekapcsolódásával ún. peptidek jönnek létre. A polipeptidek néhány 100 aminosavból álló elágazásmentes molekulák.
A polipeptidlánc aminosavsorrendjét szekvenciának nevezzük.
Szekvencia
Általában a makromolekulák monomerjei – így pl. az aminosavak - viszonylag egyszerű felépítésűek és minden élőlényben azonosak az élővilágban. A fajokra jellemző egyediséget, különbséget a fehérje (és nukleinsav) monomerek sorrendje határozza meg.
Másképpen fogalmazva, a természetben a fajok tekintetében tapasztalható óriási változatosságot nem a monomerek sokféleségében kell keresnünk, hanem a nukleinsav és fehérje monomerek összekapcsolódásának változatosságában, sorrendjében. Ebben az esetben tehát a monomerek kapcsolódási sorrendjének információt meghatározó jelentősége van, amely információ a fajok egyediségének meghatározója. Számszerűen, pl. a fehérjéket 20-féle monomer – aminosav – építheti fel, a véletlenszerű összekapcsolódásuknak megfelelően
- 2 aminosav kétféleképpen kapcsolódhat egymáshoz, attól függően, hogy melyik helyezkedik el az N-terminálison (NH2-csoport végen).
- 3 aminosavat már 6-féle sorrendben kapcsolhatunk össze.
- 100 db - 20-féle - aminosavból már 20100-féle polipeptid alkotható.
Ez a hatalmas variációszám hatalmas mennyiségű információt hordozhat. További változatosságot eredményez, hogy a különböző makromolekulák eltérő méretűek, mérettartományuknak elvi felső határa nincs, ami azt jelenti például, hogy elvileg végtelen számú fehérjemolekula-féleséget tudunk elképzelni.
Az élővilág összes faja azonos alkotóelemekből, azonos elvek alapján hoz létre fajra jellemző makromolekulákat. A monomerek sorrendje információt hordoz, az egyes elemek kicserélése megváltoztatja az információ jelentését.
A szekvencia döntően meghatározza a fehérjék tulajdonságait, ezért az aminosavsorrendet a fehérjék elsődleges szerkezetének nevezzük.
Akár egyetlen aminosav helyének megváltoztatása az egész fehérje működésére hatással lehet.
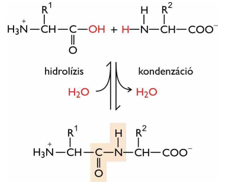 |
 |
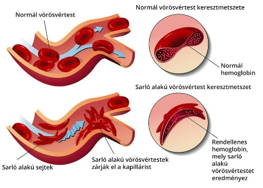 |
Sarlósejtes vérszegénység
A szekvencia jelentőségét jól tükrözi a sarlósejtes vérszegénység. A rendellenesség nevét onnan kapta, hogy a betegek vérében lévő - az egyébként korong alakú – vörösvértestek sarló formájúak. A hibás vörösvértestek vérrögöket képezve kiszelektálódnak, aminek következtében csökken a vörösvértestszám (vérszegénység).
A betegség oka az, hogy a vörösvértesteket kitöltő hemoglobin egyik polipeptidláncában – egy pontmutáció miatt - az egyik aminosav kicserélődik egy másikra. Emiatt a hemoglobin oldékonysága megszűnik, kikristályosodik, merev szálakká alakul, megváltozik a sejt alakja és oxigénszállítása jelentősen csökken. Genetikai eredetű betegség (autoszómális recesszív).
A polipeptidek térszerkezete
A természetben a polipeptidláncnak csak két szabályos stabil formája létezik:
- a béta-redő,
- és az alfa-hélix.
Béta-redő (-szalag)
A béta-szerkezetben a polipeptidláncok – amidcsoportok - összetolt háztetőkhöz hasonló felületet hoznak létre. A szerkezet azáltal stabilizálódik, hogy a láncok párhuzamosan egymás mellé rendeződnek és a peptidkötések – amidcsoportok - között H-kötés alakul ki.
Alfa-hélix
Az alfa-hélixben a polipeptidlánc csavarvonalszerűen tekeredik. A spirál szerkezetét a láncon belül a peptidkötések – amidcsoportok - között kialakuló H-kötések stabilizálják.
A polipeptidlánc térszerkezetét - alfa-hélix vagy béta-redő - a fehérjék másodlagos szerkezetének nevezzük.
A másodlagos szerkezetet alapvetően az aminosavak minősége és sorrendje határozza meg (azaz az elsődleges szerkezet).
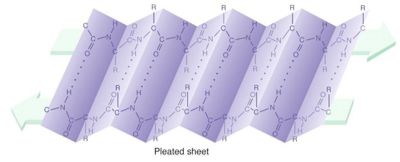 |
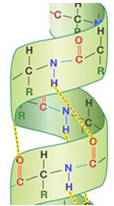 |
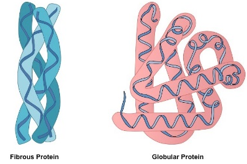 |
Fibrilláris fehérjék
Azokat a fehérjéket, amelyek végig azonos másodlagos szerkezettel jellemezhetők - végig alfa-hélix vagy béta-redő -, fibrilláris fehérjéknek nevezzük.
A fibrilláris fehérjék hosszú, elnyúlt, szálas szerkezetűek, igen stabilak, vízben nem oldódnak.
Általában a szerkezeti fehérjék ilyenek pl.: a keratin, a haj fehérjéje, a kollagén, a miozin, a fibrin stb.
A globuláris fehérjék
A globuláris fehérjékben a polipeptidlánc térszerkezete szakaszonként váltakozik, ezért a molekula egésze gömb alakú. A globuláris fehérjék szerkezetét, az ún. harmadlagos szerkezetet a polipeptidlánc további térbeli elrendeződése határozza meg.
A harmadlagos szerkezet stabilitását a molekulán belül az egyes oldalláncok közötti különféle kötések biztosítják:
- hidrogén-kötés,
- van der Waals-féle kötések,
- ionos kötés,
- kovalens kötés, ilyen a diszulfid-híd.
Az egyes másodlagos szerkezettel rendelkező szakaszok egymáshoz viszonyított térbeli helyzete, tehát a harmadlagos szerkezettel jellemezhető.
A globuláris fehérjék jól oldódnak vízben, kolloid állapotot hozva létre. Ez annak köszönhető, hogy a poláris, hidrofil oldalláncok a gombolyag felületén, míg az apoláris hidrofób oldalláncok a molekula belsejében helyezkednek el. A felszínen levő hidrofil aminosavrészek jól hidratálódnak, az apoláris részek egy hidrofób belső magot hoznak létre. A belső hidrofób mag, ill. a külső hidrátburok nagymértékben hozzájárul a fehérjék stabilitásához.
Ugyanakkor fontos tény, hogy a fehérjék térszerkezete rendkívül bonyolult, ebből következően igen érzékenyen válaszolnak térszerkezetük megváltozásával a környezet hatásaira. A fehérje térszerkezetének jelentősebb megváltozása a denaturáció.
A fehérjék harmadlagos szerkezetét befolyásoló környezeti tényezők:
- a hőmérséklet,
- a közeg hidrogénion koncentrációja (pH),
- a nehézfémsók,
- mechanikai hatások,
- könnyűfémsók.
A hőmérséklet emelésekor a molekularészek hőmozgása egyre intenzívebb lesz, aminek következtében az oldalláncok közötti stabilizáló kötések felszakadnak.
A változás hatására a molekula elveszti jellegzetes térszerkezetét, letekeredik, denaturálódik. A letekeredett láncok összeakadva térhálót alkotnak, melynek hézagaiban vízmolekulák helyezkednek el. A rendszer oldott kolloid állapota megszűnik, a fehérjék kicsapódnak, koagulálnak. A folyamat visszafordíthatatlan, irreverzibilis. (Kis méretű peptidek esetén lehet reverzibilis.)
Irreverzibilis denaturáció zajlik le tojásfőzéskor, hús sütéskor is, minek hatására a molekulák véglegesen elvesztik harmadlagos szerkezetüket, biológiai aktivitásuk megszűnik.
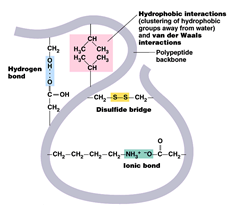 |
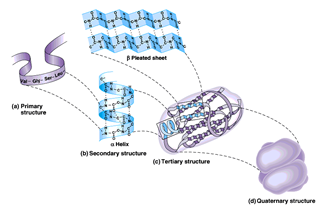 |
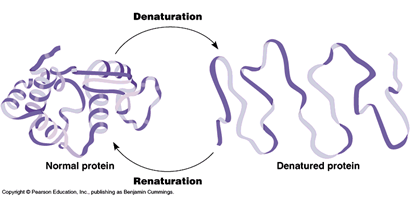 |
Az élő sejtek pH-ja 7,4 körül van, a sejtekben a fehérjék működése, szerkezete ekkor optimális. Amennyiben változik a pH - azaz megváltozik a H+-ion koncentráció -, a bevitt ionok hatására megváltoznak a fehérjemolekulák töltésviszonyai. Az aminosav oldalláncok töltésének megváltozásakor a molekulát stabilizáló kötések felszakadnak, a molekulagombolyag letekeredik, a fehérje irreverzibilisen denaturálódik.
A nehézfémsók - pl. CuSO4, PbCl2, - hatására a fehérjék szintén irreverzibilisen denaturálódnak. A nehézfémionok hozzákapcsolódnak a polipeptidlánchoz, felszakítják a láncot stabilizáló kötéseket.
Ugyancsak denaturálódást eredményezhet a fehérjeoldatok erős mechanikai igénybevétele, pl. ultrahanggal való kezelése, erős rázása.
A könnyűfémsók – pl. NaCl - koncentrációjának emelésekor az oldatba kerülő ionok hidratálódnak és nagy koncentrációjuk esetén saját hidrátburkuk kialakításához a vízmolekulákat a fehérjék hidrátburkából vonják el. A fehérjemolekulák, mivel hidrátburkukat elvesztik, összecsapódnak, azaz koagulálnak (denaturáció nem történik).
Kiválva az oldatból megszűnik a kolloid állapotuk. A folyamat reverzibilis, azaz megfordítható mivel, ha a kicsapódott fehérjékhez feleslegben vizet adunk, a molekulák hidrátburka helyreáll, ismét kolloid állapotba kerülnek.
A folyamatot kisózásnak is nevezzük, amely pl. NaCl vagy (NH4)2SO4 hatására következhet be.
Az alkohol egyrészt elvonja a fehérjék hidrátburkát (dehidratál), másrészt hidrogénkötéseket alakít ki a fehérjékben lévő poláris csoportokkal, ami a molekulaláncok közötti eredeti hidrogénkötések felszakadásához, s így a fehérje denaturálódásához vezet.
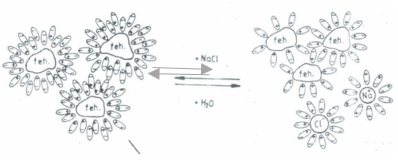 |
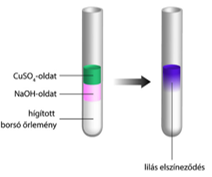 |
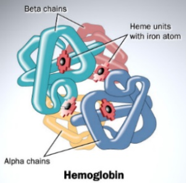 |
A fehérjék kimutatása: biuret-próba
A biuret-próbával a fehérjékben jelenlévő amidcsoportot lehet kimutatni. Pozitív próba esetén ibolyaszíneződést tapasztalunk, mivel a reagensben található rézionok komplexet képeznek az amidcsoporttal.
Egy kevés fehérjeoldathoz adjunk néhány csepp 40%-os nátrium-hidroxid-oldatot, majd 1-2 csepp réz(II)-szulfát reagenst.
https://drive.google.com/file/d/1FeMFJxsNbFNSiFUHMECStgkHkSBipG8G/view?usp=sharing
Negyedleges szerkezet
Ismertek olyan fehérjék, amelyek nem egy, hanem több polipeptidláncból épülnek fel. A fehérjét felépítő egyes polipeptidláncokat alegységnek nevezzük. Az alegységek egymáshoz viszonyított térbeli helyzetét a negyedleges szerkezettel jellemezzük.
Ilyen fehérje pl. a hemoglobin. A hemoglobin négy alegysége egy tetraéder 4 csúcsának megfelelően helyezkedik el, melyeket másodrendű kötések tartanak egyben. Mindegyik alegység egy oxigénmolekulát képes kötni (így a hemoglobinmolekula összesen 4-et).
A fehérjék csoportosítása összetételük szerint is történhet.
- Proteidek vagy összetett fehérjék valamilyen nem fehérjetermészetű, ún. prosztetikus csoportot is tartalmaznak. A prosztetikus csoport erősen kötődik a polipeptidlánchoz, eltávolítása a fehérje térszerkezetének megváltozását eredményezi, ami a biológiai funkció megszűnését jelenti. Ilyen összetett fehérje pl. a hem-et tartalmazó hemoglobin (a hem 4 pirrolgyűrűből felépülő vas-porfirin komplex).
- Proteinek vagy egyszerű fehérjék. Csak aminosavakból állnak:
- albuminok a vérben,
- kollagén,
- inzulin stb.
Stresszfehérjék, dajkafehérjék Forrás: Csermely Péter
A stresszfehérjék legfontosabb szerepe, a hibás, nem megfelelő harmadlagos szerkezettel rendelkező fehérjék helyretekerése. A stresszfehérjék sejtjeink igen nagy mennyiségben jelen lévő, létfontosságú alkotóelemei. A stresszfehérjék segítsége nélkülözhetetlen abban, hogy a hibás szerkezetű fehérjék megtalálhassák a rájuk jellemző, helyes szerkezetet.
A stresszfehérjék szerepe kettős:
- a stresszhatásoknak kitett, károsodott fehérjék szerkezetének helyreállítása (stresszfehérjék),
- a hosszabb polipeptidláncok megfelelő harmadlagos szerkezetének kialakítása (dajkafehérjék).
Ha a sejtet környezeti stressz éri, a sejtben lévő fehérjék károsodnak, kitekerednek. E fehérjéknek az újratekeredéshez éppen úgy a dajkafehérjék segítségre van szükségük, mint születésük pillanatában. Innen a stresszfehérje elnevezés.
Környezeti stressz szinte bármilyen változás lehet:
- lényegesen melegebb, vagy hidegebb;
- a környezet lényegesen savasabb, vagy lúgosabb;
- túl kevés az oxigén, vagy túl sok;
- vízhiány.
A kis méretű fehérjék betekeredése kedvező körülmények között másodpercek alatt végbemegy. A nagyobb fehérjék nehezen találják meg a rájuk jellemző megfelelő harmadlagos szerkezetet, igen gyakran tekeredési csapdák áldozatai lesznek. Ebben az esetben a félkész fehérje egy átmeneti állapotban stabilizálódik, a hidrofób aminosavak a fehérje felszínén maradnak. A hidrofób felszínek az ilyen félkész fehérjéket egymással való összetapadásra, aggregációra teszik hajlamossá. A fehérjeaggregátumok felszaporodása a legtöbb neurodegeneratív betegség, pl. az Alzheimer-kór, a Parkinson-kór, a prionbetegségek egyik fontos oka.
A stresszfehérjéket a többi fehérjét terelgető, szerkezetük kialakításában segítő hatásuk miatt dajkafehérjéknek is nevezik.
Prionok
Amennyiben megváltozik egy fehérjének a térszerkezete, vagy elveszti funkcióját, vagy átalakul egy egészen más szerepű molekulává. Ez utóbbira példa a prionok kialakulása. A prionok olyan fertőző képződmények, melyek nukleinsavat nem, csupán fehérjét tartalmaznak. A prionok a normálisan is meglevő sejtes fehérjék – celluláris prionok – szerkezeti módosulásai. A prionok aminosavsorrendje nem, de a másodlagos szerkezete eltér a normálisan is meglevő sejtes fehérjétől. Felhalmozódva a hidrofób felszínű fehérjék, összecsapódva oldhatatlan rögöket képeznek és az idegsejtek elhalását okozzák. A kóros prionok bontó enzimekkel szemben nem érzékenyek, vízben nem oldódnak (lásd 3.1. fejezet).
A prion betegségre jellemző, hogy nincs ellenanyag-termelés, nincs gyulladásos reakció, a lappangási idő évekig tarthat. Ilyen betegség a juhoknál a surlókór, a szarvasmarháknál a kergemarhakór (BSE), az embernél a Creutzfeldt–Jakob-szindróma és a kuru.
8. tétel: A nukleinsavak biológiai szempontból fontos jellemzői |
A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak nemcsak a sejtmagban vannak, hanem pl. citoplazmában, színtestekben, mitokondriumokban is. A sejtek információtároló és -szállító molekulái.
A nukleinsavakat szerkezet és funkció szempontjából két csoportba oszthatjuk:
- DNS, mely eukariótákban főleg a sejtmagban egyes sejtszervecskékben, mint pl. mitokondriumokban, színtestekben található,
- RNS-ek, melyek főleg a citoplazmában, ill. a fenti sejtszervecskékben fordulnak elő.
A nukleinsavak polimer vegyületek, makromolekulák, s mint ilyenek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
Nukleotidok
A nukleotidok funkciója többrétű:
- a biológiai információtároló és -átadó rendszert alkotó nukleinsavak építőkövei, monomerjei,
- az energiatárolás és -átalakítás központi vegyületei, pl. ATP, GTP,
- enzimek koenzimjeinek alkotórészei, pl. NAD, NADP, KoA, amelyek különböző csoportok szállítását végzik.
A koenzimek az enzimekhez lazán kapcsolódó szállítómolekulák. Ezek felépítésében gyakoriak a vitaminjellegű csoportok, ezért a vitaminhiány gátolja a koenzimek felépülését, ezen keresztül egyes biokémiai folyamatok lejátszódását.
A nukleotidok molekulái savval főzve hidrolizálnak, s a hidrolizátum:
- foszforsavat,
- pentózt, azaz öt szénatomos cukrot,
- N-tartalmú szerves bázisokat tartalmaz.
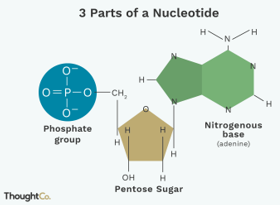 |
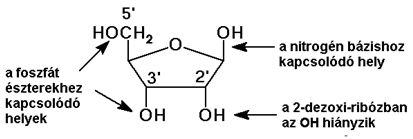 |
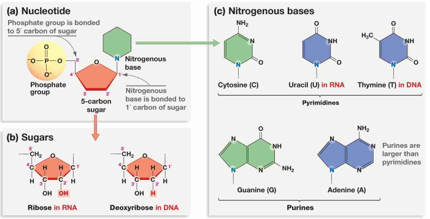 |
A foszforsav
Minden nukleotidban, ill. mind a DNS-ben, mind az RNS-ben megtalálható.
A pentóz lehet:
- ribóz, amely mono- és dinukleotidokban, ill. az RNS-ben található meg,
- dezoxiribóz, amely a DNS-ben fordul elő.
A N-tartalmú szerves bázisok lehetnek:
DNS-ben RNS-ben
A adenin adenin
G guanin guanin
C citozin citozin
T timin U uracil
Tehát az RNS-ben timin helyett uracil van.
Az A, és a G purin-, míg a C, T, U pirimidinvázas vegyületek.
Ezekből a vegyületekből kondenzációval jönnek létre a nukleotidok.
Ha a nukleotidban
- egy foszfátcsoport van: mono-,
- ha kettő: di-,
- ha három: trifoszfátokról beszélünk.
A foszfátcsoportok között nagy energiájú – makroerg - kötés van. Nagy energiájúak azok a kötések, amelyek hidrolízisekor több, mint 25 KJ/mol energia szabadul fel. Ezeknek a kötéseknek az energiatárolásban van kiemelkedő szerepük.
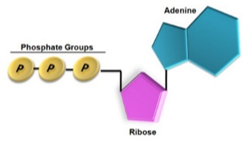 |
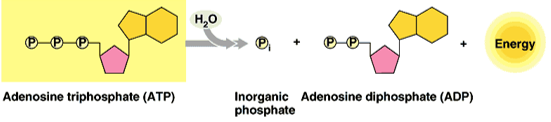 |
Fontosabb mono-, ill. dinukleotidok
ATP (adenozin-trifoszfát)
Az ATP a sejtek legfontosabb energiaraktározó vegyülete. Gamma-foszfátjának hidrolízisekor 30 KJ energia szabadul fel mólonként.
ATP = ADP + Pi Q = 30 KJ/mol
- A lebontó anyagcsere során felszabaduló energia egy része – 40%-a - ATP szintézisére fordítódik és így ATP formájában tárolódik.
- A felépítő folyamatokhoz szükséges energia viszont az ATP hidrolízisekor felszabaduló energiából származik.
Például A és B anyag kondenzációjához energia szükséges. Az ATP bontásakor felszabaduló energia felhasználása közvetett módon történik. Első lépésben a lehasadó foszfátcsoport az összekapcsolódó molekulák egyikére kerül egy nagy energiájú kötéssel, létrehozva ezáltal egy magas energiatartalmú köztes terméket (A~P). Ezt követően energiafelszabadulás közben a köztes termékről lehasad a foszfátcsoport, s a keletkező energia segítségével összekapcsolódik A és B molekula.
A + B + ATP = A~P + ADP + B = A-B + Pi + ADP
Kevésbé jelentősek, de ismertek még GTP, UTP, TTP, CTP energiatároló vegyületek is.
Koenzim – A (KoA)
Szállítómolekula, amely az anyagcserében központi szerepet játszó acetilcsoport - CH3CO - szállítását végzi. A molekulában szereplő pantoténsav B5-vitamin-származék.
NAD+ és NADP+ (nikotinsavamid-adenin-dinukleotid (foszfát))
Az anyagcserében fontos H-szállító vegyületek.
- A NAD+ oxidálószer (H-t vesz fel az átalakuló molekulától) a lebontó (pl. biológiai oxidáció),
- a NADP+ redukálószer (H-t ad át az átalakuló molekulának) a felépítő folyamatokban (pl. fotoszintézis).
A hidrogén szállításakor 2 elektront és 1 protont vesznek fel, a másik proton az oldatban marad.
NAD+ + 2H = NADH + H+
 |
 |
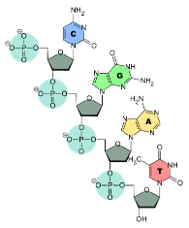 |
Nukleinsavak
A nukleinsavak nukleotid egységekből felépülő polinukleotidok, többnyire makromolekulák, melyekben akár több millió nukleotid is összekapcsolódhat. Az egyes nukleotidokat a foszfátcsoportok kapcsolják össze úgy, hogy hidat képeznek két pentóz 3. és 5. C-atomja között, foszfodiészter-kötést hozva létre.
Tehát a polinukleotid lánc gerincét az egymást követő …foszfát-cukor-foszfát-cukor … váz alkotja, s a bázisok a vázhoz a pentózon keresztül kapcsolódnak.
A nukleotidok kondenzációval kapcsolódnak össze és a polinukleotid lánc hidrolízissel bomlik nukleotidokra.
A polinukleotid lánc elsődleges szerkezete a nukleotidok kapcsolódási sorrendje, melyet szekvenciának nevezünk.
A polinukleotidoknak a felépítésükben résztvevő pentóztól és bázisoktól függően két nagyobb csoportjuk van:
- a DNS-ek és
- az RNS-ek.
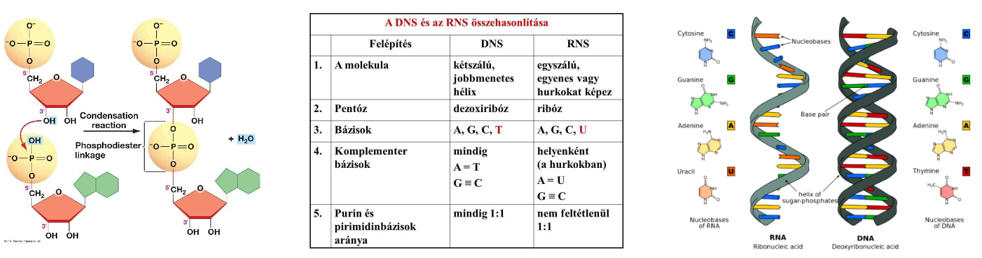 |
A DNS, dezoxiribonukleinsav
Fő tömegük eukariótákban a sejtmagokban koncentrálódik, de előfordulhatnak mitokondriumokban, kloroplasztiszokban, prokariótákban, vírusokban. A DNS egyetlen funkciója a genetikai információ tárolása, ill. továbbadása a szaporodás során.
A polinukleotid lánc felépítésében
- foszforsav,
- dezoxiribóz,
- és négyféle bázis vesz részt: A, T, G, C.
A polinukleotid láncnak két végét különböztetjük meg: 5' és 3' vég. A polinukleotid lánc 5’ végén olyan P-csoport található, amely a pentóz 5. C atomjához kapcsolódik, míg a 3’ végen a cukor 3. C atomjához egy -OH csoport kötődik. A DNS-molekula két egymással szemben levő és ellentétes irányba futó – antiparallel – polinukleotid láncból épül fel. Az antiparallel lefutás azt jelenti, hogy az egyik szál 5’ végével szemben a másik szál 3’ vége található.
A kettős polinukleotid szál a hossztengelye körül spirális formában feltekeredik, s így kialakul a DNS-molekulára jellemző kettős hélix szerkezet. A kettős hélixnek köszönhetően a DNS igen stabil, kémiailag közömbös, nem reakcióképes.
A két láncot hidrogénkötések kapcsolják össze, amelyek a két lánc bázisai között jönnek létre. A H-kötések kialakulását a bázisok szerkezete és térbeli helyzete határozza meg. A bázisok a hélix belseje felé, a hossztengelyre merőlegesen helyezkednek el. Térbeli okok miatt egy nagyobb méretű purin bázissal szemben csak egy kisebb méretű pirimidin bázis helyezkedhet el. A láncok közötti távolság állandó.
Így a következő bázispárok alakulhatnak ki:
- adeninnel szemben mindig timin van,
- guaninnal szemben mindig citozin van.
A G=C bázispárok között 3, az A=T bázispárok között 2 H-kötés létesülhet. Tehát az egyik lánc bázissorrendje egyértelműen meghatározza a másikét is.
Chargaff-szabályok:
- A = T és G = C
- T+C (pirimidinek) = A+G (purinok)
A DNS-molekula lehet:
- nyílt láncú (lineáris), mint pl. az eukariótákban,
- de lehet zárt láncú (cirkuláris), mint pl. a prokariótákban.
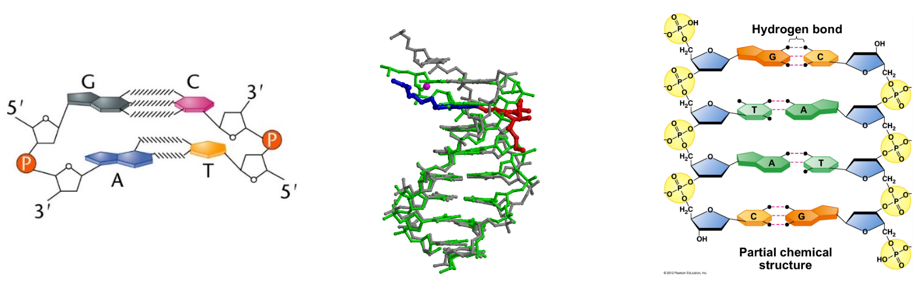 |
RNS-ek, Ribonukleinsavak (részletesen lásd még 6.1. fejezet)
A ribonukleinsavak olyan polinukleotidok, amelyek felépítésében
- foszforsav és
- ribóz mellett
- négyféle bázis vesz részt: A, U, G, C.
A polinukleotid láncra jellemző, hogy mindig egyszálú, legfeljebb a lánc bizonyos helyeken visszahajlik, hurkokat képez, s a molekulán belül jönnek létre hidrogénkötéssel bázispárok (adeninnel szemben U, guaninnal szemben C). Térszerkezetét a nukleotidok sorrendje határozza meg.
A ribonukleinsavaknak a természetben több fajtája ismert, amelyeket funkció szerint csoportosítunk:
Messenger RNS-ek, mRNS -ek
Feladatuk a DNS-ben, a génekben kódolt információ pontos továbbítása a citoplazmába, a fehérjeszintézis helyére. A DNS-ről íródnak át.
Transzfer RNS-ek, tRNS-ek
Feladatuk, hogy a fehérjeszintézis során egy-egy aminosavat szállítanak a fehérjeszintézis helyére, az ún. riboszómákhoz, ahol azok összekapcsolódnak polipeptidlánccá.
Riboszóma RNS-ek, rRNS-ek
A fehérjeszintézis objektumainak, a riboszómáknak a felépítésében vesznek részt.
9. tétel: A lipidek biológiai szempontból fontos jellemzői |
A lipidek olyan szerves vegyületek gyűjtőcsoportja, amelyek igen különböző szerkezetűek, azonban közös sajátságuk, hogy apoláris oldószerekben jól oldódnak.
Fontosabb csoportjaik
- Neutrális zsírok
- Foszfolipidek
- Szteroidok
- Koleszterin
- D-vitamin
- Epesavak
- Hormonok
- Karotinoidok
Neutrális zsírok (trigliceridek)
Ide tartoznak a természetes zsírok és a nem illó olajok. Apolárisak, vízben nem, alkoholban jól oldódnak.
Szobahőmérsékleten azokat a neutrális zsírokat, amelyek
- szilárdak, zsíroknak,
- folyékony halmazállapotúak, olajoknak nevezzük.
A neutrális zsírok kémiailag a glicerin zsírsavakkal alkotott észterei (triacil-gliceridek).
Glicerinből és zsírsavakból kondenzációval jönnek létre, melynek eredményeképpen a kapcsolódó csoportok között észterkötés jön létre.
Bomlásuk hidrolízissel történik, pl. az emésztés során lipáz enzim hatására a vékonybélben.
A zsírsavak lehetnek:
- telítettek, ahol a szénatomok között csak egyszeres kötések találhatók, ilyen a palmitinsav (16) és a sztearinsav (18),
- vagy telítetlenek, melyek molekuláiban a szénatomok között kétszeres kötések is előfordulnak, mint pl. ilyen az olajsav (18).
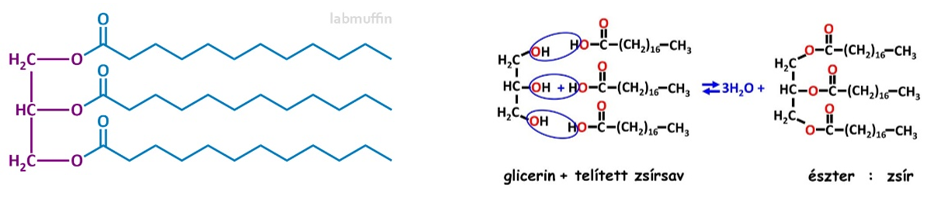 |
A neutrális zsírok biológiai szerepe
Raktározott tápanyagok, kétszer annyi energiát tartalmaznak, mint az azonos tömegű szénhidrátok, mivel
- egyrészt vízmentesen tárolhatók, tekintve, hogy apolárisak,
- másrészt több hidrogén-szén kötést tartalmaznak, azaz redukáltabbak, mint a szénhidrátok, ezért oxidációjukkal több energia nyerhető.
Heterotróf szervezetekben elsődleges energiaraktárak. Állati szervezetekben a zsírszövetekben halmozódnak fel, pl. bőr aljában, szervek körül.
Növényekben pl. olajos magvakban találhatóak meg.
- Helyzetüknél fogva - a bőr alatt - hőszigetelő szerepet látnak el, ill.
- mechanikai szerepük van, pl. szervek helyét rögzítik, talpon puha párnákat képeznek.
- Zsírban oldódó vitaminok (D, E, K, A) oldószerei és raktározó helyei.
Az állati szervezetekben minden feleslegben felvett tápanyag zsírrá alakul.
Foszfatidok (Foszfolipidek)
Alapvegyületük a foszfatidsav, amelynek részei:
- glicerin,
- két zsírsav,
- foszforsav.
A foszfolipidek kettős oldódású, amfipatikus vegyületek, mivel poláris és apoláris részekből állnak, így oldódnak poláris és apoláris oldószerekben egyaránt.
- A glicerin, a foszforsav és a kapcsolódó csoport poláris,
- a zsírsavak apolárisak.
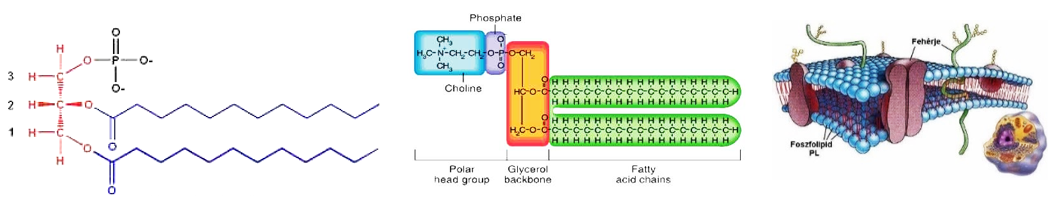 |
Vízben kolloid méretű micellákat, ill. liposzómákat alkotnak, amelyekben a molekulák a vizet maguk közül kiszorítva, apoláris részeikkel egymás felé fordulva, kettős rétegbe rendeződnek.
Legfontosabb biológiai szerepük, hogy a sejtekben határhártyákat, membránokat hoznak létre, így vizes tereket választanak el.
Szteroidok
Az ide tartozó vegyületek közös szerkezeti sajátsága a szteránváz. Az egyes szteroidok a szteránvázhoz kapcsolódó funkciós csoportok minőségében térnek el egymástól.
Koleszterin
Állati zsírokban, vérben, epében, sejthártyában fordul elő. Részben táplálékkal vesszük fel, állati zsiradékok tartalmazzák nagyobb mennyiségben, részben a szervezetünk állítja elő a pl. a májban. Fontos előanyaga a különböző szteránvázas vegyületek – pl. hormonok - szintézisének, ugyanakkor magas koncentrációja a vérben érszűkületet, érelmeszesedést okoz.
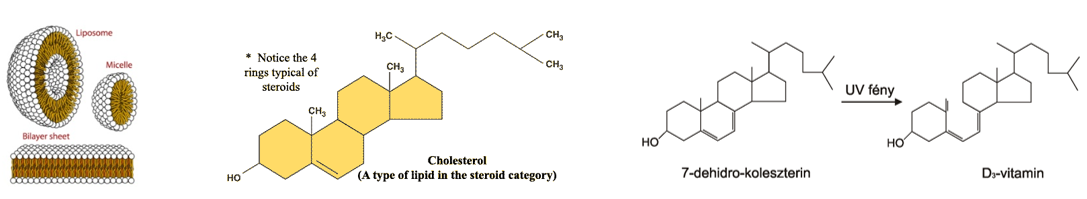 |
D3-vitamin
A szervezetünk optimális kalcium-anyagcseréjéhez szükséges zsírban oldódó vitamin.
E vegyületet
- részben a táplálékkal vesszük fel, magas D-vitamin tartalmú ételek a halmájolajok, tejtermékek, tojás,
- részben a szervezetünk maga állítja elő a bőrben a Nap UV-sugárzásának segítségével.
Képződése röviden:
- koleszterinből előanyag (7-dehidrokoleszterin) képződik a bőrben,
- ebből a szintén a bőrben UV hatására inaktív D3-vitamin (kolekalciferol) lesz,
- ez a májban raktározódik és tovább módosul (25-hidroxi-kolekalciferollá),
- majd a vesében aktív vitaminná alakul (1,25-dihidroxikolekalciferollá).
Szerepe
A D3-vitamin egy kalciumtranszport-fehérje képződését segíti elő a vékonybél nyálkahártyának hámjában génátírás aktiválása révén.
Ezért:
- fokozza a kalciumionok felszívódását a bélből,
- ezáltal emeli a vér kalcium koncentrációját,
- elősegíti a kalcium beépülését a csontokba.
Továbbá fokozza az immunrendszer működését. (Télvégi influenzajárványok egyik oka a szervezet alacsony D-vitamin tartalma.)
Hiánya
A D-vitaminhiány
- kalciumhiányt eredményez,
- ami miatt csonttömegvesztés jön létre,
- fiatalkorban az angolkór kialakulásához vezet, jellegzetes csontrendszeri elváltozásokat (a gerinc és a mellkas torzulásai) okoz.
Nem csak a vitaminok hiánya, hanem a zsírban oldódó vitaminok esetén azok túladagolása (hipervitaminózis) is egészségügyi kockázatokkal jár, mivel zsírokban oldódva nem ürülnek ki, felhalmozódnak a zsírszövetekben. Pl. a D-hipervitaminózis magas vérkalciumszinttel jár, ami erek és szövetek elmeszesedéséhez, vesekő képződéséhez vezethet.
A folyamatosan adagolt mesterséges vitaminkészítmények esetén, mindig ügyelni kell a kontrollált és optimális vitaminmennyiségre.
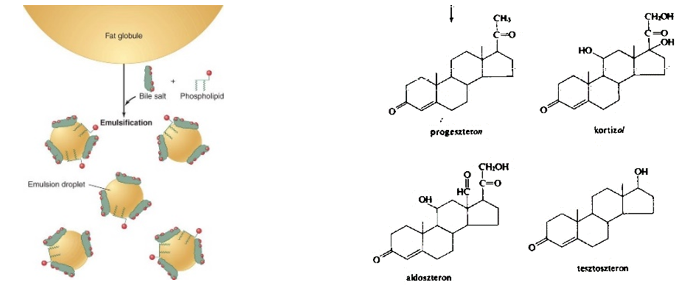 |
Epesavak olyan szteroidok, amelyek az epében találhatók meg, sók formájában fordulnak elő, ezért az epe nem savas, hanem lúgos.
A középbélben segítik – nem végzik - a zsírok emésztését,
- egyrészt aktiválják a zsírbontó lipázokat,
- másfelől a bélmozgások miatt felaprózódott zsírcseppek felületén elhelyezkedve csökkentik az emulgeált zsírgolyócskák felületi feszültségét, stabilizálják azokat, és fajlagos felületüket nagyobbítva kedvezőbb feltételeket teremtenek a lipáz emésztés számára (hatásuk a szappanokéhoz hasonló).
Az epesavak amfipatikus vegyületek, apoláris részükkel (szteránváz) körbeveszik a zsírcseppeket, poláris részükkel (OH csoportok) pedig a vizes fázis felé fordulnak. Hatásuk a szappanokéhoz hasonlítható.
Szteránvázas hormonok Részletesen lásd hormonális rendszer
Mellékvesekéreg-hormonok:
- kortizol: szénhidrát-anyagcserére hat.
- Aldoszteron: a só visszaszívását serkenti a vesében.
A petefészekben termelődő női nemi hormonok:
- progeszteron: megőrzi a terhességet.
- Ösztrogén: egyik fő feladata a női másodlagos nemi jellegek kialakítása.
A herében termelődő férfi nemi hormon:
- tesztoszteron: férfi másodlagos nemi jellegek kialakításában játszik szerepet.
Karotinoidok
A karotinoidok többnyire szénhidrogének. Konjugált kettőskötés-rendszerűek, ahol az egyszeres és a kétszeres C-C kötések szabályosan váltakoznak. Az ilyen elektronrendszer delokalizált, könnyen gerjeszthető, ezért a karotinoidok
A karotinoidok, elektronrendszerük könnyű gerjeszthetősége révén, különféle fotokémiai reakciókban jelentősek, mint pl. fotoszintézis, látás.
A karotin a sárgarépa színanyaga, tartaléktápanyaga, az A-vitamin előanyaga, fotoszintetikus pigmentként a fotoszintézis fényszakaszában a fényenergia elnyelésében játszik szerepet.
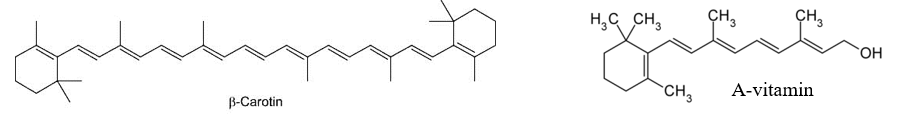 |
Az A-vitamin (retinol) a májban karotinból keletkező, majd raktározódó zsírban oldódó vitamin.
Szerepe
- A retina fényérzékenységét biztosító rodopszin (fényérzékeny anyag) egyik összetevőjének, a retinalnak előanyaga, ezért a normális látás fenntartásában nélkülözhetetlen,
- fontos a hámfelületek védelmében, ezért véd a hámon keresztül történő fertőzések ellen.
Fő forrása
- Vitamin formájában a halmájolajok, tejtermékek, máj tartalmazzák. Előanyagát a karotint, tartalmazza a legtöbb zöldség.
Mivel vízben nem oldódik, a felszívódásához zsiradék szükséges.
Hiánya
- Szürkületi vakságot (farkasvakságot), sőt teljes vakságot okozhat.
- Hámszövetek, mirigyek károsodnak, a bőr kiszáradása, a szőrzet és a hajszálak törékenysége, kihullása tapasztalható.
K1,2 –vitamin
Szerepe
- A normális véralvadáshoz kell, jelenlétében a máj egy véralvadási faktort, protrombint szintetizál.
Fő forrása
- K1-t növényi olajok, zöld leveles zöldségek, káposzta, brokkoli, paraj tartalmazzák.
- A K2-t bélbaktériumok termelik. Éppen ezért hosszan tartó, bélgyulladással járó betegség, vagy antibiotikumos terápia következményeként K-vitaminhiány állhat elő.
Hiánya
- Vérzékenységhez, véralvadási zavarokhoz vezet.
Retinál
A szem fényérzékeny anyaga, az A-vitaminból keletkezik. A látás lényege, hogy fény hatására a cisz-retinál transz-retinállá izomerizálódik (fotokémiai reakció).
10. A sejtanyagcsere és a mitokondrium |
Eukarióta sejtekben a lebontó folyamatok – biológiai oxidáció - nagy része külön sejtszervecskékben, a mitokondriumokban zajlik.
A mitokondriumokban folyik
- a citrát-ciklus,
- a terminális oxidáció.
Prokarióta sejtekben a mitokondrium funkcióit – pl. terminális oxidáció - a sejthártya betüremkedései során létrejövő membránrendszer végzi.
A mitokondriumok minden eukarióta – növényi, állati, gomba - sejttípus alapvető sejtszervecskéi. Mivel a disszimiláció során a terminális oxidációban keletkezik a legtöbb ATP, ezért a mitokondriumokat a sejtek erőműveinek tekinthetjük.
A mitokondriumok mérete, alakja, egy-egy sejtben számuk rendkívül változatos.
- Méret: baktérium méretűek, hosszuk 5-1O mikrométer.
- Alak: fonalszerű (mítosz), ill. gömbszerű (kondrion), lehet ovális.
- Számuk: egysejtűekben 1-2, emlősök májsejtjében több ezer, kétéltűek petéjében akár 3OO OOO. Mennyiségük arányos a sejtek lebontó anyagcseréjének intenzitásával.
Felépítés
Kettős membránrendszerből épülnek fel. A citoplazmától a külső határoló membrán választja el, amely alatt található a változatos felépítésű belső membrán. (Mintha egy kisebb zsákba begyűrnénk egy nagyobb zsákot.) A két membrán között rés található, amelyet külső kamrának nevezünk. A belső membrán által határolt teret belső kamrának nevezzük, az azt kitöltő anyagot pedig mátrixnak hívjuk, ez a mitokondrium alapállománya.
A mátrixban találhatók
- a citrát-ciklus enzimjei,
- a mitokondriális DNS, RNS-ek,
- riboszómák.
A mitokondrium két membránja igen eltérő felépítésű és működésű.
A belső membrán belső felületén apró gömbök – ATP-szintázok - találhatók, melyek vékony nyéllel kapcsolódnak a belső membránhoz. Ezek a részecskék az ATP-képződés helyei.
Ugyanitt – a belső membránban - található a terminális oxidáció teljes enzimkészlete és az elektrontranszportlánc. Az enzimek, a citokrómok membránban való elhelyezkedése biztosítja, hogy az egyes lánctagok megfelelő sorrendben kövessék egymást. Ez lehetővé teszi a folyamatok összerendezését, azok egyirányúságát.
A külső membrán feladata a mitokondrium belső állományának az elhatárolása és összekötése a citoplazmával. Ennek megfelelően elsősorban transzportfolyamatokat lát el, így membránjában igen sok szállítófehérje van.
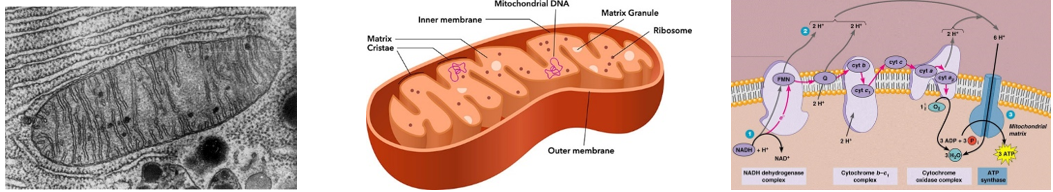 |
A mitokondrium működése
A citoplazmában folyik a szénhidrátlebontás glikolízis szakasza, a glükóztól kiindulva a piroszőlősav (PS) keletkezéséig.
Anaerob körülmények esetén a PS a citoplazmában marad és erjedés útján alakul tovább (pl. emlősök vázizomrostjaiban tejsavvá).
- Aerob körülmények között a PS a citoplazmából bejut a mitokondrium belső terébe, s ott egy enzim segítségével acetil-csoporttá alakul.
- Az acetil-csoport bekerül a mátrixban zajló citrát-ciklusba.
- A felszabaduló CO2 eltávozik, és a keletkező H-ek NADH közvetítésével eljutnak a belső membránban található elektronszállító-rendszerhez, amelynek elején a protonok és az elektronok szállítása különválik.
- Az elektronok áramlásának eredményeképpen a felszabaduló energia protonok szállítására fordítódik a mátrixból a külső kamrába.
- A protontranszport eredményeképpen a külső kamrában jelentősen megnő a protonkoncentráció, így a membrán két oldalán kialakul egy protonkoncentráció-gradiens.
- Amikor a protonkoncentráció-kiegyenlítődés megindul, a belső membránban lévő ATP-szintázok segítségével, a felszabaduló energia ATP szintézisére fordítódik.
- Az elektronokat végül a légköri oxigén veszi fel, amely a beáramló protonokkal vízzé egyesül.
Az ATP-képződés mechanizmusát a Mitchell-féle kemiozmotikus elmélet írja le, melynek lényege, hogy az elektronok áramlása közben felszabaduló energia segítségével az elektron transzportlánc fehérjéi protonokat pumpálnak az alapállomány felől a külső kamrába. A folyamat többszöri lejátszódása miatt jelentős protonkoncentrációbeli különbség alakul ki a belső membrán két oldala között. A különbség kiegyenlítését az alapállomány felé néző enzimkomplex, az ATP-szintáz végzi. Az enzimkomplexen átáramló protonok energiája ATP-szintézisre fordítódik.
A mitokondriumban keletkezett ATP kijut a mitokondriumból és a sejt egyéb energiaigényes folyamataihoz felhasználódik (pl. felépítő folyamatok, aktív transzport).
Összefoglalva a citoplazma és a mitokondrium kapcsolatát:
Citoplazmából a mitokondriumba:
- piroszőlősav,
- NADH,
- elemi oxigén, O2,
- ADP + P.
A mitokondriumból a sejtplazmába:
- ATP,
- NAD+,
- víz,
- CO2.
A petesejtben több százezer mitokondrium van, a spermiumban azonban csak alig néhány száz. A megtermékenyítés után a zigótában egyedül az anyától, a petesejtből származó mitokondriumok találhatók meg, mivel a spermiumok petesejtbe bejutó feji része nem tartalmaz mitokondriumokat. A mitokondriális géneket tehát kizárólag az anyától kapja az utód, az ezek által meghatározott tulajdonságok csak anyai úton öröklődnek.
A mitokondriumok DNS-üknek, riboszómáiknak köszönhetően képesek megkettőződni.
Számos betegség a mitokondriális DNS mutációi következtében alakul ki. Mivel a mitokondrium fő funkciója az energiatermelés, ezért a mitokondriális betegségek a legnagyobb energiaigényű szerveket érintik leginkább, mint pl. a vázizomzatot, ill. az idegrendszert.
A biológiai oxidáció
Ha a glükóz lebomlása oxigén jelenlétében folyik, aerob körülmények között, biológiai oxidációról beszélünk. A glükóz lebomlásának leghatékonyabb módja, melynek terméke a szén-dioxid és a víz.
C6H12O6 + 6 O2 = 6 CO2 + 6 H2O
Három fő szakaszra osztható:
- a glikolízis,
- a citrátkör,
- a terminális oxidáció.
A glikolízis
Glikolízisnek nevezzük a glükóztól a piroszőlősavig vezető reakciósorozatot.
- A glükózlebomlás első szakasza.
- Nem kell hozzá oxigén, ezért
- az erjedési folyamatok részét is képezi.
- A sejtek citoplazmájában játszódik le.
A glikolízis eredményeképpen a glükózból molekulánként keletkezik:
- 2 molekula piroszőlősav (piruvát),
- nettó 2 molekula ATP,
- 2 molekula NADH + H+ (H-leadás miatt oxigén nélküli oxidáció történik!).
A glikolízis nettó energianyeresége 2 ATP glükózmolekulánként (szubsztrát szintű foszforiláció).
A piroszőlősavnál (piruvát) a reakcióutak szétágaznak.
- Amennyiben a lebomlás anaerob körülmények között zajlik, a piroszőlősav az erjedés folyamatában alakul tovább (lásd később).
- Azonban, ha a disszimiláció oxigén mellett folyik, a reakcióút a citromsavciklusba
Aerob körülmények között a piroszőlősav CO2 és H vesztése mellet acetilcsoporttá alakul.
Az acetilcsoport szállítómolekulához, KoA-hoz kapcsolódik, amely elszállítja a citrátkörbe.
Piroszőlősav + KoA = acetil-KoA + CO2 + NADH + H+ (oxidatív dekarboxileződés)
A piroszőlősav aerob bomlása és a további reakciók eukariótákban már a mitokondriumban, prokariótákban a citoplazmában játszódnak le.
A citrátkör, citromsavciklus, Szent-Györgyi-Krebs-ciklus
A citrátkör lényege, hogy az acetilcsoport C-atomjai szén-dioxiddá alakulnak. A keletkezett szén-dioxid a légzés útján távozik szervezetből.
Az oxidáció során felszabaduló hidrogének H-szállító koenzimekhez (NAD+, FAD) kapcsolódnak.
A ciklus során több lépésben távoznak a szén-dioxid- molekulák, ill. a H-atomok.
Fő lépések röviden
Az acetilcsoportot egy négy szénatomos karbonsav az oxálecetsav veszi fel, amely ily módon egy 6 szénatomos vegyületté, citromsavvá alakul.
A citrát több lépésen keresztül lebomlik.
Közben:
- 2 molekula CO₂ távozik,
- 3 NADH, 1 FADH₂, és 1 GTP képződik.
A ciklus végén újra oxálacetát keletkezik.
A folyamat energianyeresége minimális, mindössze egyetlen GTP keletkezik acetilcsoportonként, melynek bomlása ATP képződését teszi lehetővé.
A citrátkör az anyagcsere egyik legfontosabb anyagelosztója. A folyamat köztestermékei különböző bioszintézisek kiindulási vegyületei.
Az eddigi folyamatok során a glükóz C-atomjai szén-dioxiddá alakultak:
- piroszőlősav acetilcsoport átalakuláskor (2 CO2),
- a citrátciklus során (4 CO2).
A glükóz hidrogénjei H-szállító koenzimekhez kapcsolódtak, NADH-kat képezve, amelyek nagy energiatartalmú vegyületek, mivel erősen redukáltak. Energiatartalmuk felszabadítása végső oxidációjukkal történik, amely a terminális oxidáció folyamatában valósul meg.
 |
A terminális oxidáció
A terminális oxidáció lényege, hogy a NADH-ról származó hidrogének elektronjai - a fotoszintézis fényszakaszához hasonlóan - egy elektrontranszportláncba kerülnek, ahol az elektronok áramlása energiafelszabadulással jár, amely ATP szintézisére fordítódik (oxidatív foszforiláció). Az elektronok szállításában vastartalmú összetett fehérjék, ún. citokrómok vesznek részt.
Az elektrontranszportlánc utolsó tagja oxigént köt meg, így az elektronok végső soron az oxigénre kerülnek. Az így keletkezett oxidion az oldatban található protonokkal vízzé egyesül.
Tehát a NADH szubsztrátoktól átvett protonjai és elektronjai – hidrogénjei - egy bonyolult elektron-transzportláncon keresztül jutnak el a végső elektronfelfogó molekulához, az oxigénhez.
A teljes folyamatsort, a glükóztól a szén-dioxid és a víz keletkezéséig, sejtlégzésnek, biológiai oxidációnak nevezzük.
A biológiai oxidáció során felszabaduló energia közel 95%-a a terminális oxidáció során szabadul fel, ami 34 mol ATP-t jelent 1 mol glükóz esetén.
Ezen kívül a glikolízisben 2, a citrátciklusban is 2 ATP (GTP) jön létre glükózonként.
Összesen 38 (újabban 32) ATP keletkezik glükózonként, ami átszámítva grammonként 17,2 kJ energiát jelent mólonként. A glükóz biológiai oxidációjának hatásfoka 40%, azaz a glükóz energiatartalmának csupán 40%-a épül be ATP-be, a többi hővé alakul.
11. tétel: Zöld színtest és a fotoszintézis |
Plasztiszok-színtestek
A színtestek vagy plasztiszok a növényi sejtekre jellemző, kettős membránnal határolt sejtszervecskék. A citoplazmában helyezkednek el, kiemelkedő jelentőségűek a sejtek felépítő anyagcsere folyamataiban. Egysejtű és többsejtű növények sejtjeire jellemzők, prokariótákban, állatokban, gombákban nem találhatók meg.
A színtestekben lévő színanyagok ill. működésük alapján több típusukat különböztetjük meg, legfontosabbak a többnyire zöld színű, klorofillokat tartalmazó, fotoszintetizáló kloroplasztiszok-zöld színtestek.
A kloroplasztiszok-zöld színtestek
A legjellemzőbb növényi sejtszervecskék, amelyek a fotoszintézis lebonyolításában vesznek részt, így jelentőségük az egész földi élet szempontjából rendkívüli. Jellemzően előfordulnak a táplálékkészítő alapszövet sejtjeiben, továbbá a bőrszöveti zárósejtekben, ugyanakkor fénytől elzárt növényi részekben – pl. gyökerekben – nem találhatók meg.
Felépítés
Minden kloroplasztiszt kettős membrán határol el a citoplazmától. Belsejében a finoman szemcsés alapállományt sztrómának nevezzük. A sztrómában zajlik a fotoszintézis sötét szakasza, emiatt sok keményítőszemcsét tartalmaz.
Ezenkívül található még itt DNS, RNS, riboszómák, amely arra utal, hogy a plasztiszoknak bizonyos fokig önálló genetikai és fehérjeszintetizáló apparátusuk van. A DNS-tartalommal hozzák összefüggésbe a kloroplasztiszok osztódóképességét.
A sztrómában belső membránrendszer van, amelyet tilakoid-membránnak nevezünk. A belső membránrendszer felépítése alapján megkülönböztetünk lemezes, ill. gránumos kloroplasztiszokat. A lemezes kloroplasztiszok eukarióta moszatokban fordulnak elő.
A gránumos kloroplasztiszok mohákban, harasztokban, valamint nyitva- és zárvatermő magvas növényekben találhatók meg.
- Számuk változatos, egy-egy sejtben akár több száz is lehet.
- Alakjuk főleg lencse vagy gömb.
- Méretük 5-8 mikrométer.
A belső membránrendszer
A tilakoid membrán korongokat képez, melyek oszlopokká – gránumokká szerveződnek. A gránumok alakja az egymásra helyezett korongokból álló oszlopra hasonlít leginkább. Az egyes gránumok nem függetlenek egymástól, hanem összeköttetésben állnak. Megfigyelhető, hogy egy-egy membránlemez áthúzódik egy vagy több gránumon.
 |
A tilakoid membránok felépítése, működése
- A kloroplasztiszt körülvevő kettős határolómembrán nem vesz részt a fotoszintézisben, hanem működése elsősorban szállítófolyamatokkal kapcsolatos.
- A gránumok membránjában zajlik a fotoszintézis fényszakasza.
- A fényenergia megkötését a tilakoid membránban lévő pigmentrendszerek végzik.
- A fotorendszereket az elektronszállító-rendszer köti össze.
- A fotolízis a gránumok belső terében játszódik le, a keletkező elektronok az elektronszállító-rendszerbe kerülnek, a felszabaduló protonok pedig a membránzsákok belső terében halmozódnak fel.
- Az elektronok áramlásakor felszabaduló energia közvetlenül arra fordítódik, hogy a gránumok membránjában található protonpumpák protonokat szállítanak a sztrómából a gránumok belsejébe.
- Ennek köszönhetően a gránummembrán két oldala között jelentős proton koncentrációkülönbség alakul ki.
- A protonok kiáramlását a sztrómába speciális enzimek, ún. ATP-szintázok teszik lehetővé.
- A protonok koncentrációjának kiegyenlítődése energiafelszabadulással jár, amely ATP szintézisére fordítódik.
- A sztrómába kijutott protonok az elektronokkal egyesülve H-atomként a NADP-hez kapcsolódnak, amely így NADPH-vá alakul.
A citoplazma és a zöld színtest kapcsolata:
Citoplazmából a színtestbe:
- CO2,
- H2O.
A színtestből a citoplazmába:
- O2,
- glükóz.
Tehát a színtestben keletkezett ATP kizárólag a színtest energiaigényes folyamataihoz használódik fel, pl. a sötét szakaszhoz, amely a sztrómában játszódik le.
A fotoszintézis
Lényege
Azon folyamatok összessége, amelynek során a fényenergia kémiai energiává alakul át, azaz fényenergia segítségével nagy energiájú vegyületek - ATP, NADPH – jönnek létre. Az így átalakított energia felhasználásával szerves anyag – glükóz - előállítása történik.
Jelentősége
- Az egész élővilág energiaigényes folyamataihoz az energiát végső soron a fotoszintézis során átalakított fényenergia szolgáltatja, vagyis a földi élet végső energiaforrása a Nap sugárzó energiája. A bioszféra összes szerves anyaga – a jelentéktelen bakteriális kemoszintézist leszámítva – a fotoszintézisben keletkezik a tápláléklánc termelői szintjén, ez halad végig a táplálékláncon és építi fel az élőlények testét, ill. ennek a szerves anyagnak a bontása szolgál minden energiaigényes folyamat energiaforrásaként.
- A Föld légkörének teljes oxigéntartalma fotoszintetikus eredetű.
A fotoszintézis előfordulása
Fotoszintézisre mind prokarióta, mind eukarióta szervezetek képesek, így baktériumok (pl. kékbaktériumok), egysejtű algák, növények.
A fotoszintézis általános egyszerűsített egyenlete
6H2O + 6CO2 = C6H12O6 + 6O2
A tényleges arányokat kifejező bruttó egyenlet: (6 CO2 + 12 H2O = C6H12O6 + 6 H2O + 6 O2)
A CO2 redukciója továbbá nemcsak glükózt, ill. keményítőt ad, hanem minden más molekula szénváza végső soron a fotoszintézisben megkötődött szén-dioxidból származik.
A fotoszintézis folyamatát két fő szakaszra bonthatjuk:
- Fényszakasz - csak fény jelenlétében játszódik le.
Ebben a szakaszban történik a CO2 redukciójához szükséges fényenergia elnyelése, átalakítása kémiai energiává, az ATP- és a NADPH-molekulák szintézise. A növények az őket ért fényenergia kb. 1%-át kötik meg és hasznosítják.
- Sötét szakasz – közvetlenül nem szükséges hozzá fény, azonban lejátszódása előfeltételezi a fényszakasz működését. Itt történik a CO2 megkötése és redukálása, a glükóz előállítása az előző folyamatban átalakított fényenergia – ATP és NADPH - segítségével.
A fényszakasz - a fényenergia megkötése és átalakítása kémiai energiává
A fényszakaszban
- a fényenergia felhasználásával vízbontás történik, melynek során a víz hidrogénre és oxigénre bomlik (a folyamat mellékterméke az oxigén, melyet minden aerob szervezet a biológiai oxidációban hasznosít),
- a vízből nyert hidrogén elektronja egy elektrontranszportláncban szállítódik, majd a szintén vízből származó proton felhasználásával NADP redukálására, vagyis NADPH előállítására használódik fel.
- A nagy energiájú elektronok áramlásakor felszabaduló energia ATP szintézisére fordítódik.
A fényenergia megkötését speciális elektronszerkezetű színes molekulák, az ún. fotoszintetikus pigmentek végzik: ilyenek
- a klorofillok,
- karotinok és
- xantofillok.
A karotinoidok nem csupán a fényenergia megkötését végzik, hanem védik a klorofillokat az oxigén jelenlétében bekövetkező károsodástól, fotooxidációtól, ami erős fényben jöhet létre.
Ezekben a molekulákban egyaránt megtalálható a delokalizált konjugált kettőskötés-rendszer, amely felelős a fényenergia megkötéséért.
A klorofillok
A magasabb rendű növények kétféle klorofillt tartalmaznak:
- kékeszöld kl-a-t és
- sárgászöld kl-b-t.
Mindkettő Mg-porfirin-komplex. A gyűrű közepén egy Mg2+-ion található.
A földfelszínt elérő napsugárzás zöme a 4OO-8OO nm hullámhosszúságú tartományba esik. A fényenergia megkötése úgy történik, hogy a fényt elnyelő pigment delokalizált elektronrendszere gerjesztődik, azaz az elektronok távolabb kerülve az atommagtól magasabb energiájú pályára lépnek. Megfelelően nagy mennyiségű energia elnyelésekor egy elektron leszakadhat a molekuláról.
Minden pigmentre egyformán jellemző a fényelnyelő képesség, de közülük csak néhány képes arra, hogy leadjon elektront. Az elektronleadás csak az ún. reakciócentrumokban megy végbe.
A többi pigment esetén a gerjesztési energia átadódik a szomszédos molekulának, ezek a pigmentek mintegy az antenna szerepét töltik be. Az elektronjaik gerjesztődnek, majd visszatérnek alapállapotba és a felvett energiát a reakciócentrumban levő, egy fehérjéhez kapcsolt speciális klorofill-a molekulának sugározzák, ahol az nagy mennyiségű energiát összegyűjtve lead egy elektront.
 |
Pigmentrendszerek - fotorendszerek
A különféle típusú pigmentek az együttműködés érdekében ún. pigmentrendszerekbe tömörülnek. A pigmentrendszerek - fotorendszerek - közepén található a reakciócentrum. A reakciócentrum körül helyezkednek el az antennapigmentek.
Az elektronszállító rendszer
A fotorendszerekbe begyűjtött fényenergia segítségével a leadott elektronok egy elektronszállító rendszerbe kerülnek.
Az elektronszállító rendszert többek között ún. citokrómok építik fel. A citokrómok olyan összetett fehérjék, amelyek nem fehérje része Fe-porfirin komplex.
A citokróm-molekulák szorosan egymás mellett elhelyezkedve adják át egymásnak az elektronokat. A transzportlánc egy tagja az előtte elhelyezkedő molekulától felveszi az elektront, a porfirinváz közepén levő Fe3+-ion Fe2+-ionná redukálódik, majd továbbadva a következő tagnak visszaoxidálódik Fe3+-ionná.
Az elektronszállító rendszerben az áramló elektronok végső soron a vízből származnak. A víz bontása a rendszer elején folyik:
H2O = 2H + 1/2 O2
2H = 2H+ + 2e-
A keletkező elektronok bekerülnek az elektronszállító rendszerbe. Mivel a víz bontása kapcsolatban van a 2. fotorendszerrel és fényenergia szükséges hozzá, ezért a folyamatot fotolízisnek nevezzük. A folyamat mellékterméke az oxigén.
Az elektronszállító rendszer végén az elektronok és a H+-ok a NADP+-ra kerülnek, amit NADPH-vá redukálnak.
NADP+ + 2H = NADPH + H+
A víz rendkívül stabil vegyület, ezért bontása, ill. a belőle származó elektronokkal a NADP+ redukciója nagy energiát igényel.
Tehát a fényszakaszban a víz felől a II.-es, majd I.- es fotorendszer közvetítésével folyamatos elektronáramlás zajlik a NADP+ felé a fényenergia segítségével.
A két fotorendszer között az elektron áramlása energiafelszabadulással jár, amely ATP szintézisére fordítódik.
A fényszakasz működésének eredményeként a fényenergia megkötődik és nagy energiájú vegyületekben tárolódik, NADPH-ban és ATP-ban.
Összefoglalva
- A fényszakaszban történik a glükóz előállításához szükséges fényenergia megkötése.
- A fényenergia megkötését különféle pigmentek végzik, az elnyelt energiát elektronok leadására fordítják.
- Az elektronok pótlása végső sorban a vízbontásból történik, mely folyamat mellékterméke az oxigén.
- A vízből származó hidrogének végső soron NADP+-re kerülnek.
- A NADP redukciója energiaigényes, ezért az elektronok egy elektronszállító rendszeren – melybe beiktatva helyezkedik el a két fotorendszer, biztosítva az energiaigényt – keresztül jutnak el az elektronfelvevő molekuláig, a NADP-ig.
- Az elektronok áramlása során energia szabadul fel, amely ATP formájában konzerválódik.
- A fényszakasz működésének eredményeként a fényenergia megkötődik és nagy energiájú vegyületekben tárolódik, NADPH-ban és ATP-ban.
A fotoszintézis sötét szakasza (Calvin-ciklus, redukciós ciklus, C3-as út)
A fotoszintézis sötét szakasza a
- CO2 megkötését,
- redukálását,
- a glükóz bioszintézisét jelenti.
A fotoszintézis fényszakaszában történik a fényenergia megkötése, ill. konzerválása ATP és redukált NADPH formájában.
A szénhidrátok szintézise a CO2 redukciója útján ezen energia felhasználásával történik.
A glükóz elsőként a zöld színtestben keményítő formájában raktározódik, majd éjszaka szacharóz formájában a háncsrészben elszállítódik a növény többi részébe. Az elkészült glükóz kiindulási vegyülete lesz a növényi szervezetet felépítő többi vegyületnek, pl. keményítőnek, lipideknek, fehérjéknek, nukleinsavaknak.
A CO2-fixáció eme útját felfedezőjéről – Malvin Calvin – Calvin-ciklusnak (C3-as út) nevezzük (1961 Nobel-díj). Fő folyamatai:
- karboxiláció – a CO2 megkötése,
- redukció – a CO2 redukciója, glükóz bioszintézise,
- regeneráció – a RuDP újraképződése.
Animáció
https://www.youtube.com/watch?v=gnNDC1aOBU0
12. tétel: Anyagcsere típusok |
A biológiai rendszerek anyagi rendszerek. Az anyagi rendszereket általában anyag- és energiaáramlás jellemzi és működésük fenntartásához sokszor energiára van szükség. A Föld globális rendszerének működéséhez a meghatározó külső energiaforrás a Nap sugárzó energiája.
Az anyagi rendszerek csoportosíthatók aszerint, hogy az adott rendszer és környezete között van-e anyag- és energiaforgalom. Eszerint:
- Az izolált rendszerek sem anyag-, sem energiaforgalomban nem állnak a környezetükkel.
- Zárt anyagi rendszer esetén energiaforgalom lehetséges, de anyagáramlás nincs a rendszer és környezete között.
- Nyílt anyagi rendszer olyan rendszer, amely a környezetével anyag- és energiaforgalmat bonyolít le. Ilyenek az élő rendszerek.
Az élő rendszerek működése szempontjából az anyag- és energiaáramlás mellett elengedhetetlen az információáramlás. A háromféle folyamat összekapcsolódik, egymástól szét nem választható, az energia és az információ anyagok formájában áramlik.
Az élő rendszerek, mint nyílt anyagi rendszerek a környezetükből anyagot és energiát vesznek fel, ill. adnak le.
Az élő rendszerek energiaforgalma
A sejtek – és általában az élő szervezetek – működésükhöz, életfolyamataik fenntartásához folyamatosan energiát igényelnek.
Az élő szervezetekben a legjelentősebb a kémiai energia. A kémiai energia kémiai kötések hasadásához szükséges, illetve kialakulásukkor felszabaduló energiamennyiség.
A biológiai energia a kémiai energia egy speciális fajtája, amely a sejtekben nagy energiájú kémiai kötésekkel kapcsolatos. A nagy energiájú kötések energiát tároló vegyületekben találhatók meg. Ilyen minden élő szervezetben megtalálható energiatároló és -szállító vegyület az ATP.
- Az élő szervezetekben minden energiaigényes folyamat – mozgás, anyagszállítás, -felvétel, -leadás, nagy molekulájú szerves vegyületek előállítása - közvetlen energiaforrása az ATP nagy energiájú kötésének bomlásakor felszabaduló energia.
ATP = ADP + Pi + energia
- Másfelől pedig a szerves vegyületek lebontása során felszabaduló energiát, vagy a fotoszintézis során megkötött fényenergiát a sejtek ATP előállításával képesek eltárolni.
ADP + Pi + energia = ATP
A sejtek ATP-készlete állandó, energiaigényes folyamatokban energiát szolgáltatva bomlik, energiatermelő folyamatokban pedig a felszabaduló energiát megkötve regenerálódik.
Az intermedier anyagcsere
Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az anyagcsere során az élőlények a környezetből anyagokat és energiát vesznek fel, ill. adnak le.
Az élőlények a felvett anyagokat átalakítják:
- beépítik, felépítő folyamataikban használják fel, vagy
- energianyerés céljából lebontják, majd
- a felesleges, ill. a fel nem használható anyagokat (salakanyagokat) eltávolítják.
A felvétel és a leadás közötti átalakító folyamatok összességét intermedier - köztes - anyagcserének nevezzük, melynek köszönhetően az élőlények a szüntelen változó környezetben fenntartják egyediségüket, viszonylagos állandóságukat, rendezettségüket, működésük rugalmas változásával.
Az élőlényeket az anyagcseréjükhöz szükséges
- szénforrás, ill.
- az energiaszerzés formája szerint lehet csoportosítani.
Szénforrás szerint
Autotrófok: azok az élőlények melyek a testük felépítéséhez szükséges anyagokat az élettelen környezetből veszik fel. A felvett anyagok mindig szervetlen anyagok:
- CO2,
- H2O,
- NO2-, ill. nitrátion - NO3- stb. (ásványi sók).
Tehát a szerves vegyületeik C-tartalma közvetlen a CO2-ból származik (autosz = önmaga, trófosz = táplálkozó, gör.). Az autotrófok a szerves vegyületeik előállításához szükséges energiaforrás típusa szerint lehetnek:
a) fototrófok: ahol az energiaforrás a Nap fényenergiája. A folyamat a fotoszintézis, melynek során az élőlények szén-dioxidból és vízből a Nap energiájának segítségével saját testük felépítéséhez szükséges szerves anyagokat állítanak elő. A folyamat mellékterméke az oxigén.
Ilyenek a fotoszintetizáló baktériumok, algák, ill. a zöld növények.
b) Kemoszintetizálók (autotróf kemotrófok): a szerves vegyületek felépítéséhez szükséges energia valamilyen szervetlen anyag oxidációjából származik, a folyamat a kemoszintézis. Erre kizárólag baktériumok képesek.
A talajban élő aerob nitrifikáló baktériumok ammónia - NH3 – oxidálásából szabadítanak fel energiát, a folyamat végterméke nitrition - NO2-, ill. nitrátion - NO3-
Heterotrófok: testük felépítéséhez szükséges anyagokat az élő környezetből veszik fel szerves anyagok formájában, s ennek
- egy részének lebontásával nyernek energiát, ezért kemotrófok is,
- más részét átalakítják saját testük anyagaivá.
Ilyenek az állatok, gombák, heterotróf egysejtűek, heterotróf baktériumok.
Energiaforrás szerint
Csoportosíthatjuk az élőlényeket aszerint is, hogy a testük felépítéséhez szükséges anyagok - szerves anyagok - előállításához honnan nyerik az energiát.
Fototrófok, lásd fent.
A fototróf élőlények csak autotrófok lehetnek: baktériumok, egysejtű algák és növények.
Kemotrófok
A szerves anyagaik előállításához különböző forrásból származó kémiai energiát használnak fel. Anyagokat bontanak le, oxidálnak, ezzel nyernek energiát.
Attól függően, hogy milyen anyagok átalakításával nyernek energiát, a kemotróf élőlények lehetnek:
- autotrófok, ha az energia szervetlen anyagok oxidálásából származik (nitrifikálók). Ekkor a folyamat a kemoszintézis.
- Heterotrófok, ha az energia szerves anyagok lebomlásából származik, mint pl. a gombáknál, ill. az állatoknál.
Az intermedier anyagcserét, irányát tekintve, két fő folyamatrendszerre oszthatjuk:
- asszimiláció vagy felépítő folyamatok (anabolikus folyamatok), melyek többnyire redukáló jellegűek (redukciónak nevezzük a hidrogén felvételével, ill. oxigén leadásával járó reakciókat),
- disszimiláció vagy lebontó folyamatok (katabolikus folyamatok), melyek sokszor oxidáló típusúak (oxidációnak nevezzük a hidrogén leadásával, ill. az oxigén felvételével járó reakciókat).
Az asszimiláció
Az asszimiláció lényege, hogy a szervezetek egyszerű felépítésű, kis méretű molekulákból – szerves vagy szervetlen - bonyolult, nagy méretű szerves vegyületeket állítanak elő.
- Ha a kiindulási vegyület szervetlen, akkor autotróf asszimilációról beszélünk.
- Fototróf asszimiláció növényekben, egyes baktériumokban (fotoszintézis).
- Kemotróf asszimiláció kemotróf baktériumokban (kemoszintézis).
- Ha a kiindulási vegyület szerves, akkor heterotróf asszimilációról beszélünk (állatokban, gombákban, heterotróf baktériumokban). (A növények is képesek saját szerves anyagaik átalakítására, de ettől még autotrófok.)
A felépítő folyamatok során, végső soron a képződött monomerekből kondenzációval makromolekulák jöhetnek létre, mint pl. glükózból keményítő vagy cellulóz, aminosavakból fehérjék, nukleotidokból pedig nukleinsavak.
A disszimiláció
A disszimiláció lényege, hogy az élőlények bonyolult, nagy méretű, szerves vegyületeiket kis méretű, egyszerű molekulákká bontják.
- Amennyiben a lebontás oxigén jelenlétében zajlik, biológiai oxidációról beszélünk.
- Abban az esetben, ha a lebontás oxigén hiányában történik, erjedésről van szó.
A lebontás célja:
- egyrészt az energianyerés – ATP-szintézis - a különböző életfolyamatokhoz, pl. mozgás, felépítő folyamatok, aktív transzport (a lebontás során felszabaduló energia kb. csak 40%-a fordítódik ATP szintézisre, a többi hővé alakul),
- másrészt az anyagátalakítás, mivel a lebomlás köztes termékei kiindulásként szolgálnak különböző szerves vegyületek szintéziséhez. Az anyagátalakító folyamatok ugyanakkor jelentős energiaveszteséggel járnak, mivel pl. a glükóz biológiai oxidációjának hatásfoka csupán 40%, azaz a glükóz energiatartalmának 40%-a épül be ATP-be, a többi hővé alakul. Továbbá a felépítő folyamatok energiaigényét az ATP bomlásakor felszabaduló energia fedezi, aminek során a keletkező energia jelentős része veszteségként szintén hő formájában távozik.
A lebontó folyamatok első lépése többnyire a makromolekulák hidrolízise monomerekké, amely lejátszódhat mind a tápcsatorna üregében, mind a sejtek citoplazmájában:
- a keményítő és a glikogén glükózzá,
- a fehérjék aminosavakra,
- a nukleinsavak nukleotidokra bomlanak.
A monomerek további átalakítása már a sejteken belül játszódik le.
13. tétel: A Sejtosztódás |
A kromatinállomány
Nem osztódó sejtekben a sejtmag állománya elektronmikroszkópos felvételeken világosabb és sötétebb területekből áll. Ezeken belül, ún. kromatinrostok figyelhetők meg, amelyek DNS-ből és fehérjékből állnak. A rostok együttese a kromatinállomány.
A kromatin elemi egységei a nukleoszómák, melyek hisztonkorongokra feltekeredett hiszton-DNS komplexek.
A kromatinnak festődés alapján két állományát különböztetjük meg.
- Az erősebben festődő részt heterokromatinnak,
- a világosabb területeket eukromatinnak nevezzük.
Az eltérő festődés különböző szerkezetre vezethető vissza.
- A lazább, kitekeredett szerkezetű eukromatin a DNS működő formája, ezeken a helyeken folyik a transzkripció.
- A tömörebb szerkezetű heterokromatin a DNS inaktív formája, ezeken a szakaszokon átírás nincs.
A heterokromatinban a nukleoszómafüzér egy spirál mentén tekeredik fel, ahol már az egyes nukleoszómák érintkeznek egymással, tehát igen sűrűn helyezkednek el.
A sejtosztódások közötti időszakban - az interfázisban - a kromatinállomány többnyire laza szerkezetű, a DNS egyes szakaszairól – a génekről - intenzív átírás folyik, a sejt működik (eukromatin).
A kromatinállomány szerkezete azonban a sejtosztódás ideje alatt jelentősen megváltozik, erősen spiralizálódik, sűrűsödik, a DNS-állomány igen kis területen csomagolódik össze.
A kromatinállomány eme erősen felcsavarodott formáját hívjuk kromoszómának.
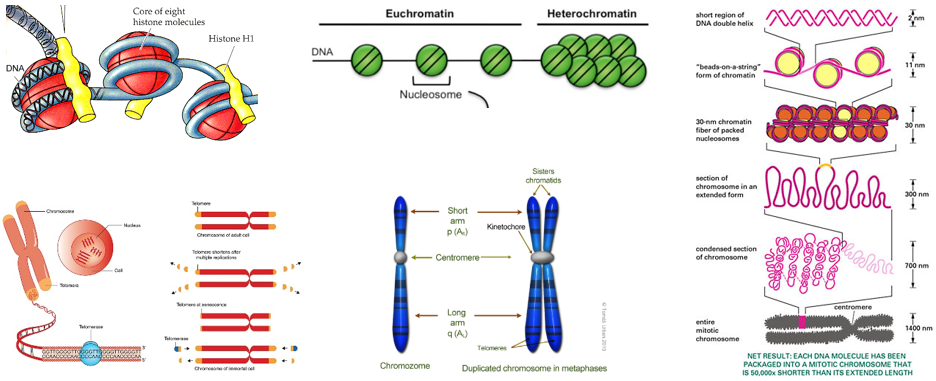 |
A kromoszóma szerkezete
A sejtosztódás idejére kialakuló kromoszóma az előzőleg megkettőződött kromatinállomány – DNS-állomány - transzportformája. A jelentős sűrűsödés lehetővé teszi az egyébként rendkívül hosszú DNS mozgatását a sejten belül.
Az eukarióta kromoszóma két DNS-molekulából áll, amelyek egyetlen DNS megkettőződésével jönnek létre. A két DNS-molekula jól láthatóan elkülönül egymástól és a két ún. kromatidát alkotja, melyek genetikai tartalma teljesen megegyező. A szemikonzervatív replikáció mechanizmusa miatt kromatidák DNS kettős hélixeiben az egyik szál a régi DNS-ből származik, a másik szál újonnan szintetizálódik.
Az osztódás alatt a kromoszómák jól láthatók, megrövidülésük ekkor éri el a legnagyobb mértéket, ezért szerkezetük vizsgálata ekkor a legalkalmasabb (metafázisban).
Alak
Minden kromoszómán megfigyelhető egy elvékonyodás, az ún. elsődleges befűződés vagy másképpen centromer.
A kromoszómák végeit telomereknek nevezzük. A telomerek genetikai információt nem hordoznak, az a szerepük, hogy védjék a kromoszómák információt hordozó részeit a sérülésektől, ill. az összetapadástól az osztódások során, mivel minden egyes osztódáskor a kromoszómák végei töredeznek, megrövidülnek.
A kromoszómák száma
Fajra jellemző és mindig állandó. Általában nincs összefüggés a kromoszómaszám és az illető faj fejlettsége között (ember testi sejtjeiben 46, kutyáéban 78 db.). A kromoszómaszám lehet
- egyszeres, ahol minden kromoszómából csak egy van, az ilyen sejteket haploid sejteknek nevezzük, jelölésük:
- A kromoszómaszám lehet kétszeres, ahol minden kromoszómából kettő van, azaz két kromoszóma azonos alakú, méretű és ugyanazokat a géneket hordozzák. Az ilyen sejtek diploid sejtek, jelölésük: A kromoszómapárok tagjainak egyike apai, a másik anyai eredetű. Ezeket homológ kromoszómáknak nevezzük.
- Állatokban az ivarsejtek - petesejt, hímivarsejt - mindig haploidok, a testi sejtek diploidok.
- Növényeknél a spórák és a belőlük fejlődő ivaros nemzedéknek sejtjei és az ivarsejtek mindig haploidok, a testi sejtek lehetnek haploidok (mohák), diploidok (harasztok, nyitvatermők, zárvatermők) egyaránt.
Váltivarú szervezetekben az egyik kromoszómapár két tagja - általában hímnemű egyedekben – alakilag, méretileg eltér egymástól. Ezeket a kromoszómákat ivari kromoszómáknak nevezzük, ezek a nemiség meghatározói.
A faj haploid sejtjeire jellemző kromoszómák összességét kromoszómaszerelvénynek nevezzük. A haploid sejtekben egyszeres, a diploid sejtekben kétszeres a kromoszómaszerelvény.
A megtermékenyítés során egy haploid hímivarsejt és egy haploid petesejt olvad össze, így a létrejövő zigóta és a belőle keletkező sejtek diploidok, azaz kromoszómakészletük minden kromoszómát két példányban tartalmaz. A két kromoszóma a diploid sejtekben homológ kromoszómapárt alkot, melyben a hímivarsejtből származó kromoszóma az apai, a petesejtből származó kromoszóma az anyai kromoszóma. A homológ kromoszómapárok tagjai azonos géneket tartalmaznak, ugyanakkor ezen belül többnyire eltérő allélokat.
A sejtciklus
A soksejtű szervezetek kifejlett állapotban nagyszámú és sokféle testi sejtből épülnek fel, pl. egy átlagos súlyú felnőtt emberi szervezet kb. 1O14 (1OO billió) sejtből áll. Ezek folyamatosan elhasználódnak, pusztulnak, ezért állandóan pótlódniuk kell.
A szervezet sejtjei azonban nemcsak sokasodnak, hanem növekednek és fejlődnek is.
Egy testi sejt miután létrejött két fejlődési út elé nézhet:
- Soha többé nem osztódik, megindul a differenciálódás útján, specializálódik valamilyen szöveti sejtté alakul (G0 fázis). Ez jellemző sejtjeink többségére.
- A másik lehetőség szerint, a sejt rövid pihenőt követően ismét osztódik, amikor is az osztódás és az osztódások közötti időszakok szabályosan követik egymást. Ebben az esetben sejtciklusról beszélünk. Ez jellemző az embrionális sejtekre, ill. az állandóan osztódó sejtekből álló szövetekre, mint pl. vörös csontvelő, bélhám, többrétegű elszarusodó hám, ivarszervek csírahámjai, továbbá ilyenek a tumorsejtek.
A sejtciklus alapvetően két részre osztható:
- az osztódás fázisára,
- az osztódások közötti ún. interfázisra.
Az interfázisos sejtben
- a maghártya élesen elkülöníti a mag állományát,
- melynek belsejét a kromatinállomány tölti ki.
Az osztódó sejtben
- a maghártya feldarabolódik, majd eltűnik,
- a kromatinállomány kromoszómákká tömörödik, a kromoszómák jól felismerhetőek.
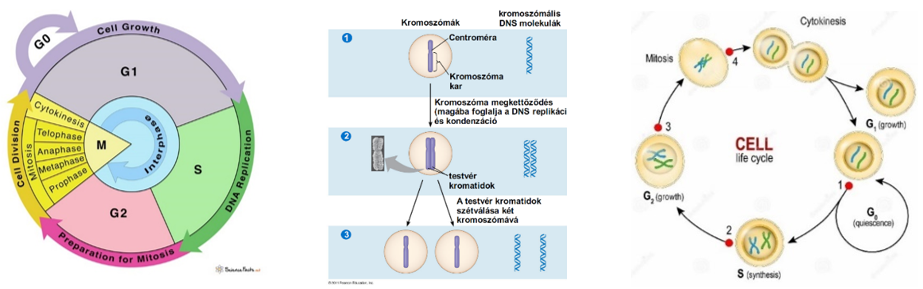 |
Az interfázis
Az interfázis alatt a sejtben számos olyan folyamat zajlik, amely a sejtosztódást készíti elő. Az interfázist három szakaszra osztjuk:
- G1 szakasz, (G, angol gap = (idő)rés)
- S szakasz,
- G2 szakasz.
A G1 szakasz az osztódást közvetlen követi. A létrejött utódsejtek:
- növekednek,
- a sejtszervecskék megsokszorozódnak,
- lemerült energiaraktáraikat feltöltik.
Azok a sejtek, amelyek már nem osztódnak tovább, a G1 szakaszból a G0 szakaszba kerülnek, ahol a sejt determinálódik és valamelyik differenciálódási pályán indul el. Maga a determináció, az elköteleződés azt a pillanatot jelenti, amelyben az illető sejt egy specifikus fejlődési pályára lép. Ez egy adott genetikai program beindulását jelenti.
A differenciálódás a sejtek sokféleségének az alapja, amelynek során az eredetileg egyforma sejtek más és más speciális képességekre tesznek szert.
Az S fázisban történik:
- a replikáció, melynek során 46 DNS molekulából - melyek egykromatidás kromoszómák - 92 DNS-molekula lesz (46 kétkromatidás kromoszóma),
- tehát létrejönnek a kromoszómák másik kromatidái.
A kromoszómák az interfázisban is léteznek, csak ilyenkor a laza kromatinállományt alkotják, melyben a kromoszómák csak egyik kromatidája van jelen. Az S fázisban az egykromatidás kromoszómák DNS-e megduplázódik, és a keletkező másolat a kromoszóma másik kromatidáját alkotja. A kétkromatidás kromoszómákban tehát a két kromatida – a másolási hibáktól eltekintve – teljesen azonos információtartalmú.
A G2 fázis
- Rövid nyugalmi szakasz, amelyben a sejt létrehozza azokat az anyagokat, amelyek az osztódáshoz kellenek (pl. húzófonalak fehérjéit). Mind a G1, mind a G2 szakaszra intenzív fehérjeszintézis jellemző.
- A sejtközpont kettéosztódik, a sejt két pólusára vándorol.
A sejtek szaporodása
Az eukarióta sejtek osztódással szaporodnak. Az osztódás lényegesebb formái a mitózis és a meiózis.
A mitózis
A mitózis fonalas sejtosztódást jelent (mitosz = fonál). A mitózis, más néven számtartó sejtosztódás lényege, hogy az interfázisban megkettőződött DNS - melyek a kromoszómák egyes kromatidáit alkotják – egyenlő mértékben megoszlik a két utódsejt között, két utódsejtmag jön létre, amelyekben ugyanolyan számú kromoszóma van, mint az osztódás előtti magban volt. A számtartó sejtmagosztódás bonyolult, általában néhány óra alatt lejátszódó folyamat.
Mitózissal osztódnak:
- a növények testi sejtjei,
- állatok testi sejtjei, azaz a zigóta és az abból létrejövő sejtek.
A mitózis folyamatán belül 4 fázist különböztetünk meg.
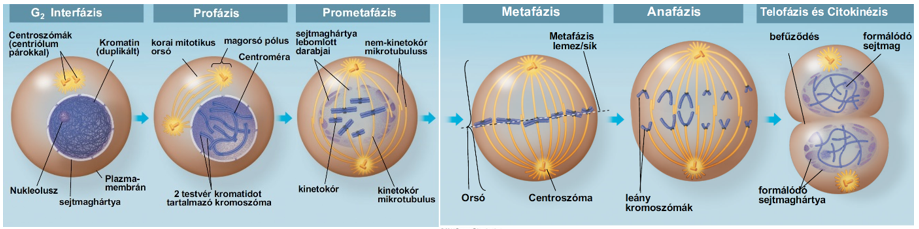 |
Előszakasz, profázis
- A sejtmagvacska eltűnik, a maghártya kezd hólyagokká feldarabolódni.
- A kromatin fokozódó tömörödése miatt láthatók lesznek a kromoszómák.
- A már kettéosztódott sejtközpontok a sejt két ellentétes pólusára vándorolnak.
Középszakasz, metafázis
- Ekkorra a kromoszómák a legnagyobb mértékben rövidülnek,
- a kromoszómákat a sejt középső síkjában rendezi el,
- a kromoszómák centromérjeihez húzófonalak kapcsolódnak, ez a magorsó.
Utószakasz, anafázis
- A húzófonalak a kromatidákat a sejt két pólusára mozgatják.
Végszakasz, telofázis
- A pólusokra érkezett kromatidák fokozatosan kitekerednek,
- a magorsó eltűnik, új maghártya szerveződik.
Az örökítő anyag szétosztását a citoplazma osztódása követi, amelynek során a sejthártya befűződik, a sejt kettéválik, a sejtszervecskék kb. egyenlő mértékben elosztódnak az utódsejtek között. A folyamat a citokinézis.
Minden mitózis után két utódsejt keletkezik, melyek genetikailag azonosak egymással és az anyasejttel (mutációtól, másolási hibáktól eltekintve).
Genetikai szempontból legfontosabb eseménye az, hogy a két kromatidából álló kromoszómák kromatidái (DNS kettős hélixei) az osztódás során szétválnak, majd az utódsejtekbe kerülnek. Az utódsejtekben tehát ismét egy kromatidából (egy DNS kettős hélixből) álló, de változatlan kromoszómaszámú kromoszómakészlet található.
Meiózis
- A meiózis az állati ivarsejtek, növényi, ill. gombaspórák termelésére specializálódott diploid sejtek osztódástípusa.
- A meiózis során egyetlen diploid anyasejtből 4 haploid utódsejt keletkezik.
- Az utódsejtek kromoszómaszáma - ezáltal genetikai információtartalma - pontosan fele az anyasejtének (számfelező sejtosztódás).
- A keletkezett négy haploid utódsejt információtartalma egymástól különböző.
A meiózis két főszakaszra bontható.
- A meiózis első szakaszának profázisában a homológ kromoszómák párt alkotnak, kromatidáik között átfedések, törések, majd szakasz – gén - kicserélődések következhetnek be. Ennek a folyamatnak, az átkereszteződésnek (crossing over) az eredménye az intrakromoszómális rekombináció. A rekombináció során a tulajdonságokat kialakító genetikai tényezők újrarendeződnek, így új génkombinációk a szülőktől eltérő fenotípusokat eredményeznek.
Az átkereszteződést követően a homológ kromoszómapárok tagjai szétválnak egymástól és külön utódsejtbe kerülnek, mialatt a testvérkromatidák még együtt maradnak (számfelezés). Véletlenszerű, hogy az egyik ill. másik utódsejtbe a kromoszómapárok apai vagy anyai eredetű kromoszómája fog vándorolni.
- A második osztódási szakaszban a testvérkromatidák válnak szét egymástól, így az utódsejtek kromoszómánként egy kromatidából (egy DNS kettős hélixből) álló, de az anyasejthez képest fele kromoszómaszámú kromoszómakészlettel rendelkeznek.
Első főszakasz
Profázis I:
- A homológ kromoszómák hosszuk mentén párt alkotnak, így egy komplexet 4 db. kromatida alkot.
- Átkereszteződések jelennek meg a kromatidák között. A törések következtében a homológok egyik kromatidájában kromoszómaszakaszok cserélődhetnek ki.
Metafázis I A homológok az egyenlítői síkban rendeződnek, a homológok centromereihez az ellenkező pólushoz tartozó húzófonalak tapadnak.
Anafázis I A homológ párok tagjai az ellenkező pólusra jutnak (számfelezés!).
Telofázis I Kialakul a sejtmembrán, de DNS-szintézis nincs!
Az első osztódás redukciós, mert a kromoszómák számát a felére, a haploid számra (n) csökkenti az utódsejtekben.
Második főszakasz
- Lényegében egy mitózis.
- A kromoszómák az egyenlítői síkba rendeződnek.
- A kromatidák szétválnak.
- A húzófonalak az ellentétes pólusokra viszik a kromatidákat.
- 4 haploid utódsejt keletkezik.
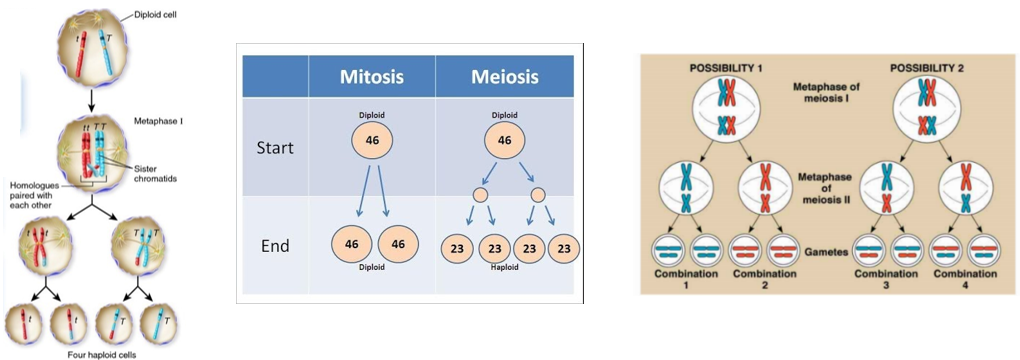 |
Az utódsejtek genetikai változatossága
A rekombináció során olyan haploid génkombináció jön létre, amely különbözik a szülői génkombinációtól. 2-féle rekombinációt különbözetünk meg:
Intrakromoszómális – kromoszómán belüli rekombináció.
A meiózis profázisában történik, a homológ kromoszómák egy-egy kromatidája között.
Az interkromoszómális független rekombináció. Szintén az első főszakaszban zajlik a homológ kromoszómák véletlenszerű szétválásának köszönhetően.
- Egy kromoszómapár: 2 lehetőség van.
- Kettő kromoszómapár: 4-féle ivarsejt,
- három kromoszómapár: 8-féle ivarsejt,
- N kromoszómapár: 2N-féle ivarsejt jöhet létre.
- Ember: 23 pár kromoszóma,
- 223 » 8400000-féle ivarsejt képződhet.
A rekombináció és így az ivaros szaporodás az alapja a genetikai változatosságnak, a populációk genetikai diverzitásának, amely az evolúció genetikai alapját teremti meg.
Az ivaros szaporodás során az alábbi események a véletlenszerű tulajdonságkombinációk „végtelen” lehetőségét biztosítják:
- az ivarsejtek véletlenszerű találkozása,
- az átörökítő DNS átrendeződése (átkereszteződés),
- a homológok szétválása,
- illetőleg a DNS szerkezetében beálló változás (mutáció).
14. tétel: Transzportfolyamatok |
A biomembránok – és így a sejthártya - biztosítják az elhatárolt egység, a sejt működéshez szükséges belső környezet állandóságát. A membránok egyrészt fizikailag elhatárolnak, másrészt azonban a rajtuk keresztül folyó, szabályozott anyagmozgás révén egyben össze is kapcsolnak.
Ha az anyagmozgás a sejten kívüli és a sejten belüli tér között a membrán síkján való áthatolással történik, akkor membrántranszportról beszélünk. A membrán transzportfolyamatai csoportosíthatók:
a membránon való áthatolás mechanizmusa szerint:
- vizes csatornán át történő diffúzió,
- lipidfázison át zajló diffúzió,
- a membránban lévő szállítók segítségével (lehet passzív és aktív egyaránt),
a membránon át történő anyagmozgás energiaigénye szerint:
- passzív, ahol a folyamat energiát nem igényel, a transzport a koncentráció-gradiensnek megfelelő irányba zajlik, eredményeképpen a koncentrációkülönbség kiegyenlítődik,
- aktív, ahol a folyamat energiát igényel, az anyagtranszport a kisebb koncentrációjú hely felől történik a nagyobb felé. Koncentrációkülönbséget hoz létre.
Vizes csatornán át történő diffúzió
Vizes közegben történő diffúzióval mozognak a membránon át
- az ionok,
- a kisebb poláris molekulák és a víz.
Az átjutásnak ez a módja azt bizonyítja, hogy a lipidtartalmú membránon át vizes csatornák, ún. pórusok húzódnak. A csatornákat integráns membránfehérjék alkotják, amelyek szerkezetváltozása a csatornákat nyitja vagy csukja. A különböző ionok különböző ioncsatornákon jutnak át, így ismerünk Na+-, K+-, Ca2+-, Cl- stb. csatornákat. A folyamat energiát nem igényel, azaz passzív.
Lipidfázison át zajló diffúzió
A zsíroldékony anyagok a membrán lipidfázisában oldva jutnak át a membránon. Ilyen módon jut át az oxigén, CO2, szteroidhormonok, alkohol stb.
A membránban lévő szállítók segítségével
Az ide sorolható folyamatok közös jellemzője, hogy a szállított anyag kötődik egy, a membránban lévő szállítómolekulához, s ennek segítségével jut át a szállítófehérje szerkezetváltozása révén. A folyamat lehet passzív vagy aktív.
Facilitált – könnyített - diffúzió
A szállított anyag mindig a koncentrációgradiense által megszabott irányba mozog diffúzióval, passzív transzporttal, szállítók segítségével. A szállított anyag így gyorsabban jut át, mint a molekula méretéből, lipidoldékonyságából, koncentráció-gradienséből következne.
A sejtbe bejutott anyagok igen gyorsan hígulnak, ami biztosítja a folyamatok egyirányúságát. Pl. a glükóz gyorsan foszforilálódik, a glükóz-foszfát a szállítóval már nem kapcsolódik.
Segített diffúzióval veszik fel a különféle szöveti sejtek a glükózt. Többféle – 14 - glükóztranszporter ismert (GLUT).
Inzulin hatására a GLUT-1 transzporterek mellé, az addig a sejt belsejében, vezikulákban tárolt GLUT-4 transzporterek beleolvadnak a külső membránba, s megnövelik a zsírsejtek és az izomrostok glükózfelvételét.
Aktív transzport
Az aktív transzportfolyamatok egyenlőtlen anyageloszlást hoznak létre a membrán két oldala között. Mivel az anyag a koncentrációgradiens által meghatározott spontán mozgási iránnyal szemben mozog, az aktív transzport mindig E-t igényel.
Az energia ATP hidrolíziséből származik. Az aktív transzport mindig szállítókkal történő transzport. Energiaigényes szállítófolyamatok pl. a különféle ionokat szállító pumpamechanizmusok. Kapacitásuk véges, bizonyos sebesség felett működésük tovább nem fokozható (telítődési görbe).
Pumpamechanizmusok
A membránpumpák olyan transzportrendszerek, amelyek egy adott anyagot a membrán egyik oldalán felhalmoznak, míg a membrán másik oldalán a folyadék az adott anyagban elszegényedik. A ma ismert pumpamechanizmusok ionszállító transzportrendszerek, amelyek egyben ATP-bontó enzimek is.
H-ionok aktív transzportja (uniport)
A protonpumpák minden élő sejtben megtalálhatók. Igen fontos működést látnak el:
- mitokondrium-, kloroplasztisz-membránban,
- vesecsatornák hámsejtjeiben,
- gyomornyálkahártya egyes sejtjeiben.
Na+-K+- pumpa (ATP-áz) (antiport)
Az állati sejtek legfontosabb ionpumpája. A sejtek anyagcseréje során keletkező ATP-mennyiség 20-30%-át ez az enzim használja fel. A sejt nyugalmi állapotában 1 ATP hidrolízise során 3 Na+-ot pumpál ki és 2 K+-ot visz be a sejtbe.
A pumpa működése döntően fontos:
- a sejt ozmotikus egyensúlyának és ezzel a sejt térfogatának és alakjának fenntartásában, (ha nem működik, a sejt vizet vesz fel, duzzad, kipukkadhat),
- a membránpotenciál szinten tartásában,
- biztosítja a sejten belüli enzimműködések számára fontos magas K+- koncentrációt és távol tartja a legtöbb folyamatban gátló Na-ionokat.
Glükóz-Na+ kotranszport (csatolt anyagtranszport, szimport)
Ismertek olyan szállítók a membránban, amelyek kétféle anyagot is képesek egyszerre átjuttatni a sejthártyán. Ebben az esetben csatolt transzportról – kotranszport - beszélünk. Ilyen módon veszik fel a tápcsatornában a bélhámsejtek a glükózt.
A glükóz felvétele aktív transzport, az alacsonyabb koncentrációjú hely felől a magasabb koncentrációjú hely felé szállítódik, amihez az energiát a Na+ koncentrációgradiense által meghatározott diffúziója biztosítja. Tehát a glükóz energetikailag kedvezőtlen irányba megy, a Na+ pedig kedvezőbe, de végső soron a kettő eredője kedvező.
A mechanizmus másodlagos aktív transzport, hiszen a folyamat feltétele a Na+ koncentrációkülönbsége a sejthártya két oldalán, melyet a Na+-K+ pumpa hoz létre ATP felhasználásával.
Endo- és exocitózis
Olyan anyagok (baktériumok, makromolekulák stb.) sejtbe való bejutása – endocitózis -, ill. sejtből való kijutása - exocitózis - is lehetséges, amelyek pl. méretüknél fogva a membrán síkján nem képesek áthatolni. Az így szállított anyagok membránnal körülvett hólyagocskákba csomagolva jutnak át a sejthatáron. Mivel a folyamatok során a membránok átrendeződnek, ezért energiaigényesek.
Endocitózis
Az endocitózis biológiai jelentősége:
- az egysejtűek táplálkozásában,
- immunfolyamatokban, az ún. sejtes védekezésben van.
Amennyiben a bekebelezendő anyag szilárd halmazállapotú fagocitózisról beszélünk.
A fagocitózis lépései
- Adszorpció, amelynek során a bekebelezendő anyag megkötődik a membrán felszínén levő receptorokon.
- A kötődés hatására a sejtváz befelé húzza a membránrészletet a receptorral és a megkötődött anyaggal együtt. Az ilyen módon a sejtbe került kis hólyagocskát fagoszómának nevezzük.
Exocitózis
Exocitózissal a sejt belsejéből igen különböző eredetű és természetű anyagok ürülhetnek.
Az exocitózis biológiai jelentősége:
- mirigysejtek váladékürítésében (emésztőenzimek, hormonok),
- egysejtűek salakanyag-leadásában,
- idegsejtek ingerületátvivő anyagának ürítésében van.
15. tétel: Vírusok, prionok |
A vírusok felépítése
A vírusok fertőzőképes, kórokozó sejtparaziták, makromolekuláris rendszerek, melyek nem érik el a sejtes szerveződés szintjét, nem tekinthetők élőlényeknek, az élő és az élettelen világ határán állnak.
Méret
Nanométeres nagyságrendűek, méretük 20 - néhány száz nm-ig terjed (1nm = 10-9 m), csak elektronmikroszkóppal láthatók néhány 10 000 x-es nagyítás mellett.
Szerkezet
Nem érik el a sejtes szerveződési szintet, csupán kétféle anyagból állnak.
- A vírus belső részében található az örökítőanyag, amely lehet DNS vagy RNS (retrovírusok). Az örökítőanyag feladata a vírus felépítésére vonatkozó információ tárolása.
- Perifériásan egy fehérjeburok található, melyet toknak nevezünk. Feladata az örökítőanyag védelme, ill. lehetővé teszi a megtapadást a gazdasejten.
A vírusok szaporodása
A vírusok nem tekinthetőek sem élőnek, sem élettelennek, az élő és az élettelen rendszerek határán állnak.
A vírusnak két megjelenési formája ismert:
- virion a sejten kívüli fázis, amely élettelen, kristályos szerkezetű.
- A sejten belüli vegetatív fázis, amely a működő, szaporodó vírus.
A vírusok önmagukban semmilyen életjelenséget nem mutatnak, nincs anyagcseréjük. A vírusok kizárólag azáltal képesek szaporodni, hogy megtámadnak valamilyen sejtet, behatolnak, átszervezik a sejt működését, arra kényszerítik, hogy a saját anyagait felhasználva, a bejutott vírushoz hasonló vírusokat hozzon létre. A folyamat a sejt pusztulásához vezet. Ez a jelenség a parazitizmus vagy élősködés.
A vírus által megtámadott sejtet gazdasejtnek nevezzük. A gazdasejt típusától függően beszélünk:
- emberi: influenza, koronavírusok, kanyaró, nátha, bárányhimlő, AIDS, veszettség, rubeola, herpesz, hepatitisz, HPV,
- állati: sertés és baromfi pestis, száj és körömfájás, veszettség,
- növényi vírusokról,
- bakteriofágokról.
A vírusok többsége gazdaspecifikus, azaz kizárólag a rá jellemző gazdaszervezetben képes szaporodni. Kivételek is ismertek, ilyen pl. a veszettség, száj és körömfájás, tehénhimlő stb. vírusa.
A vírus tehát
- nem élő, mert a gazdasejt nélkül nem képes szaporodni, nincs önálló anyagcseréje.
- De nem is élettelen, mivel a gazdasejtbe jutva, annak anyagait felhasználva szaporodni képes, megváltoztatja a gazdasejt működését.
A fertőzés folyamata, bakteriofágok litikus és lizogén ciklusa
- A virion a közeg áramlásával jut el a gazdasejthez, mivel önálló mozgásra képtelen.
- A virion megtapad a gazdasejt felületén. A megtapadás specifikus víruskötő helyeken (receptorokon) történik. A receptorok mindig az adott sejttípusra jellemzőek, így ezért gazdaspecifikusak a vírusok.
- Bakteriofágok esetén a megtapadás fehérjefonalakkal történik, majd a talpon található enzim (lizozim) kilyukasztja a sejtfalat. A lyukon keresztül a DNS a sejtbe injektálódik, a burok kívül marad.
A sejten belül a vírusok kétféleképpen viselkedhetnek:
- Litikus életciklus: a bejutás után azonnal megkezdődik az új vírusok képzése. A sejt elpusztul, szétszakad.
- Lizogén életciklus: a vírus DNS beépül a gazdasejt DNS-ébe. Ilyenek pl. az RNS tumorvírusok, herpesz vírusok, a HPV, HIV. A beépülés után a vírus sokáig észrevétlen maradhat, majd még nem tisztázott okok miatt – pl. immunrendszer legyengülése - aktívvá válhat.
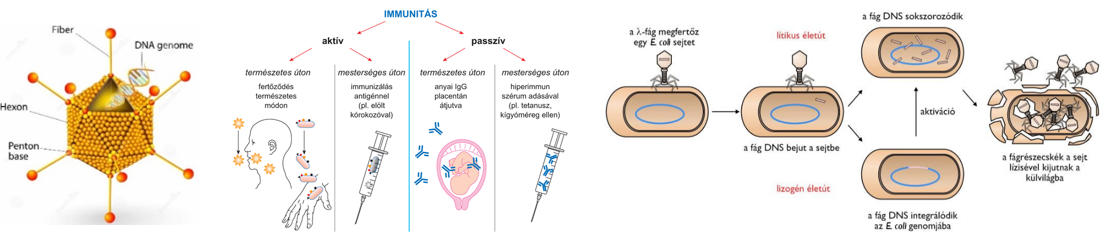 |
A vírusbetegségekről (lásd még 4.8.5. fejezet)
A tudomány jelenleg olyan vírusellenes gyógyszert nem ismer, amely megfertőződés esetén a vírusokat maradéktalanul megsemmisíti. Ugyanakkor pl. a hepatitis-C és a HIV ellen ismertek már olyan hatóanyagok, melyeknek köszönhetően a vírus gyakorlatilag eltűnik a testnedvekből, miáltal ún. tartós vírusmentesség érhető el.
Egyrészt a virion nem élő szervezet, ezért a szervezeten belül elpusztítani nem lehet, másfelől a vírus vegetatív formája a gazdasejtben van, így csak a sejttel együtt lehetne megsemmisíteni, azonban a gyógyszerek nem tudják a fertőzött sejteket megkülönböztetni az egészségesektől.
A vírusbetegségekkel szembeni védekezés leghatékonyabb módja a megelőzés. Ez többek között immunizálással, ill. higiénés szabályok betartásával lehetséges.
Az immunizálás történhet
- mesterségesen, különféle védőoltásokkal,
- természetes módon,
továbbá lehet
- aktív vagy
- passzív, attól függően, hogy az immunizálandó szervezet immunrendszere részt vesz-e a folyamatban vagy sem.
A vírusbetegségek jellegzetes tünetei a magas láz, levertség, izom és ízületi fájdalom. A vírusok legjobban 35-36 Celsius fokon szaporodnak, így a láz nem kedvez a fertőzésnek.
A magasabb rendű eukarióta sejtekben az evolúció során kiépültek különféle védekező mechanizmusok a vírusokkal szemben.
Elsősorban növényekre jellemző az RNS interferencia jelensége. A növényi vírusok egy része kettős szálú RNS genomot tartalmaz. A sejtekbe bejutó virális eredetű kettős szálú RNS-szakaszok egy nagyobb, RISC nevű fehérjekomplexbe épülnek be. Ez a komplex a későbbiekben minden olyan bejutó vírus mRNS-t lebont, amely szekvenciája hasonlít a korábban beépült vírus RNS-hez.
Másik példája a vírusokkal szembeni védekezésnek az emberi szervezetben az interferonok működése. Az interferonokat a vírussal fertőzött sejtek bocsátják ki. Az interferonok a szomszédos sejtek receptoraira kapcsolódnak, és aktiválják azokat a védekező mechanizmusokat, amelyek segítenek a fertőzés leküzdésében. A még egészséges sejtekben olyan enzimek képződését váltják ki, amelyek akkor keletkeznek, mikor bekövetkezik a fertőzés. Ezek az enzimek pl. bontják a vírus mRNS-t, gátolják a fehérjeszintézist stb.
 |
Fontosabb emberi vírusos betegségek
Influenza
Az influenzát magas láz, orrfolyás, köhögés, fejfájás, rossz közérzet, az orr és a légutak nyálkahártyájának duzzanata, végtagfájdalom jellemezi. Az influenzavírusok az egyéb légúti fertőzések kórokozóihoz hasonlóan a köhögéskor, tüsszögéskor és beszéd közben keletkező légúti váladékcseppecskék révén, cseppfertőzéssel terjednek, elsősorban a tüdő hámsejtjeit támadják meg. A megbetegedés elsősorban a légzőrendszert és a keringési rendszert érinti. Szövődményei lehetnek tüdőgyulladás, szívizomgyulladás, agyvelőgyulladás. A beteg leggyakrabban magától meggyógyul, javasolt az ágynyugalom, valamint a megfelelő folyadékpótlás.
A kórokozó a burkos vírusok közé tartozik, ismert A, B és C típusa is. A burokban különféle H és N jelű molekulák találhatók, amelyek alapján a vírusok tovább csoportosíthatók, pl. H1N1. s ez az alapja. Minden évben újra és újra kialakulnak az influenzajárványok, mivel a felszíni molekulák szerkezete gyakran megváltozik, s így az immunrendszer nem ismeri fel a korábban már leküzdött kórokozót.
A vírus állatok közvetítésével nagyobb genetikai változáson is átesik időnként, ami az ember immunrendszerét felkészületlenül érheti, ezért világméretű, igen súlyos járványok alakulhatnak ki, mint pl. 1918-ban a spanyolnátha (egyesek szerint madárinfluenza).
A 2020. évben azonosított új koronavírus (SARS-CoV-2) okozta fertőzések (COVID-19)
Forrás: koronavirus.gov.hu
A koronavírusok alapvetően állatok körében fordulnak elő, ugyanakkor egyes koronavírus törzsekről ismert, hogy képesek az emberben is fertőzést okozni. A koronavírusok mind állatról emberre, mind emberről emberre terjedhetnek. A mostani járványt okozó új koronavírus 2019 végén került azonosításra Kínában. Az új koronavírus elnevezése „súlyos akut légúti tünetegyüttest okozó koronavírus 2” (SARS-CoV-2), az általa okozott megbetegedés pedig a „koronavírus-betegség 2019” (Coronavirus Disease 2019), melynek rövidített elnevezése a COVID-19. Ez a vírustörzs a kínai Vuhanban a 2019 decemberében kitört járvány előtt ismeretlen volt.
A koronavírusok lipid burokkal rendelkező, egyszálú RNS vírusok. A koronavírus fertőzések okozta megbetegedések változó súlyosságúak lehetnek: a hétköznapi náthától a súlyosabb, halálos kimenetelű légúti megbetegedésekig.
Terjedés módja: az emberről-emberre történő terjedés jellemzően cseppfertőzéssel és a fertőzött váladékokkal történik.
Az új koronavírussal történő fertőződés esetén a legtöbb emberben kialakul a megbetegedés. Ritkák az olyan esetek, ahol tünetmentesen zajlik le a fertőzés (becsülten 1-3%).
A WHO adatai szerint a megbetegedés jellemzően lázzal, száraz köhögéssel, fáradékonysággal jár. Előfordul nehézlégzés, torokfájás, fejfájás, izom- vagy ízületi fájdalom, hidegrázás. Ritkább tünet a hányás, hasmenés.
A betegség leggyakrabban (kb. 80%-ban) enyhe vagy közepesen súlyos formában zajlik, ekkor a klinikai kép az enyhe légúti fertőzéstől a nem súlyos tüdőgyulladásig terjedhet.
A halálozás 2-3% között változik, amelyet jelentősen befolyásol a vírus terjedésének intenzitása, illetve az érintett terület jellemzői (pl. demográfiai mutatók, egészségügyi ellátáshoz való hozzáférés), általában legmagasabb az idősek körében.
A betegség súlyos formája leginkább a 60 évnél idősebbeket és a krónikus alapbetegségben (pl. magas vérnyomás, cukorbetegség, szív- és érrendszeri, krónikus légúti megbetegedés, daganatos megbetegedések) szenvedőket érinti.
 |
AIDS
Acquired Immuno Deficiency Syndrome (Szerzett Immunhiányos Tünetegyüttes)
Az AIDS betegség vírusa a HIV, amely az immunrendszer sejtjeit támadja meg és pusztítja el, aminek következtében néhány év alatt a szervezet teljesen kiszolgáltatottá válik a különféle fertőző megbetegedéseknek. Igen súlyos betegség, kezelés nélkül halállal végződik.
A betegség 1980-as években, az USA-ban fiatal homoszexuális férfiak körében jelent meg. Később fertőzött vérkészítményekkel, közös injekciós tű használattal, prostitúcióval terjedt tovább. Mára világméretű járvánnyá vált, jelenleg kb. 37 millió vírusfertőzött ismert. A vírus valószínűleg majmokból – SIV- került át az emberbe Afrikában.
A HIV szerkezete
- Retrovírus, azaz RNS az örökítőanyag, amely a sejtekben a reverz transzkriptáz enzimmel DNS-sé másolódik.
- Tok (kapszid).
- Peplon: kettős foszfolipid membrán, gp41 és gp120 glikoproteidek, melyek segítségével a CD 4 jelű receptorokat hordozó fehérvérsejtekhez képes kötődni.
A vírusfertőzés folyamata
- Vírus megtapad a sejt felületén, a peplon összeolvad a sejthártyával, majd az örökítőanyag a tokkal bejut a gazdasejtbe.
- A vírus RNS a reverz transzkriptáz segítségével DNS-sé íródik át. Ez a DNS beépül a gazdasejt DNS-ébe.
- A gazdasejt elkezdi a beépült DNS alapján a vírus alkotórészeit „legyártani”.
- A kész vírusok kiszabadulnak a sejtből (több százezres nagyságrendben). A gazdasejt általában elpusztul.
A fertőzés terjedése
- Védekezés nélküli szexuális úton.
- Fertőzött vérrel vagy vérkészítménnyel.
- Anyáról gyermekre (méhen belüli fertőzés, szülés és szoptatás során).
- Injekciós intravénás kábítószer által.
- Nem megfelelően sterilizált orvosi eszközök révén.
Nem terjed…
- kézfogással,
- puszival,
- rovarcsípéssel,
- együtt fürdéssel stb.
A tünetek
HIV-fertőzés lefolyása 3 szakaszra osztható.
- Kezdeti fertőzés,
- tünetmentes szakasz,
- súlyos immunelégtelenség (=AIDS!).
- 3-6 héttel a fertőzés után influenzára emlékeztető tünetek jelentkezhetnek, esetleg nyirokcsomó megnagyobbodás tapasztalható.
- A vírus elleni antitestek a fertőzés után 1 hét - 3 hónap múlva mutathatók ki.
- A fertőzés után akár évekig tartó lappangási szakasz következhet (1-15 év), ekkor már HIV pozitivitásról beszélünk, a vírus a szervezetből nem ürül ki.
- Amikor az immunsejtek száma egy bizonyos érték alá csökken, kialakul az AIDS betegség.
Ez hajlamot jelent ún. opportunista fertőzéses betegségekre, azaz minden olyan kórokozó, ami a normál immunitású emberekre veszélytelen, az AIDS betegeknél súlyos, akár életveszélyes állapotot is elő tud idézni, pl. tüdőgyulladás, hasmenés, gombásodás, TBC, hepatitis-B, hepatitis-C stb., egyes daganatos megbetegedések kockázata sokszorosára növekszik, mint pl. a Kaposi-szarkómáé (lásd jobb felső kép).
Hepatitis, fertőző májgyulladás
Májgyulladást (hepatitis) okozhatnak vírusok, autoimmun betegség, alkohol, bizonyos gyógyszerek és méreganyagok. Világszerte a vírusos májgyulladás a leggyakoribb. Szövődménye lehet májzsugor, vagy rosszindulatú daganat.
Általános tünetek
Jobb bordaív alatti fájdalom, a bőr és szemfehérje sárgás elszíneződése, sötét vizelet, fáradtság és levertség, hasi panaszok. A beteg máj nem képes a lépből származó bilirubint megfelelő módon és mértékben feldolgozni, átalakítani (vízoldhatóvá tenni), ezért az felszaporodik a vérben, a szövetekben és a bőrben.
A jelenleg ismert hepatitis vírusokat A-tól G-ig jelzik és X-szel jelölik a még nem azonosított vírusokat. A vírusok a kórokozó típusától függően két fő formában terjednek:
- a hepatitis A és E a tápcsatornán keresztül (például fertőzött élelmiszer, fertőzött széklet),
- a hepatitis B, C, D, G és a többi vírus vérrel és testnedvekkel (ondó, hüvelyváladék).
A hepatitis A és E
A betegséget a beteg és a tünetmentes fertőzött személy székletével szennyezett kéz, tárgyak, élelmiszerek, ivó- és fürdővíz terjesztheti. Tünetei a néhány napig tartó láz, elesettség, étvágytalanság, sárgaság.
Hepatitis B és C
Szexuális úton, vérrel, vérkészítményekkel, testváladékokkal, ondóval, hüvelyváladékkal terjed. A tünetmentes hordozók és a krónikus betegek életük végéig fertőzőképesek lehetnek! A fertőzés késői következménye a májzsugor és a májrák.
A HBV ellen aktív védőoltás javasolt, ami a 14 éves korosztály számára kötelező és ingyenes.
A HCV rendkívüli mutációs készsége miatt hatásos védőoltás jelenleg még nincsen.
HPV (Human Papillomavirus)
A HPV szexuális úton terjedő kórokozó, mely rendellenes sejtburjánzás következtében kialakuló szemölcsöket okozhat a szervezet külső hámfelületein, a bőrön, a nemi szerveken és a végbélnyílás körül. Rendkívül elterjedt kórokozó, a népesség mintegy fele élete során átesik HPV fertőzésen, de nagy részük a fertőzést leküzdi, így nem alakul ki benne semmilyen tünet. A HPV kórokozók eddig több mint 130 ismert típusát azonosították, melyeket daganatkeltő képességük alapján különböző kockázatú csoportokra osztották. A magas kockázatú vírusok olyan daganatos betegségek kialakulásában játszhatnak közre, mint a méhnyakrák, a hímvessző, a hüvely rákja, bizonyos szájüregi rákok, valamint a végbél laphámrákja. A HPV vírusok genetikai anyagukat beépítik a gazdaszervezet örökítő anyagába (lizogén életciklus).
Kanyaró
A kanyaró lázas, hurutos (hurut: a nyálkahártyák erős váladéktermelésével járó gyulladása), kiütéses, igen fertőző súlyos betegség.
Rózsahimlő vagy rubeola
A rubeola egy enyhe – gyengeséggel, fejfájással és hőemelkedéssel járó -, cseppfertőzéssel terjedő gyermekkori kiütéses betegség. Azonban a várandós nők megbetegedése a terhesség korai szakaszában súlyos magzati károsodást okoz, mint pl. veleszületett szívbetegség, a teljes vagy részleges vakság, a kétoldali süketség, szellemi visszamaradottság.
Bárányhimlő
A bárányhimlő cseppfertőzéssel terjedő, piros, viszkető, hólyagos pattanásokat és lázat okozó megbetegedés. Rendkívül fertőző. Gyermekkorban leggyakoribb szövődmény a hólyagok másodlagos fertőződése miatt kialakuló bőrfertőzés. A betegségen átesett embereknél később kialakulhat övsömör, mert ez a betegség a bennük lappangó vírus reaktivációja miatt jöhet létre.
Herpesz
A herpesz vírusa elsősorban a bőrön, ill. nyálkahártyákon okoz tüneteket, ritkán az idegrendszert betegíti meg. Kis területen égő, viszkető hólyagocskák jellemzik. Többnyire ártalmatlan, viszont makacsul visszatérő megbetegedés, mivel a vírus lappangó formában megmarad a szervezetben (lizogén életciklusú). A tünetek megjelenésének az oka lehet a szervezet immunrendszerének legyengülése, valamilyen gyulladás vagy lázas betegség, erős lelki trauma, stressz, lelki problémák, más fertőzések stb.
Veszettség
A veszettség vírus okozta halálos kimenetelű fertőző betegség, a vírus veszett állatok testváladékaival, főként nyálával terjed. A vírus a fertőzött személy központi idegrendszerébe jutva, gyakorlatilag minden esetben halálos kimenetelű agyvelőgyulladást okoz. A lappangási idő 3-7 hét.
Nátha
Az orr nyálkahártyájának hurutja – intenzív váladéktermeléssel járó nyálkahártya gyulladás -, a leggyakoribb megbetegedések egyike. A közönséges heveny nátha legtöbbször egyszerű meghűlésből ered, bakteriális, ill. vírusfertőzés okozza.
A vírusok szerepe az ökológiai rendszerek működésében
- A bakteriofágok jelentősen hozzájárulnak a Földön a baktériumok egyedszámának a szabályozásához. A baktériumok elpusztításával óriási tömegű szerves anyagot termelnek, mely más heterotróf lebontó szervezetek számára fontos tápanyagot jelent. Tehát a vírusok részt vesznek az ökoszisztémák anyag- és energiaforgalmában, mivel közvetve hozzájárulnak a szerves anyagok szervetlen anyagokká – szén-dioxid, víz, ásványi anyagok stb. - alakulásához.
- A vírusok szerepet játszanak a gazdaszervezetek populációi egyedszámának a szabályozásában. Túlnépesedés esetén a vírusok járványokat okozhatnak, amelyek csökkentik a populáció egyedszámát. Ez segít fenntartani az ökológiai egyensúlyt és megakadályozza a populációk túlnépesedését.
Prionok
A prionok (PrPSc) olyan fertőző, kórokozó fehérjék, melyek különféle neurodegeneratív betegségek, mint pl. emberben a Jakob-Creutzfeldt betegség (fertőző szivacsos agyvelősorvadás), szarvasmarhákban a szivacsos agyvelősorvadás (kergemarhakór, szivacsos agyvelőgyulladás) kialakulásában játszanak szerepet. A kórfolyamat előrehaladtával a központi idegrendszer sejtjei elhalnak, az agyban üregek jönnek létre, állománya szivacsossá válik.
A prion betegségre jellemző, hogy nincs ellenanyag termelés, nincs gyulladásos reakció, a lappangási idő évekig tarthat.
A prionoknak nincs örökítő anyaguk, semmilyen életjelenséget nem mutatnak, a vírusoknál is egyszerűbb képződmények. A prionok az egyébként normálisan is meglevő sejtes fehérjék – celluláris prion (PrPc) – szerkezetileg módosult változatai.
A celluláris prionok (PrPc) általában idegsejtek felszínén a membránban találhatók, az idegsejtek egészséges működésében játszanak szerepet. A normál sejtes prionok vízoldékonyak, érzékenyek lebontó enzimekkel szemben.
A kóros prionok (PrPSc) aminosavsorrendje nem, de a másodlagos szerkezete – konformációja - eltér a normálisan is meglevő sejtes fehérjétől, s emiatt bontó enzimekkel szemben nem érzékenyek, vízben pedig nem oldódnak. Feltételezik, hogy a prionok (PrPSc) a sejtekbe jutva elősegítik – katalizálják - a normális fehérjék prionná átalakulását, melyek felhalmozódva, a hidrofób felszínükkel összecsapódva, oldhatatlan rögöket képeznek és az idegsejtek elhalását okozzák.
16. tétel: Baktériumok |
A prokarióták csoportjai
A biológia tudománya az élők világát legújabban három alapvető doménre – birodalomra - osztja:
- a valódi baktériumokra,
- az ősbaktériumokra,
- az eukariótákra (valódi sejtmagvasok).
A valódi baktériumok és az ősbaktériumok a prokariótákhoz tartoznak, a legegyszerűbb felépítésű, már sejtes szerveződést mutató élőlények.
A prokarióták (legszembetűnőbb) közös sajátsága, hogy sejtjeikben nincs membránnal körülhatárolt sejtmag. (Pro=előtt, karion=mag gör.). Ezen kívül persze számos eltérés van e két birodalom között, a pro- és az eukarióták között nagyobb a különbség, mint az állatok és a növények között, a törzsfejlődés során kb. 2,5 milliárd évre volt szükség ahhoz, hogy a prokarióták kialakulását követően az első eukarióták megjelenjenek.
Földünk legrégibb ismert élőlényei. Legkorábbi geológiai maradványaik a sztromatolitok, kb. 3,4 milliárd évesek. A sztromatolitok réteges, párnaszerű üledékes kőzetek, amelyek egyes ősi kékbaktériumok sejtjei körüli mészkiválás eredményeképpen jöttek létre.
Abban az időben a földi légkör nem tartalmazott oxigént, tehát az ekkor élt baktériumok anaerobok voltak. Kb. 2,5 milliárd évvel ezelőtt terjedtek el tömegesen azok a fotoszintetizáló aerob baktériumok, amelyek lassan oxigénnel töltötték meg a légkört.
 |
Az ősbaktériumok az élővilág korai formáit képviselő szervezetek. Többnyire szélsőséges körülmények között élnek.
Az általuk kedvelt élőhely alapján három csoportba szokás osztani őket:
- termofilek meleg élőhelyeken, például hőforrásokban fordulnak elő.
- a halofil fajok extrém sós környezetben élnek,
- a metanogén fajok anaerob környezetben – kérődzők gyomrában, mélytengerekben - fejlődnek és ecetsavból vagy H2-ből CO2-al metánt termelnek. A szennyvizek tisztításában, a biogáz előállításában (metántermelők) jelentős szerepet játszanak.
A Föld őstörténetének egy hosszú periódusában az élővilág fő tömegét valószínűleg ők alkották. Az ősi elnevezés arra utal, hogy egyes feltételezés szerint ezek a szervezetek az eubaktériumoknál előbb léteztek. Az ősbaktériumok törzsfejlődéstanilag azonban sok tekintetben közelebb állnak az eukariótákhoz, mint a baktériumokhoz. Genetikai anyaguk működésének (átírás, fehérjeszintézis) hasonlósága alapján az eukarióták ősi ágának is tekinthetők.
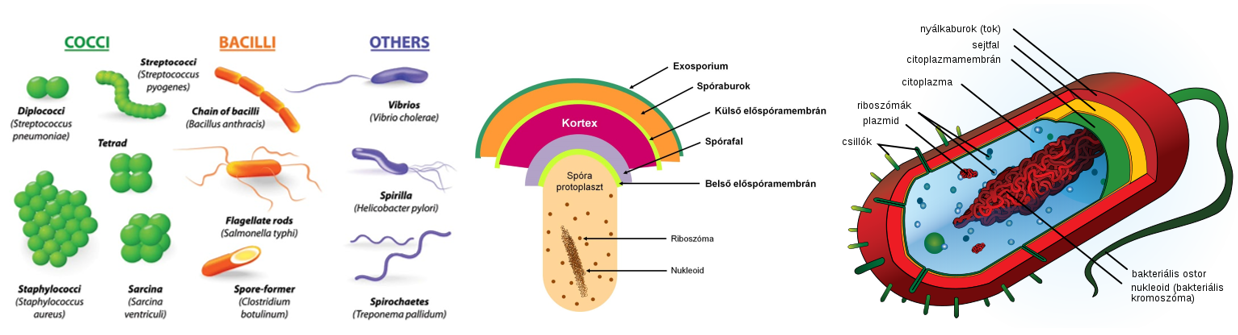 |
Az eubaktériumok (valódi baktériumok)
Az eubaktériumok a hagyományos értelemben vett baktériumok. A Föld minden élőhelyén megtalálhatóak: vízben, szárazföldön vagy a levegőben, még mélytengeri hőforrásokban is.
Egy gramm talaj kb. 40 millió, egy ml felszíni víz egymillió baktériumsejtet tartalmaz. A Földön pedig összesen mintegy 5 kvintillió (5 × 1030) baktérium élhet. Tízszer annyi baktérium van az emberi testben, mint emberi sejt. A legtöbb baktérium a bőr felszínén és az emésztőrendszerben található.
Méret
Már fénymikroszkóppal láthatók néhány 100 x-os nagyítás mellett. Átlagos méretük 1 -10 mikrométer (1µm = 10-6 m).
1 mikrométer átmérőjű kokkuszokat véve, 1 mm3-ben 109 sejt található, amelyek összfelülete 3.1 négyzetméter. Egy felnőtt ember testfelülete 1.9 négyzetméter.
A baktériumok szerveződése
A sejtek magányosak vagy osztódás után együtt maradva laza kolóniákat, ún. sejttársulást hozhatnak létre.
Alak
3 nagyobb csoportba oszthatók:
- gömb (kokkuszok)
- pálcika (bacillusok)
- görbült pálca, amin belül pl.
- csavar (spirillumok)
- félhold (vibrió)
A sejtek felépítése
Sejtfal
Minden baktériumot kívülről sejtfal borít, amely biztosítja a sejt állandó alakját és egyben véd is.
Kedvezőtlen körülmények között spórát (endospórát) képeznek: a plazma vizet veszít, összezsugorodik, újabb falat képez az örökítőanyag és egy kis plazma köré. Tehát a baktériumok spórája nem ivartalan szaporítósejt, mint a növényeknél, hanem egy hőnek, sugárzásnak és a roncsoló hatású kémiai anyagoknak ellenálló képződmény. Pl. a lépfene baktérium így a 130 Celsius fokot is elviseli.
A baktériumok penicillin érzékenysége is a sejtfalnak köszönhető, ugyanis a penicillin a bakteriális sejtfal képzését gátolja.
Tok
Egyes baktériumoknál a sejtfalon kívül még egy nyálkás, kocsonyás tok található. A tok feladata
- a védelem az immunrendszerrel szemben, védi a sejtet a bekebelezéstől,
- szerepet játszik a tápanyag megkötésében,
- segítségével összetapadhatnak a sejtek (sejttársulás).
Sejthártya
Vékony hártya (foszfolipid kettősréteg), mely a sejtfal alatt található, lebonyolítja az anyagforgalmat a sejt és környezete között.
Citoplazma (sejtplazma)
A sejt alapállománya, a sejtanyagcsere színtere. Itt található a DNS állomány (genom), amely egy vagy több gyűrűs DNS-ből áll, nem határolódik el membránnal a citoplazmától. Ezt a DNS-t baktérium kromoszómának is nevezzük.
Ezen kívül a sejtplazmában vannak még kisebb DNS gyűrűk, a plazmidok, melyeken többnyire az antibiotikumokkal szembeni ellenállóképességet lehetővé tevő antibiotikum rezisztencia gének találhatók, továbbá riboszómák, a fehérjeszintézis színhelyei.
Csillók
A baktériumok egy része ún. csillók segítségével képes mozogni. A bakteriális csillók vékony, rendszerint a sejt hosszát akár többszörösen is meghaladó hosszúságú, fehérjékből felépülő sejtfelszíni képződmények.
A csillók száma baktériumfajtól függően egy és néhány száz között változhat. A csillók kémiailag és szerkezetileg is eltérnek az eukarióták hasonló funkciójú képződményeitől.
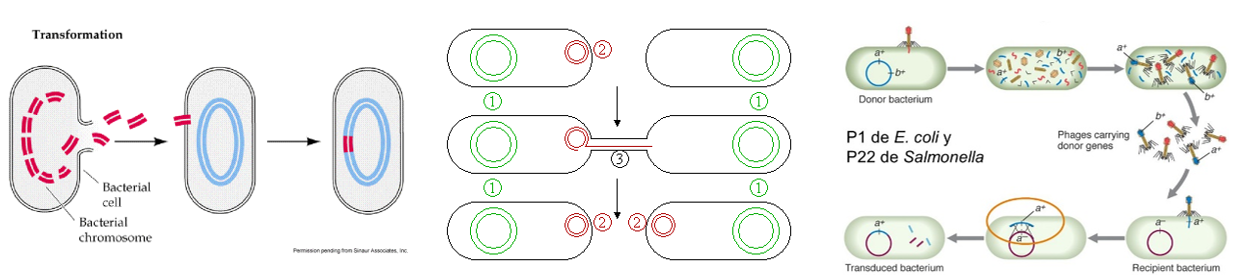 |
Szaporodás
A prokarióták nagyon gyorsan tudnak szaporodni. A sejtek 20 percenként kettéosztódnak, tehát számuk 20 percenként a kettő hatványainak sorozatában nő.
Ha egyetlen baktériumból indulunk ki, és feltételezzük, hogy a belőle keletkező utódok minden 20-dik percben kettéosztódnak, akkor 48 óra alatt a Földnek megfelelő tömeg jönne létre (2144 db sejt).
A prokarióták a legegyszerűbb módon főleg hasadással szaporodnak, amikor is a sejt egyszerűen kettéfűződik. A folyamatot ivartalan szaporodásnak tekintjük, mivel az osztódást nem előzi meg 2 sejt egyesülése.
A baktériumoknál hiányzik az ivaros szaporodásnak az eukariótáknál megszokott formája. Nincs meiózis, mitózis és nincs az ivarsejtek teljes összeolvadásához hasonlító megtermékenyítés.
Ugyanakkor ismertek olyan ún. ivaros folyamatok, melynek során a baktériumok bizonyos körülmények között képesek más egyedekből vagy fajokból származó DNS szakaszok felvételére, aminek következtében a sejtek rekombinálódnak és új tulajdonságokra tehetnek szert.
A DNS-átvitel történhet transzformáció, konjugáció és transzdukció útján. Mindhárom esetben a génátadás egyirányú.
A transzformáció
A transzformáció során a sejt a környezetéből idegen DNS-t vesz fel és a rajta elhelyezkedő gént vagy géneket beépíti saját genetikai anyagába. A DNS darabnak a sejtfalon és a sejthártyán való áthaladását specifikus fehérjék segítik elő. Így adhatja át az egyik baktérium a másiknak, pl. az antibiotikumokkal szembeni ellenállóképességet (rezisztenciát).
Az ivaros folyamat után a megváltozott információtartalmú – rekombinálódott - sejtek hasadással szaporodnak tovább.
Konjugáció
Két baktérium között egy plazmahíd jön létre. A konjugáció során a plazmid megkettőződik és a hídon keresztül a „hím” (donor) baktérium DNS-t ad át a „nő” (recipiens) baktériumnak, megváltoztatva annak genetikai készletét.
Transzdukció
A transzdukció a génátvitelnek az a módja, amelyben az egyik baktériumsejtből a másikba bakteriofágok segítségével jut át genetikai információ. A vírus DNS és a baktérium DNS-e kombinálódik, az új vírusrészecskék tartalmazzák a donor bakteriális DNS-t, melyet a kiszabaduló vírusok visznek át a következő baktériumba. A molekuláris genetikában a génátvitel egy bevált módszere.
Életmód
Az élőlényeket – így a baktériumokat is –
- a szénforrás, ill.
- az energiaszerzés szerint - azaz az anyagcsere szempontjából lehet csoportosítani.
Szénforrás szerint az élőlények lehetnek:
Autotrófok: melyek a testük felépítéséhez szükséges anyagokat az élettelen környezetből veszik fel. A felvett anyagok mindig szervetlen anyagok: CO2, H2O, NH3 (autosz=önmaga, trófosz=táplálkozó, gör., növekedésükben nincsenek más élőlényre utalva).
Az autotróf baktériumok az energiaforrás típusát tekintve lehetnek:
- fototrófok: ahol az energiaforrás a nap fényenergiája. A folyamat a fotoszintézis, mikor az élőlények szén-dioxidból és vízből a nap energiájának segítségével saját testük felépítéséhez szükséges szerves anyagokat állítanak elő.
A folyamat mellékterméke lehet oxigén. Ilyenek pl. a kékbaktériumok.
- Kemoautotrófok, kemoszintetizálók: a szükséges energia valamilyen szervetlen anyag oxidációjából származik. Ezek a baktériumok fényhiány ellenére is életképesek.
A nitrifikálók az ammóniát oxidálják nitrit-, ill. nitrát-ionná, amiből nyerik a szerves anyagok felépítéséhez szükséges energiát.
Heterotrófok: testük felépítéséhez szükséges anyagokat az élő környezetből veszik fel szerves anyagok formájában, s ennek egy részének lebontásával nyerik az energiát, más részét átalakítják saját testük anyagaivá. Ide tartoznak heterotróf baktériumok.
Energiaforrás szerinti csoportosítás
Csoportosíthatjuk az élőlényeket aszerint is, hogy a testük felépítéséhez szükséges anyagok (szerves anyagok) előállításához honnan nyerik az energiát.
- Fototrófok, lásd fent.
A fototróf élőlények lehetnek baktériumok, ill. eukarióta egysejtű algák és növények.
- Kemotrófok
A szerves anyagaik előállításához különböző forrásból származó kémiai energiát használnak fel. A kemotróf élőlények lehetnek:
- autotrófok, ha az energia szervetlen anyagok oxidálásából származik (nitrifikálók).
- Heterotrófok, ha az energia szerves anyagok lebomlásából származik, mint pl. a gombáknál, ill. az állatoknál (kemoorganotrófok).
A heterotróf baktériumokat életmód szerint tovább csoportosíthatjuk:
Szaprofiták: az elpusztult élőlények szerves anyagait bontják le, alakítják vissza CO2-á és vízzé, lehetővé téve az autotrófok által történő újrafelhasználást (szaprosz=korhadt, fita=növény, gör.). A szaprofiták nélkül megállna az anyagok körforgása, pl. az összes szén az elpusztult élőlényekben halmozódna fel. Ebbe a csoportba tartoznak a korhasztó különféle talajbacik és az erjesztő baktériumok, mint pl. a tejsavbaktériumok, amelyek a tejcukrot tejsavvá bontják le.
Szimbionták: amikor egy baktérium egy másik élőlénnyel olyan együttélésben él, amely mindkét fél számára előnyös. A prokarióták több olyan különleges anyagcsereúttal rendelkeznek, amelyekkel a magasabb rendű élőlények nem (a légköri nitrogén felhasználása, cellulóz bontása). Az eukarióták ezeket a képességeiket igénybe veszik, cserébe a baktériumok szaporodásához megfelelő körülményeket teremtenek.
- Pillangósvirágú növények gyökérgümőiben élő nitrogénkötő baktériumok,
- a növényevő állatok beleiben élő cellulózbontó baktériumok.
- Ember beleiben coli baktériumok.
Paraziták A parazita baktériumok különféle betegségeket okoznak.
A baktériumok okozta betegségekről lásd még 4.8.5. fejezet
A baktériumos fertőzés jellemző tünetei: láz, fejfájás, hányás, hasmenés, gyulladások, melyeket a baktériumsejt által termelt méreganyagok (baktériumtoxinok) váltanak ki. A különböző baktériumok különböző toxinokat termelnek, és ezeknek a toxinoknak különböző hatásmechanizmusuk van. A baktériumtoxinok lehetnek
- az idegi jelátviteli folyamatokat megzavaró neurotoxinok.
Például a legerősebb ismert természetes toxin a Clostridium botulinum által termelt botulin neurotoxin (botox) egy grammja egy millió ember halálát okozhatja. A baktérium okozta élelmiszermérgezések egyik legsúlyosabb formája a botulizmus. A baktérium oxigéntől elzárt környezetben, talajban él, s az állatok feldolgozása során a baktérium a nyers húsra kerülhet, és a feldolgozott termékekben hőkezelés hiányában elszaporodva botulotoxint termel. A táplálékkal az emberi szervezetbe jutott kórokozó méreganyaga a bélfalon keresztül kerül a vérbe. A vérből a környéki idegvégződésekhez kötődik, ahol gátolja az ún. acetilkolin ingerületátvivő anyag felszabadulását az ideg-izom kapcsolódásokban.
Amennyiben botox kezeléssel meggátoljuk a ráncképződésért felelős izmok működését, elérhetjük, hogy a ráncképződés megszűnjön, az arcbőr kisimuljon. A hatás csak időleges, a kezelést 3-4 havonta meg kell ismételni.
- A sejtek enzimműködését károsító citotoxinok.
- A bélműködés zavarát okozó enterotoxinok, mint pl. a Vibrio cholerae baktérium által termelt toxin, ami a kolera nevű betegségért felelős. A toxin súlyos hasmenést és hányást okoz, amelyek súlyos dehidratációhoz vezethetnek.
Szalmonella: ételmérgezés, rendszerint fertőzött étel fogyasztása után kialakuló gyomor-bélrendszeri betegség. A baktérium a szájon át kerül be a szervezetbe szennyezett élelmiszerrel. A fertőzés magas lázzal, görcsös hasfájással, hányással, hasmenéssel jár. A kezelés során fokozottan ügyelni kell a folyadék pótlására a kiszáradás elkerülésének érdekében.
A fertőzés a nyári hónapokban gyakoribb, mert ilyenkor a meleg hatására a nem megfelelően tárolt élelmiszerekben a baktérium könnyen elszaporodik. A kórokozó a széklettel ürül, a nem megfelelő higiénés viszonyok, a kézmosás hiánya elősegítheti lakóközösségen belüli terjedését. Ritkán előfordulhat társuló ízületi gyulladás, ha a baktérium a véráram útján eljut az ízületekbe.
Tbc (tuberculosis): elsődlegesen a tüdő szövetének pusztulásával járó betegség. A betegség jellemzően a tüdőben gyulladással kezdődik, de később az egész szervezetre kiterjedhet. A megelőzés elődleges eszköze Magyarországon az újszülöttek BCG-oltása.
Tüdőszűréssel az esetlegesen kezdődő folyamatok jól kimutathatók, és idejében, hatékonyan gyógyíthatók.
Ha a kórokozók a vérárammal szétszóródnak, és más szervekben is megtelepednek, tüdőn kívüli szervi tuberkulózis alakulhat ki, melyek a következők lehetnek:
- A csont-tbc csontszövetelhalást, kóros csonttöréseket okoz.
- A vese tbc a vese szöveteinek elhalásával jár.
- A gyomor-bél- tbc A tüdőből felköhögött fertőző váladék lenyelése is okozhatja. A bélcsatornában körkörösen elhelyezkedő fekélyek formájában mutatkozik meg, mely hegesedés következtében bélelzáródáshoz vezet.
- A szív-tbc a szívburkok megbetegedése.
A tetanusz egy, a talajban élő anaerob baktérium toxinja által okozott merevgörccsel járó bénulás, mely kezeletlen esetben halálos. A kórokozó a szervezetbe földdel szennyezett sérüléseken keresztül tud bejutni. A baktérium méreganyaga az ideg-izom jelátvitelben szereplő anyag – acetilkolin – lebomlását gátolja, miáltal súlyos izomgörcsök alakulnak ki. Az izommerevség gyorsan terjed fentről lefelé, folyamatosan egyre több izmot érint, végül a légzőizmok görcse fulladásos halált okoz. A lappangási idő átlagosan egy hét.
A pertussis - szamárköhögés különböző kórokozók által okozott tünetegyüttes, amelyet rohamokban jelentkező, súlyos, kezdetben hurutos, később "szamárordításszerű” köhögés jellemez. Az életet veszélyeztető idegrendszeri és légúti szövődményei főként csecsemőkorban fordulnak elő, egyébként a betegség magától gyógyul.
A diftéria vagy torokgyík ún. álhártyás gyulladás, amely leginkább a mandulákon, ritkábban a gégében alakul ki. A betegség lassan, fokozatosan fejlődik ki, a láz nem jellemző. Súlyos szövődményként szívizom-károsodást és ideggyulladást okoz, illetve, ha az álhártya a gégében jelenik meg, ez fulladáshoz vezet.
A Lyme-kór kullancs által terjesztett bakteriális fertőzés, amely először bőrtünetekkel, később, kezeletlen esetekben pedig ízületi és izomfájdalmakkal jár, majd gyakran idegrendszeri elváltozásokat okoz. Közép-Európa a világ egyik legfertőzöttebb területe. Magyarországon évente kb. 10 000 friss fertőzés fordulhat elő. Különösen fertőzött a Budai-hegység, a Balaton-felvidék és a nyugat-magyarországi területek.
A legelső tünet általában a csípés körül megjelenő, legalább 5 cm-esre megnövő, többnyire ovális alakú bőrpír.
Magyarországon 2, 3, 4, 18 hónapos, majd pedig 6 és 11 éves korban a gyerekek kombinált DTP védőoltást (diftéria= torokgyík, tetanusz= merevgörcs, pertussis=szamárköhögés) kapnak. Ugyanakkor levegőtől elzárt, mély, szúrt, földdel szennyezett sérülések esetén szükséges ún. emlékeztető oltás az immunrendszer válaszának erősítése céljából.
A baktériumos betegségekkel szemben védőoltásokkal és antibiotikumokkal védekezhetünk.
Magyarországon 2019-ben a következő oltási rend volt érvényben:
- BCG: 0-4 hetes korban
- DTP: 2, 3, 4, 18 hónapos, majd pedig 6 és 11 éves korban.
- Hepatitis B: 12 éves korban.
- Bárányhimlő: 13 és 16 hónapos korban.
A védőoltás – az esetlegesen kialakuló mellékhatások ellenére - a lehető leghatékonyabb mód, hogy az egyes fertőző betegségeket megelőzzük, illetve a súlyos lefolyást megakadályozzuk. A védőoltásokkal megelőzhető fertőző betegségek ritkábban fordulnak elő, mert hosszú ideje nagy tömegeket oltanak ellenük. Az immunizálást mindaddig folytatni kell, amíg a betegségeket fel nem számoljuk. A védőoltásoknak köszönhetően az 1970-es évek végétől nincs feketehimlő a világban, és Európa gyermekbénulás-mentes, kb. 2000 óta. A védőoltásokkal kivédhető fertőző betegségek kórokozói állandóan jelen vannak a környezetünkben, és csak az oltás következtében kialakult védettség miatt nem tudnak betegséget okozni. A már szinte feledésbe merült betegségek újból visszatérnének, ha a mára eltűnt (szakszóval eradikált) betegség védőoltásainak beadásával felhagynánk. Forrás: Wikipédia
A védőoltások kifejlesztése az orvostudomány legnagyobb hatású vívmánya. Manapság mégis sokan ódzkodnak tőle, különféle ideológiákat gyártva. Az egyéni döntés felelőssége azonban nagy, a kórokozók ugyanis terjesztő közegként használnak bennünket, ezért a nem kellő alapossággal meghozott döntésünknek mások is megihatják a levét. Még súlyosabb a morális felelősségünk, amikor a gyermekeink ügyében döntünk.
Az oltásellenesség fő állításai, hogy a védőoltások komoly betegségeket – pl. autizmust - okozhatnak, illetve, a nem oltott gyerekek egészségesebbek, mint a beoltottak. Mindkét állítás hamis. Az ismerethiány szülte oltásmegtagadás sajnos súlyos problémákhoz vezethet, ha ennek szintje elér egy bizonyos arányt a népességben. Sok veszélyes kórokozó várakozik sötét zugokban türelmetlenül, hogy lecsapjon gyermekeinkre és a felnőttekre egyaránt. Az elvégzett számos vizsgálat és megfigyelés szerint a védőoltások kockázata rendkívül alacsony, szemben az általuk biztosított védettséggel, ami viszont igen jelentős. Önmagában az a tény, hogy egy protokoll bizonyos szintű rizikóval rendelkezik, teljesen természetes, hiszen minden másra is igaz. Az élet maga egy veszélyes létezési forma. Forrás: Index
Az antibiotikumok olyan hatóanyagok, melyek a baktériumokat elpusztítják vagy szaporodásukat gátolják. Antibiotikus hatású számos különböző összetételű és szerkezetű anyag lehet. Hagyományosan korábban antibiotikumoknak csak a különféle gombák által termelt hatóanyagokat nevezték. Azóta kiderült, hogy antibiotikus hatású anyagokat nem csak gombák termelnek, hanem pl. baktériumok is, ill. már ismertek mesterségesen előállított ún. szintetikus, ill. természetes antibiotikumok kémiai módosításával készített félszintetikus hatóanyagok is.
Amennyiben egy baktérium ellenáll az antibiotikumnak, úgy rezisztensnek tekintjük a hatóanyaggal szemben. Ennek hátterében a plazmidokon található antibiotikum rezisztencia gének állnak, amelyek olyan fehérjéket kódolnak, amelyek közömbösítik az antibiotikumok hatását, pl. úgy, hogy lebontják azokat. Mivel a plazmidokat a baktériumok ivaros folyamataikban könnyen átadják egymásnak, ezért a rezisztenciagének a baktériumok közötti génátadással terjedhetnek. Emiatt fontos, hogy betartsuk a gyógyszerek szedésére vonatkozó szabályokat. A nem megfelelő mennyiségben szedett, ill. a kezelés idő előtti befejezése a rezisztens baktériumok elszaporodását segíti elő, és így a betegség nem hogy nem gyógyul, de még súlyosbodhat is. Az antibiotikumok a szervezetben rövid idő alatt hatástalanná válnak, ezért szükséges meghatározott időben és a pontosan előírt adagban bevenni őket.
A fertőző betegségek kialakulását, terjedését különféle fertőtlenítési (higiénizálási), sterilizálási eljárásokkal is megelőzhetjük.
A különféle fertőzéses járványokat okozó betegségek és a mikroorganizmusok közötti összefüggés felderítésében és a járványok (lépfene) elleni küzdelemben úttörőszerepet játszott Louis Pasteur (1822-1895) francia vegyész és mikrobiológus.
- Tudományosan először ő bizonyította, hogy a baktériumok különféle fertőzések forrásai.
- Tanulmányozta az erjedést, kimutatta, hogy az erjedést parányi organizmusok okozzák ill., ha nincsenek jelen, az erjedés nem indul meg.
- Bizonyította, hogy hő hatására lejátszódó csírátlanítás megakadályozta az élelmiszerek romlását. Az élelmiszeriparban használt hőkezeléses eljárást feltalálójáról pasztörizálásnak nevezték el.
A másik legismertebb felfedezése a veszettség elleni védőoltás kidolgozása volt.
Kimutatta, hogy ha egy egészséges kutyát beoltanak egy veszett kutya nyúltagyának egy darabjával, az állaton megjelennek a veszettség tünetei. Ugyanakkor fertőzött állatok szárított szöveteivel – nyúlgerinc - kísérletezve Pasteurnek sikerült a vírusból olyan gyengített változatot előállítania, amelyet már felhasználhatott a védőoltásokhoz. Egy menthetetlennek látszó kilenc éves kutyaharapásos elzászi kisfiún tesztelte a vakcinát. A gyermek egészségi állapota a próbaoltás után tíz nappal látványosan javult.
Fertőtlenítés (higiénizálás): minden olyan eljárás, amely a kórokozók számának csökkentésére, elpusztítására irányul.
Sterilizálás: az az eljárás, melynek során a csíramentesítésre kerülő anyagban elpusztítjuk a mikroorganizmusokat, valamint ezek összes nyugvó formáit.
A fertőtlenítés és a sterilizálás között különbség van.
- A sterilizálás célja a teljes csíramentesítés,a sterilezés megöli az összes életképes mikroorganizmust.
- A fertőtlenítés csak csökkenti az életképes mikroorganizmusok számát, fertőzőképességét, az eljárás során a mikrobák spórái életben maradnak és a hatás megszűnte után szaporodni képesek.
A fertőtlenítésnek két alapvető módja ismert.
A fizikai eljárások során vagy hővel vagy különféle sugárzásokkal pusztítjuk el a kórokozókat. A fertőtlenítés hatékonyságát jelentősen befolyásolja, hogy nedves vagy száraz hővel történik–e. A mikroorganizmusok a száraz hőnek jobban ellenállnak, mint a nedvesnek.
A baktériumok spórás alakjai ellenállóbbak, de a forrásban lévő víz hőmérsékletén a legtöbbjük 20 percen belül elpusztul. A vírusok nagy része 50-60 ºC-on néhány perc alatt inaktiválódik. Az autoklávokban a magas hőmérséklet mellett nagy nyomást alkalmaznak, ez a tuti.
A kémiai eljárások során a fertőtlenítőszerek (baktericid, sporocid, virucid, fungicid) hatásukat annak köszönhetik, hogy pl.:
- roncsolják a sejthártyákat, vagy
- a sejtek fehérjéit kicsapják, inaktiválják létfontosságú enzimeit.
- Ilyenek a klór, jód, formalin, hidrogén-peroxid, alkohol.
A sterilizálási módok hasonlóak az egyes fertőtlenítési eljárásokhoz, azaz hővel, gázzal (formaldehid), ionizáló sugárzással történik.
A kórházakban nélkülözhetetlen higiéniára először Semmelweis Ignác (1818-1865) magyar orvos, az „anyák megmentője” hívta fel a figyelmet.
A lelkiismeretesség, a kötelességtudás és az önzetlenség mintaképe.
Szt. Rókus kórház szülészeti osztályának főorvosaként kimutatta a fertőtlenítő eljárások előnyeit a szülészetben és a sebészetben. Rájött, hogy a gyermekágyi lázat az orvosok és orvostanhallgatók okozzák azzal, hogy boncolás után mentek át a szülészeti osztályra, s ott fertőtlenítetlen kézzel vizsgálták a várandós nőket. Fertőtlenítésként klórmeszes kézmosást ajánlott kollégáinak.
Ezzel kezdődött szélmalomharca az akkori hivatalos, tudományos világgal. Mind gyakorlatával, mind írásaiban próbálta terjeszteni nézeteit, de sajnos az orvostársadalom nem vett róla tudomást. Élete végén magatartásában az elmezavar jelei mutatkoztak, Döblingben elmegyógyintézetbe zárták, ahol – ápolói agresszív magaviselete miatt súlyosan bántalmazták – tisztázatlan körülmények között két hét után meghalt.
Miért maradhattak fenn a prokarióták az evolúcióban?
A baktériumok anyagcseréje rendkívül sokféle, számos olyan működésre képesek, amelyekre az eukarióták nem, ezért az élet számára nélkülözhetetlenek.
-
- Nitrogénkötés,
- cellulózbontás.
A fentieknek köszönhetően igen szélsőséges körülményekhez is képesek alkalmazkodni, így fennmaradhattak olyan – pl. oxigén szegény – élőhelyeken, ahol az eukarióták már nem versenyképesek.
Rendkívüli mértékben képesek szaporodni.
Összefoglalva a prokarióták jelentőségét
Környezettani jelentőség:
- Lebontók, elhalt szerves anyagok lebontásában, a talajképződésben fontosak.
- Fotoszintetizáló formáik termelőként a táplálékláncok alapját képezik. Oxigént termelnek.
- A nitrogén-kötő baktériumok többnyire pillangósvirágú (hüvelyes) növényekkel (bab, borsó, lucerna, akác stb.) élnek együtt, képesek a légköri nitrogén (N2) megkötésére, szerves vegyületekbe beépítésére, miáltal a nitrogént hozzáférhetővé teszik a velük együtt élő növények számára.
- Nitrifikáló baktériumok nitritet és nitrátot állítanak elő (ammóniából), ezzel lehetővé teszik a növények nitrogénfelvételét a talajból.
- Vizek öntisztulásában szerepet játszanak pl. a bomló szerves anyagok eltávolítása révén.
- Túlnépesedett populációkban járványokat okoznak, ezzel hozzájárulnak a populációk egyedszámának a szabályozásához.
Egészségügyi jelentőség:
- A parazita szervezetek betegségeket, járványokat
- Az emberi test egészséges működéséhez nélkülözhetetlenek a velünk együtt élő baktériumok, a vastagbélben, a bőr felületén, a hüvelyben.
- Az emberi szervezetben élő mikrobák – szimbióta, kommenzalista, parazita baktériumok, gombák, egyéb mikroorganizmusok – összességét mikrobiomnak nevezzük (részletesen lásd 4.8.5. fejezet).
Ipari jelentőség:
- Iparban hulladékfeldolgozásra, szennyvíztisztításra hasznosítják őket.
- Egyes lebontó baktériumokat olajfoltok megszüntetésére használják.
- Metántermelő baktériumok biogáz (CH4) előállítására képesek.
- Élelmiszergyártásban az emberiség évezredek óta használja az erjesztő baktériumokat (az élesztőgombákkal és penészgombákkal együtt) olyan alapvető élelmiszerek készítésére, mint a bor, sajtok, savanyúság, ecet, szójaszósz, savanyú káposzta vagy joghurt.
- A gyógyszeriparban egyes gyógyszerek, vitaminok – B12 - előállításában szintén baktériumok működnek közre, mint pl. a biotechnológiában alkalmazott baktériumok inzulin, növekedési faktorok előállítására képesek.
Mezőgazdasági szerep:
- A pillangósokat vetésforgóban alkalmazzák, mivel az eljárás megnöveli a talaj nitrogén tartalmát. A vetésforgó a növénytermesztés olyan tervszerű rendszere, amelyben a különféle növényeket térben és időben előre kidolgozott sorrend szerint termesztik.
- Biológiai védekezés során a mezőgazdasági kártevőket – rovarokat, gombákat – elpusztító baktériumokat alkalmaznak védekezésként, mivel az emberre, az élővilágra és a hasznos rovarokra kicsi vagy semmilyen káros hatással nincsenek, környezetbarát rovarirtónak tekinthetőek.
Evolúciós jelentőség:
- Ők voltak az elsőként megjelenő élőlények,
- megkezdték az oxigénlégkör kialakítását,
- eukarióták őseinek tekinthetők.
Az eukarióta sejtek kb. 1,5-2 milliárd évvel ezelőtt jöttek létre. Kialakulásukat ma az ún. endoszimbióta elmélettel magyarázzák. Ennek lényege röviden az, hogy az eukarióta sejt egyes sejtszervecskéi, pl. a zöld színtest, a mitokondrium különféle prokarióta sejtekből alakultak ki. Ez úgy történhetett, hogy egy ősi prokarióta sejt bekebelezett pl. egy kékbaktériumot, melyet nem emésztett meg, hanem tartós szimbiózisra lépett vele, így létrejöttek a színtestek. Bizonyítékok:
- mindkét sejtszervecskének saját genetikai állománya van, amely a prokariótákéhoz hasonló.
- A mitokondriumot és a színtesteket is kettős hártya határolja, közülük
- a legbelső a prokarióták sejtmembránjához hasonló szerkezettel rendelkezik,
- a külső membrán ettől eltérő szerkezetű és szerepű, a bekebelezés során alakulhatott ki.
- Önállóan képesek osztódni.
Az eukarióta sejtek belső membránrendszere, ill. a maghártya a sejthártya betűrődéseiből leváló membránlemezekből alakultak ki.
17. tétel: mohák, harasztok |
Mohák (törzse)
A mohák már szárazföldi növények, ugyanakkor szaporodásuk még vízhez kötött. Az elsőként kialakuló szárazföldi növények, első képviselőik kb. 470 millió éve jelentek meg, a tengerek árapály zónájában élő ősi zöldmoszatokból fejlődtek ki. Azonban fejlődésük megrekedt, a magasabb rendű növények felé semmiféle kapcsolatot nem mutatnak, az evolúció egyik zsákutcáját képviselik. Korábban egységes törzsként tárgyalták őket, a mai filogenetikus rendszerek már 3 külön törzsbe sorolják képviselőiket. A mohák fejlett teleptestes szerveződésűek, a szövetes szerveződési formát nem érték el.
 |
A mohatelep gyakran testszerű, tagoltsága a magasabb rendű növényekére emlékeztet, különböző alakú és működésű sejtekből épül fel, azonban valódi szöveteket nem, legfeljebb kezdetleges szövetelemeket tartalmazhat.
- A növényt a talajhoz gyökérszerű fonalak rögzítik (gyökerecske),
- a fotoszintetizáló leveleket (levelecske)
- szárszerű képlet (száracska) emeli a talajszint fölé.
- Spóráik nyélen ülő spóratartóban fejlődnek.
A vizet és az ásványi anyagokat egész testfelületükön keresztül veszik fel, melyet a szárukban található kezdetleges vízszállító sejtek továbbítanak.
A levélkék általában csupán egyetlen sejtrétegből állnak.
Szaporodásuk többféle módon mehet végbe.
- Vegetatív úton feldarabolódással, mivel a moha száraz állapotban könnyen törik, morzsolódik, s így minden töredékből új növény fejlődhet.
- Ivartalanul spórákkal,
- ivarosan ivarsejtekkel, mely két utóbbi szaporodási forma a mohák egyedfejlődésében is nemzedékváltakozást - kétszakaszos fejlődést - eredményeznek.
Haploid, ivaros szakasz (ahol az ivarsejtek keletkeznek)
1. Egyedfejlődésük a haploid spórából indul ki.
2. A spórából mitózissal fonalas haploid előtelep keletkezik.
3. A fonalas előtelepből mitózissal kifejlődik a haploid mohanövényke.
4. A mohanövényen megjelennek az „ivarszervek”.
5. A kétostoros hímivarsejtek vízcseppben a női ivarszervekhez úsznak, és abban megtermékenyítik a petesejtet (megtermékenyítéshez elengedhetetlen a víz!).
Diploid, ivartalan szakasz (ahol a spórák keletkeznek)
6. A megtermékenyítést követően létrejön a diploid zigóta.
7. A zigótából mitózissal kifejlődik a diploid ivartalan nemzedék, amely csupán egy nyélen függő spóratartó tok.
8. A spóratartó tokban meiózissal keletkeznek a haploid spórák.
A mohák életciklusa során a haploid ivaros életszakasz sokkal fejlettebb a diploid ivartalan szakasznál, hiszen a spórán kívül ebbe az életszakaszba sorolható az előtelep, a mohanövény, az ivarszervek és az ivarsejtek.
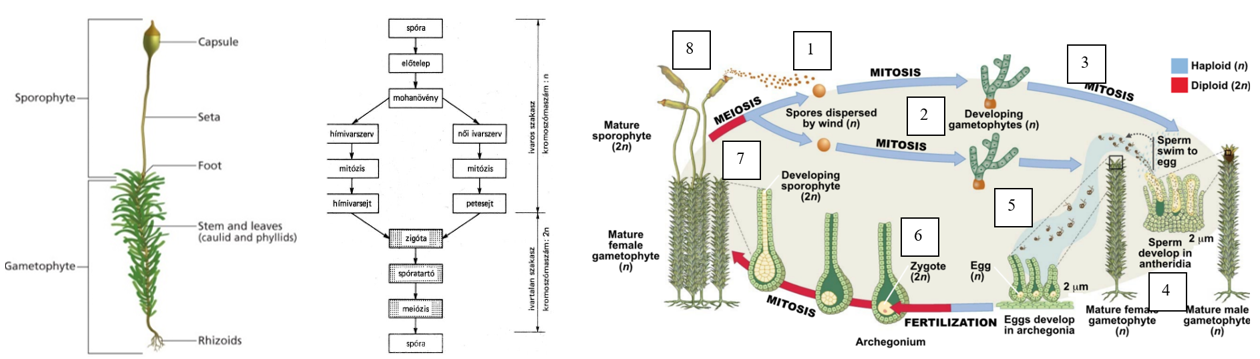 |
Életmód, előfordulás, ökológiai jelentőség
Igen elterjedtek, a sarkvidékektől egészen a trópusokig mindenhol megtalálhatók. A mohák számos olyan élőhelyet benépesítettek, ahol a vízellátás csupán rövid ideig kedvező. Légszáraz állapotban a mohák hetekig, sőt hónapokig, egyes fajok évekig megtartják életképességüket. Ebben az állapotban a plazmájuk besűrűsödik, életfolyamataik rendkívül lelassulnak. Ha vízhez jutnak életfolyamataik pillanatok alatt helyreállnak, mivel a mohák levélkéit nem borítja kutikula, így a mohalevélke teljesen átjárható a víz és a benne oldott sók számára, s e tulajdonság révén a növény a vizet és az ásványi anyagokat a teljes testfelületén képes felvenni, mégpedig igen gyorsan. Ugyanakkor a növény vízmegtartó képessége kicsi – változó vízállapotú növények. A mohák ezt úgy ellensúlyozzák, hogy szorosan, tömött párnácskákban nőnek.
A természetes társulásokban – pl. a trópusokon - igen fontos szerepet töltenek be azáltal, hogy a hirtelen lehulló nagy mennyiségű csapadékot megkötik és fokozatosan juttatják vissza a környezetbe. Így meggátolják a talajeróziót (a csapadékvíz általi talaj lemosódást), élőhelyükön kiegyenlített nedvességviszonyokat biztosítanak (ökológiai szerep).
Igénytelenségük miatt fontos szerepük van a termőtalaj kialakításában, ugyanis elsőként jelennek meg olyan helyeken, ahol más élőlények még nem fordulnak elő, ezért a mohákat pionír élőlények közé soroljuk (ökológiai szerep).
Egyes fajok egyedei bizonyos nehézfém-ionokat szelektíven megkötnek, testükben felhalmoznak, ezért alkalmasak a levegőszennyezettség kimutatására (indikátor fajok, ólmot kötnek meg), ill. pl. réz vagy nikkel lelőhelyek felkutatására.
Harasztok (törzse)
A harasztok kb. 400 millió évvel ezelőtt való megjelenése az evolúció történetében merőben új fejezetet jelentett, amennyiben a harasztok voltak az első valódi szövetes és az első valódi hajtásos – valódi szervekkel rendelkező – növények (a hajtás a leveles szár).
A mohákhoz hasonlóan szintén ősi zöldmoszatokból alakultak ki a földtörténeti ókorban, kb. 400 millió évvel ezelőtt. A valódi szövetek, a növényi szervek kialakulását a szárazföldi életmódra való áttérés indukálta.
Amíg a növény vízben élt
- nem volt szüksége gyökérre, hiszen benne élt a tápoldatban, a tápanyagok felszívását egész testfelületen végezte, ennek következtében
- szállítószöveteket sem tartalmazott,
- nem volt kitéve a kiszáradás veszélyének sem, így a bőrszövetek kialakulása sem következett be,
- mechanikai szövetek sem alakultak ki, hiszen nem kellett a növényi testet tartani.
 |
A harasztok felépítésének rövid jellemzése
- Valódi szerveik, szöveteik vannak,
- állandó vízállapotúak, azaz már képesek a vízforgalmukat – leadás, felvétel – szabályozni, vízháztartásukat bizonyos határok között függetleníteni a környezetüktől.
- Lágyszárúak (kivéve páfrányfák).
- Leveleik
- általában kicsik, pikkelyszerűek, pl. korpafüvek, zsurlók
- de lehetnek nagy felületűek, pl. páfrányok.
- Virágaik nincsenek, a spórák vagy
- külön spóratermő füzéreken, korpafüvek, vagy
- a levelek fonákján jönnek létre, pl. páfrányok.
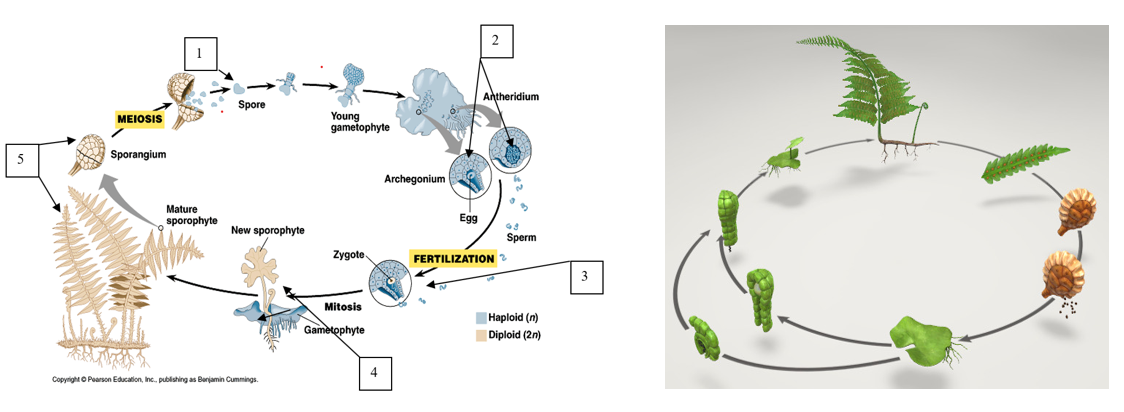 |
A harasztok szaporodása
A harasztok szaporodása több ponton eltér a mohák szaporodásától.
- 1. A harasztok egyes csoportjaiban – páfrányok - a spórák mind alakilag, mind ivarilag eltérhetnek egymástól,
- a hím jellegű spórák a mikrospórák, belőlük mikroelőtelep fejlődik, melyen megjelennek a hím ivarszervek,
- a női jellegű spórák a makrospórák, belőlük makroelőtelep alakul ki, ahol megjelennek a női ivarszervek.
- 2. A harasztnövény már nem a haploid ivaros nemzedékhez (mint ahogyan a mohanövény) hanem a diploid ivartalan szakaszhoz tartozik. Tehát a harasztoknál az ivartalan szakasz fejlettebb az ivarosnál, melyet a spórán kívül már csak az előtelep, az ivarszervek és az ivarsejtek képviselnek. A harasztoktól kezdve az ivaros szakasz egyre redukáltabb felépítésű, míg az ivartalan nemzedék maga a jól fejlett növényi test lesz.
- A harasztnövény kromoszómakészlete tehát diploid.
A harasztok nemzedékváltakozása
Az erdei pajzsika kétszakaszos fejlődésmenete.
Ivaros szakasz
1. A haploid spórából lapos, kb. 0,5 cm átmérőjű, szív alakú, zöld, fotoszintetizáló haploid előtelep fejlődik.
2. Az előtelep fonákján jelennek meg a női és a hímivarszervek.
3. A csillós hímivarsejtek vízcseppben úszva jutnak el a női ivarszervben található petesejthez, s azzal egyesülve létrehozzák a zigótát. Tehát a megtermékenyítéshez a víz, mint közeg még elengedhetetlen.
Ivartalan szakasz
4. A diploid zigóta osztódik, s létrejön a diploid csíranövény, melyből rövidesen kialakul a kifejlett haraszt, gyöktörzzsel, szárral, levéllel (valódi növényi szervek).
5. A kifejlődött diploid páfránynövény leveleinek fonákján kialakulnak a spóratartók, bennük meiózissal keletkeznek a haploid spórák.
A harasztok szaporodása nagymértékben vízhez kötött, egyrészt mivel az előtelep nem bírja a szárazságot, másrészt a megtermékenyítés folyamata sem nélkülözheti a vizet.
18. tétel: Növényi szövetek, folyadékáramlás növényekben |
A növényi szövetek
Az azonos működésű, alakú, eredetű sejtek összességét szövetnek nevezzük, ahol a különféle sejtek különböző feladatokra specializálódnak.
Alapvetően két fő szövettípus különböztethető meg:
- osztódószövetek – merisztémák -, melyek osztódásra képes sejtekből állnak:
- csúcsmerisztéma,
- oldalmerisztéma – kambium,
- közbeiktatott merisztéma.
- állandósult szövetek, melyek differenciált, állandósult sejtekből állnak, működés szerint feloszthatók:
- bőrszövet
- szállítószövet
- alapszövetek
- valódi alapszövetek
- táplálékkészítő
- raktározó
- víztartó
- átszellőztető
- szilárdító
- kiválasztó és váladéktartó alapszövetek.
- valódi alapszövetek
A szövetek kialakulása a szárazföldi életmódhoz való alkalmazkodás következménye. A fejlett hajtásos növényekben az egyszerű szövetek szövetrendszerekbe csoportosulnak, ahol eredeti funkciójukat összehangolva, egymást kiegészítve végzik, pl. a bőr-, szállító-, alapszövetrendszer.
 |
Osztódószövetek
A soksejtű növény is egyetlen sejtből, a megtermékenyített petesejtből fejlődik. A fejlődő embrió eleinte még teljes egészében osztódó sejtekből áll, ám hamarosan az osztódások a növénynek csak bizonyos helyeire korlátozódnak, ahol a sejtek osztódóképességüket a növényi élet egész tartamára megőrzik. Ezek az osztódó sejtek az ún. osztódószöveteket alkotják.
Az osztódószöveteknek köszönhetően képes a növény arra, hogy teste egész élete alatt növekedjen, szemben az állati szervezettel, s ezért hívjuk növekvő lénynek, vagyis növénynek.
Az osztódószövet sejtjeire jellemző, hogy:
- kicsik,
- zöld színtestet, vakuólumot nem tartalmaznak,
- vékony falúak,
- relatíve nagy magjuk és telt plazmájuk
Elhelyezkedésük szerint ismertek:
- csúcsi osztódószövetek a tengelycsúcsokon (hajtáscsúcs, gyökércsúcs) találhatók, melyek a növény hosszirányú növekedéséért felelősek.
- A növényi szerv oldalával párhuzamosan kialakuló osztódószövetek, melyek a növényi szerv vastagodásáért felelősek, pl. kambium.
- Közbeiktatott merisztémák
- a szártagok megnyúlását,
- levélnyél, levéllemez növekedését biztosítják.
 |
A bőrszövetrendszer
A bőrszövetrendszer a hajtásos növények testének felületén levő sejtcsoportok összessége.
Alapvető működési eltérések következtében két típusa van:
- a hajtás és
- a gyökér bőrszövetrendszere.
A fiatal hajtás bőrszövete - epidermis
A fiatal hajtás elsődleges bőrszövete. Funkciója:
- mechanikai védelem,
- kiszáradás elleni védelem,
- a párologtatás, gázcsere lebonyolítása.
A fiatal hajtás bőrszövete általában egy sejtrétegű. A sejtek igen szorosan kapcsolódnak egymáshoz. A folytonosságot csak a gázcserenyílások légrései szakítják meg. Ezek a nyílások, amelyeket két zárósejt vesz körül, nyithatók vagy zárhatók.
A gázcserenyílások bonyolítják le a párologtatást és a gázcserét a külvilág és a növény belső szövetei között.
A bőrszöveti sejtekben – kivéve a gázcserenyílások zárósejtjeit - nincsenek zöld színtestek, így az színtelen. A levél színét a mélyebben lévő szövetek színtestjeitől kapja.
A bőrszöveti sejtek felszínén összefüggő, vízhatlan kémiai védőréteg helyezkedik el, ez a kutikula, amely védi a növényt a kiszáradástól.
A levelek, gyümölcsök epidermiszén gyakran viasz is felhalmozódhat, mely többé-kevésbé összefüggő réteget alkot, pl. ezüstfenyő levél fonákján, almán, szőlőn, szilván.
A növényi szőr
A növényi szőrök a hajtás bőrszövetének képződményei, lehetnek egysejtűek vagy többsejtűek.
Működésük szerint lehetnek:
- fedőszőrök: túlzott párologtatás és a hideg ellen védenek, tőlük a növényi részek puha, selymes tapintásúak, pl. kökörcsin,
- csalánszőrök: a szőr kiemelkedő, csúcsi részén egy gömb alakú fejecske található, mely érintésre letörik, s a merev szőrcsonk, mint valami injekcióstű beszúródik a bőrbe, és a szőrsejtben lévő acetilkolint, hisztamint stb. bejuttatja a bőr szöveteibe gyulladást okozva,
- repítőszőrök: pl. gyapot magvain.
 |
A fiatal gyökér bőrszövete - rizodermisz
A fiatal gyökér elsődleges bőrszövete. A gyökér bőrszövetén nincsenek gázcserenyílások és nincsen kutikularéteg sem. A gyökér bőrszövetének zárvatermőkre jellemző speciális képződményei a gyökérszőrök. A gyökérszőrök a gyökér bőrszövet sejtjeinek kesztyűujjszerű kitűrődései. A gyökérszőrökön keresztül történik a víz és az oldott ionok felvétele.
A gyökérszőrök feladata a felületnagyobbítás. A gyökérszőrök élettartama néhány nap, a csúcstól távolabb pusztulnak, a csúcs irányába mindig megújulnak.
Másodlagos bőrszövetek
Periderma
A vastagodó hajtás és gyökér felületén, az elszakadozó elsődleges bőrszövet helyett kialakul egy több sejtrétegű, igen ellenálló, elhalt bőrszövet. Általában ismeretes a néhány éves fiatal fásszárú növények szárán, gyökerén.
Héjkéreg
A fás növények harmadlagos bőrszövetrendszere. A másodlagos bőrszövetrendszert (peridermát) váltja fel az évről évre vastagodó fás szárban. Képződése során a héjkéregbe elhalt háncselemek is belekerülnek. A vastagodó szár belső térfogat gyarapodása következtében a héjkéreg gyakran repedezett (pl. tölgy).
 |
A szállítószövet-rendszer
A kezdetlegesebb szerveződési szinten álló telepes növények testében a tápanyagok többnyire sejtről sejtre vándorolnak diffúzióval.
A hajtásos növények testében a gyökerek által felvett víznek el kell jutnia a levelekbe, ill. a test minden élő sejtjébe. Ugyanakkor a levelekben keletkezett szerves termékeknek is el kell jutniuk a raktározó szövetekhez. Ezt az összetett feladatot a szállítószövet-rendszer végzi el.
A szállítószövet-rendszer sejtjei
- a legkisebb ellenállást nyújtva a szállítás irányába megnyúltak,
- végfalaik ferdén állnak (nagyobb tapadási felület).
- A végfalak lyukacsosak, esetleg teljesen felszívódnak.
A kettős funkciót ellátó szövetrendszer két részre osztható:
- a vizet és sókat szállító farészre (xylem),
- a szerves anyagokat szállító háncsrészre (phloem).
 |
A farész
- Harasztokban és nyitvatermőkben elhalt sejtek (tracheidák) szállítják a vizet és a sókat a gyökér felől a hajtás felé.
- Zárvatermőkben a sejtek válaszfalainak felszívódásával vízszállító csövek alakultak ki, melyek oldalsó falai spirálisan megvastagodnak, védve a csöveket az összenyomódástól.
- A csövek hossza néhány cm-től 8-10 méterig terjedhet. A csövek egy vagy több éven át működőképesek, azután gyanta és gumiszerű anyagok eltömik őket.
- A faparenchima sejtek a farész élő plazmatartalmú sejtjei, amelyek főképpen a tápanyag-raktározásában vesznek részt.
- A farostok hosszúra nyúlt, kihegyezett végű, vastag, fásodott falú, elhalt sejtek. Támasztják, szilárdítják a vizet szállító faelemeket.
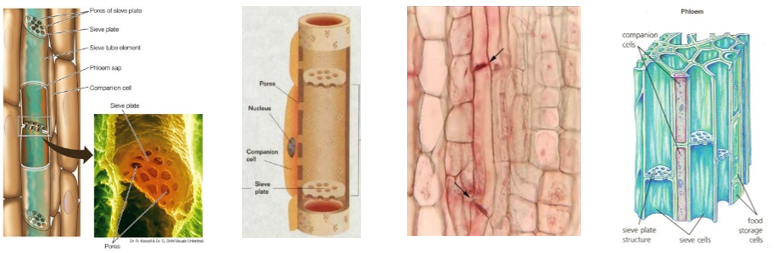 |
A háncs
- A háncsrész szerves anyagokat szállít a növényi test minden irányába.
- A harasztok és a nyitvatermők háncsrészének a szállítóelemei élő rostasejtek,
- zárvatermőkben több sejtnek, ún. rostacsőtagnak az egyesüléséből rostacsövek jönnek létre. A rostasejteket rostalemezek választják el.
- Kísérősejtek: a zárvatermők rostacsőtagjait kísérik. Plazma- és magtartalmú élő sejtek, a tápanyagszállítást segítik.
- Háncsparenchima sejtek: élő, plazmatartalmú, tápanyagokat raktározó sejtek.
- Háncsrostok: farostokhoz hasonló alakú, de cellulóz falú szilárdító elemek. Háncsrostok közé tartoznak az iparilag hasznosított rostok, pl. len, kender stb.
 |
A szállítóelemek elrendeződése alapján két fő szártípust különböztetünk meg:
- Zárt kambium-henger alakul ki, amely kifelé összefüggő háncs hengerpalástot, befelé összefüggő fahengert fejleszt. Ez a fáinkra jellemző fás szár (évgyűrűs szerkezet).
- Másik esetben a kambium elkülönült kötegekre, egységekre bomlik, s ezek különálló, ún. szállítónyalábokat fejlesztenek. Ez a típus a lágyszárúakban ismert.
Maguk a nyalábok lehetnek
- egyszerűek: ha csak fa- vagy csak háncselemekből állnak, pl. fiatal gyökerekben.
- a szárban összetettek, ahol egy nyalábban belül fa- és kívül háncselemeket egyaránt találunk.
- Nyílt nyalábnak nevezzük, ha a fa- és a háncsrész között működő kambium van, pl. kétszikű lágyszárú növények szárában.
- Zárt nyalábról beszélünk, ha a kambium a fa- és a háncsrész kifejlesztése után eltűnik. Ez a típus található a levelekben, ill. az egyszikűek szárára jellemző (már nem képesek másodlagos vastagodásra).
 |
Az alapszövetrendszer
A növényi testben mindaz, ami nem bőr-, és nem szállítószövet, az alapszövet. Működésük alapján megkülönböztetünk:
- valódi alapszöveteket,
- szilárdító alapszöveteket,
- kiválasztó (szekréciós) alapszöveteket.
Valódi alapszövetek
Élettani működésük alapján a valódi alapszöveteket további típusokra oszthatjuk.
a) Táplálékkészítő alapszövetek
A fotoszintetikus szervesanyag-építés színhelyei, a nagy mennyiségű zöld színtesttől zöldek. Előfordulnak a levelek középső részében, fiatal zöld szárakban, termésekben, csészelevelekben (mindenhol, ahol a növény zöld). Sejtjeik lazán helyezkednek el, vékony falúak, oldatok és gázok számára könnyen átjárhatók.
b) Raktározó alapszövetek
A sejtekben különböző tápanyagokat raktároznak, vízben oldhatatlan, ún. zárványok formájában: keményítőt, olajat, fehérjéket. Elsősorban a fénytől elzárt növényrészekben találhatók meg: gyökerekben, magvakban, gumókban.
Ilyenek pl. a fák faparenchima sejtjei. A faparenchima sejtekben raktározott tápanyagot a növény rügyfakadáskor használja fel, s így ezek évről évre újra raktározzák a tápanyagokat.
c) Víztartó alapszövetek
A száraz környezetben élő pozsgás növények testében valóságos víztartószövetek alakultak ki. A sejtekben nagy központi vakuólum van, amely szinte teljesen kitölti a sejtet. A vakuólumban vizes nyálka formájában raktározódik a víz. Kialakulhat a szárban (kaktuszok), vagy a levélben (kövirózsa).
d) Levegőtartó alapszövetek
Levegővel teli sejtközötti járatokban gazdag alapszövet, ahol a járatok sokszor jelentékeny üregekké tágulnak, valóságos légkamrarendszert alkotnak. Tipikusan vízinövények víz alá merült részeiben: levélnyelekben, szárakban, gyökerekben figyelhető meg.
 |
Szilárdító (mechanikai) alapszövetek
A növényi test méretének növekedése következtében, a lombozat megtartásához komolyabb szilárdításra van szükség, s ezt a szerepet a mechanikai szövetek töltik be.
A szilárdítószövetekben a sejtfalakra jellemző a sejtfalvastagodás, gyakran a sejtfalba faanyag, ún. lignin rakódik, amely a növényi részeknek szilárdságot, tartást biztosít. Ide tartoznak a farostok és a háncsrostok is.
Kiválasztó és váladéktartó alapszövetek
A növényeknek az állatok veséjéhez hasonló elkülönült kiválasztó szervük nincs. Ugyanakkor a növényi anyagcsere során is keletkeznek káros, felesleges anyagok melyek
- vagy oldhatatlan zárványok formájában a sejtekben elkülönülnek, felhalmozódnak (pl. Ca-oxalát),
- vagy a növényi szervezetből valamilyen módon kiürülnek.
Típusok
a) Egysejtű vagy többsejtű mirigyszőrök a bőrszöveten.
b) Nektáriumok, amelyek a nektárt választják ki, ami a viráglátogató rovarok egyik fő tápláléka. A kiválasztott cukor a háncsrész szállítóelemeiből származik.
c) Ozmofórák, amelyek illatkiválasztó képződmények. A virágillat szintén a rovarok csalogatásában játszik szerepet.
d) Gyantajáratok. Ilyenek vannak pl. a fenyőfélék testében. A megkövesedett gyanta a borostyán.
e) Tejcsövek. A tejcsövek összeolvadó sejtsorok, amelyek tejszerűen folyékony tejtnedvet tartalmaznak. A tejnedv a tejcső sejtnedve.
- A kaucsukfa tejnedve a kaucsuk, korábban a gumi gyártásának volt az alapanyaga.
- A mák tejnedve sok alkaloidot tartalmaz, a mák kicsorgó és megszáradó tejnedve az ópium.
 |
Folyadékáramlás növényekben
A víz és a sók felvételének helye
A gyökerek felszívási zónájának gyökérszőrei.
A víz és a sók felvételének módja
A gyökérszőrök sejthártyája féligáteresztő sajátosságú, azon a víz passzívan, ozmózissal beáramolhat, mivel a gyökérszőr sejtjeinek a citoplazmája töményebb, mint a talajoldat, így sejtplazma egyfajta szívóerőt gyakorol a gyökérsejtek körüli vízmolekulákra. A beáramlott víz növeli a sejten belüli nyomást, turgor állapotot létrehozva, ezért a víz átpréselődik a szomszédos alapszöveti sejtekbe, majd onnan a farész vízszállító csöveibe (gyökérnyomást kialakítva).
A szükséges ionokat a gyökérszőrök sejthártyája válogatja ki a növény szükségleteinek megfelelően, majd a koncentrációviszonyoktól függően (esetleg passzív, de) főleg aktív transzporttal a sejt belsejébe juttatja. Az energiaigényes transzporttal az ionok a nagyobb koncentráció irányába is szállítódhatnak.
A bőrszövettől a szállítószövetek farészéig a sók transzportja révén egy egyre növekvő koncentrációgradiens biztosítja a víz mozgását az ozmózis szabályainak megfelelően.
Tehát a gyökérben a turgornyomás és a szomszédos sejtek egyre növekvő szívóereje a vizet bepréseli a farészbe, majd onnan a szárba, amelynek eredményeképpen egy ún. gyökérnyomás alakul ki.
A gyökérnyomás tehát az ozmózis hatására létrejövő a hajtóerő, amely a vizet a szállítónyalábokba préseli. Fákban néhány méter magasságig képes a vizet felnyomni.
Még egyszer:
- az ionok aktív transzporttal jutnak be a gyökérszőrsejtbe, illetve adódnak tovább sejtről sejtre egészen a farészig,
- a víz az ionok vándorlását passzívan követi.
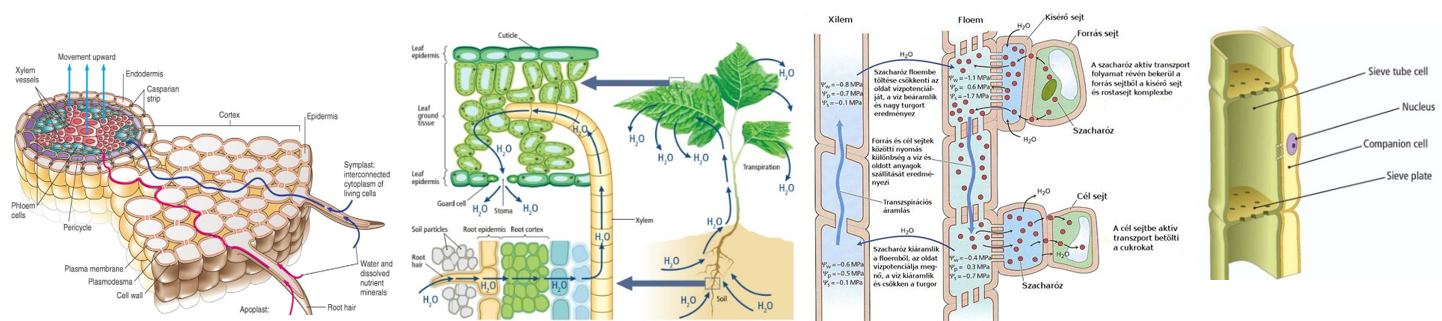 |
A víz és az ásványi anyagok szállítása
A vizet és az ásványi sókat a növények a gyökerek felszívási zónájában, a gyökérszőrökön keresztül veszik fel. A sók általában aktív transzporttal, a víz passzív transzporttal (ozmózis) jut be a gyökérszőrök sejtjeibe.
A növényi szervezetben a gyökerektől a levelekig a szomszédos sejtek szívóereje által meghatározott vízút alakul ki.
Ennek legfőbb kialakítói:
- a gyökérnyomás
- és a párologtatásból eredő szívóerő.
A párologtatásból származó szívóerő
A gyökérnyomás csupán néhány méterre képes a vizet felnyomni, így önmagában nem elegendő a fák folyamatos vízáramlásának fenntartásához.
A vízáramlás felső mozgatója a párologtatásból eredő szívóerő.
A szívóerő a levelekben keletkezik a párologtatás nyomán, melynek során a növények a vizet gőz formájában adják le a környezetükbe. A vízvesztést a levélszövetek sejtjeinek koncentráció növekedése kíséri, egyben azok ozmotikus szívóerejét növeli. A sejtek az elvesztett vizet a levélerek farészéből pótolják. A szívóerő annál nagyobb, minél több vizet veszít a sejt, azaz minél intenzívebb a párologtatás.
A párologtatás főleg a levelek gázcserenyílásain keresztül valósul meg, melynek mértékét a növény a sztómák nyitásával-záródásával szabályozni képes.
A vízmozgás szempontjából rendkívül fontos, hogy a vízoszlop folytonos legyen, sehol ne szakadjon meg. Ezt biztosítja
- a vízmolekulákat összetartó kohéziós erő, ill.
- a vízoszlop és a szállítóedény fala közti tapadás, az adhézió, a kapilláris hatás.
A szerves anyagok szállítása
A szállítószövetek háncsrészében történik, iránya szerteágazó, a levelekből főleg a raktározó szövetekhez, ill. az osztódószövetekhez.
A szerves anyag, amely szacharóz, sejtről sejtre aktív transzporttal adódik tovább, aminek aktív segítői zárvatermőkben a rostacsövek melletti kísérősejtek. A szerves anyagok szállításának nincsen kitüntetett iránya, nyáron, éjszaka a levelek felől a gyökerek raktározó alapszövetei felé, tavasszal fordítva, a gyökerek felől a képződő rügyek irányába.
A farész és a háncsrész anyagforgalma bizonyos helyeken összekapcsolódik,
- a levelekben, mivel a rostacsövekben nagyobb az oldatok ozmózisnyomása, a farész felől víz áramlik a háncsrész felé, növelve a háncselemek turgornyomását.
- A gyökérben fordított a helyzet, mivel a raktározó alapszövetek a kísérősejtek segítségével felveszik a szerves anyagokat, a rostacsövekben csökken az ozmózisnyomás, így a háncsrészből víz áramlik a farész felé.
Tehát Münch 1926-ban leírt nyomás-áramlási modellje szerint a forrás és a célállomás közötti eltérő ozmózisnyomás alapján működik a szállítás.
Eszerint a
- a levelek fotoszintézist végző sejtjeiből, aktív transzport szállítja a cukrot a rostacsövekbe,
- ahol emiatt nő az ozmózisnyomás.
- Ennek következtében a vízszállító csövekből víz áramlik a rostacsövekbe, ahol a beáramló víz miatt növekszik a turgornyomás.
- A felhasználás helyén, például a raktározó alapszövetekben, a rostacsövekből szállítja aktív transzport a cukrot a környező sejtekbe,
- ami miatt csökken a rostacsövekben az oldat ozmotikus nyomása.
- Ennek hatására a víz kiáramlik a rostacsövekből, ami miatt csökken a turgornyomás. A rostacsövekben tehát az oldat a nagyobb nyomású hely felől a kisebb nyomású hely felé áramlik.
A növények anyagszállításának vizsgálata
- Muskátli levágott szárát állítsuk piros tintával megfestett vízbe.
- Majd a végétől bizonyos távolságra vágjuk el, és készítsünk keresztmetszetet a szárból!
- Tegyük tárgylemezre, fedjük le, és vizsgáljuk mikroszkóppal!
19. tétel: Evolúciós újítások hajtásos növényekben |
Az első többsejtű növények moszatok (algák) voltak, melyek a korai óidő tengereiben jelenhettek meg. A magasabb rendű szárazföldi növények kialakulásának szempontjából az algák közül a legfontosabbak az ősi zöldmoszatok voltak. A szárazföldön egészen addig nem jelenhetett meg az élet, amíg az oxigénszint el nem érte kb. a mai értéket, aminek köszönhetően az ózonréteg vastagodása miatt a halálos UV sugarak elnyelése olyan mértékű lett, hogy a szárazföldön megjelenő élőlényeket már nem károsította. Ez nagyjából a szilur időszak végén, kb. 400 millió évvel ezelőtt következett be, s így a tengerek árapály zónájában élő ősi zöldmoszatokból kialakulhattak egymással párhuzamosan a mohák és a harasztok. Ezzel kezdetét vette a szárazföld meghódítása.
A mohák fejlett teleptestes szerveződésűek, a szövetes szerveződési formát nem érték el. Az evolúció egyik zsákutcáját képviselik, fejlődésük megrekedt, a magasabb rendű növények felé semmiféle kapcsolatot nem mutatnak.
A harasztok megjelenése az evolúció történetében merőben új fejezetet jelentett, amennyiben a harasztok voltak az első valódi szövetes és az első valódi hajtásos – valódi szervekkel rendelkező – növények (a hajtás a leveles szár).
A harasztok felépítésének rövid jellemzése
- Valódi szerveik, szöveteik vannak,
- állandó vízállapotúak, azaz már képesek a vízforgalmukat – leadás, felvétel – szabályozni, vízháztartásukat bizonyos határok között függetleníteni a környezetüktől.
- Lágyszárúak (kivéve páfrányfák).
- Leveleik
- általában kicsik, pikkelyszerűek, pl. korpafüvek, zsurlók
- de lehetnek nagyfelületűek, pl. páfrányok.
- Virágaik nincsenek, a spórák vagy
- külön spóratermő füzéreken, korpafüvek, vagy
- a levelek fonákján jönnek létre, pl. páfrányok.
A valódi szövetek, a növényi szervek kialakulását a szárazföldi életmódra való áttérés indukálta.
Amíg a növény vízben élt
- nem volt szüksége gyökérre, hiszen benne élt a tápoldatban, a tápanyagok felszívását egész testfelületen végezte, ennek következtében
- szállítószöveteket sem tartalmazott,
- nem volt kitéve a kiszáradás veszélyének sem, így a bőrszövetek kialakulása sem következett be,
- mechanikai szövetek sem alakultak ki, hiszen nem kellett a növényi testet tartani.
A harasztok első képviselői virágkorukat a karbonban élték, hiszen az akkori meleg, csapadékos klíma kedvezett elterjedésüknek (hiszen szaporodásukhoz víz kell), majd e korszak vége felé, a Föld klímájának szárazabbra fordulása miatt hanyatlásnak indultak, fajaik többsége kihalt.
A harasztok ősi képviselői nagy termetű, fás szárú növények voltak, törzsük átmérője a 2 m-t, magasságuk a 30 m-t is elérte. Óriási mocsárerdőket alkottak. Ide tartoztak a pikkelyfák és a pecsétfák, melyek nevüket a levelek lehullása után a törzsön visszamaradó jellegzetes alakú levélcsonkokról kapták. E növények maradványaiból alakultak ki a mai kőszéntelepek anaerob bomlás eredményeképpen, továbbá óriási jelentőségük, hogy ugrásszerűen megnövelték a légkör oxigéntartalmát.
 |
A következő nagy evolúciós ugrás a virág és a mag megjelenése volt, aminek köszönhetően a megtermékenyítéshez már nem volt szükség vízre. A földtörténet során elsőként megjelenő virágos, magvas növények a nyitvatermők voltak, melyek kb. 300 millió évvel ezelőtt alakultak ki ősi harasztokból.
A virágpor szél útján közvetlenül a petesejtet tartalmazó magkezdeményre került, ilyen módon az ivarsejtek egymás mellett alakultak ki, így már a megtermékenyítéshez nem volt szükség vízre.
Mivel a szaporodásuk függetlenné vált a víztől, továbbá viaszos leveleik miatt kiválóan szabályozták vízháztartásukat, így a növények közül elsőként hódították meg a szárazföldeket. Mivel jól bírták a vízhiányt, a földtörténeti középkor meleg, száraz klímája kedvezett elterjedésüknek.
A nyitvatermők főbb jellemzői
- Kivétel nélkül fás szárúak.
- Többségük örökzöld, néhány fajuk lombhullató vörösfenyő, Ginkgo biloba, leveleik főleg tű vagy pikkely alakúak, ritkán nagy felületűek (Ginkgo biloba).
- Virágaik egyivarúak, általában egylakiak.
- Egyivarú virág: vagy csak termő vagy csak porzó található a virágban.
- Egylaki növény: egy egyeden mind a termős, mind a porzós virágok megtalálhatók.
- Virágtakaró levelek nincsenek, így a virágok felépítése igen egyszerű.
- A porzós virágzat általában barkaszerű, főleg az ágak csúcsán találhatók meg, bennük porzók kicsik, pikkelyszerűek.
- A termős virágok termőlevelei lemezesek, melyek füzérei a tobozvirágzatot alkotják.
- A törzs nevét onnan kapta, hogy a magkezdemények – szám szerint kettő - a termőlevelek tövében szabadon fejlődnek (termő, zárt magház, termés nincs).
- A keletkezett virágpor a szél közvetítésével közvetlenül a magkezdeményre jut. A megtermékenyítést követően a magvak szárnyas repítőszerkezetük segítségével, a toboz felnyílása után nagyobb távolságot is megtehetnek.
A ma élő legfejlettebb növények a zárvatermők, ősi nyitvatermőkből jöttek létre. Első képviselőik a krétakorban jelentek meg, kb. 120 millió éve. A törzs a nevét a zárt termőről kapta.
A zárvatermők főbb jellemzői
- Kialakulásuk a magvaspáfrányokból (ősi nyitvatermők) indult.
- A törzs a nevét a termőlevelek összezáródásából kialakuló zárt termőről kapta.
- A termő alsó része a magház, amely a bibeszálban, ill. a bibében folytatódik.
- A magházban a megtermékenyítést követően a magkezdeményekből fejlődnek a magok.
- Jellemző a kettős megtermékenyítés (később).
- A termő falából termés képződik.
- Megjelenik a színes virágtakaró (rovarbeporzás).
- Rovarmegporzás jellemző (de lehet szél- és vízbeporzás is).
- Szállítócsövek, gyökérszőrök alakultak ki.
- Két fő csoportjuk (osztályuk) van: egyszikűek és a kétszikűek.
A zárvatermők kettős megtermékenyítése, a mag kialakulása
- A különböző módon szállított – szél, ill. rovar - virágpor végül a ragadós felületű bibén köt ki (1).
- Itt a kétsejtes pollen tömlőt hajt és behatol a bibe szövetébe. A tömlő képzésében a vegetatív sejtnek van jelentős szerepe (2).
- A fejlődő tömlőben a haploid generatív sejt mitózissal kettéosztódik, s létrejön két szintén haploid hímivarsejt (tehát a hímivarsejtek nem a porzóban, hanem a termőben jönnek létre).
- A pollentömlő a termőlevél szövetében lenő egészen a magkezdeményig, majd behatol az embriózsákba, ahol a tömlő felnyílásával az egyik hímivarsejt a petesejttel, a másik hímivarsejt a központi sejttel egyesül (3).
A megtermékenyítést követően a magkezdeményből kialakul a mag, mely során
- a petesejtből zigóta, majd abból sokszoros mitózissal csíra vagy embrió fejlődik,
- a központi sejtből a mag triploid táplálószövete,
- a magkezdeményt borító burokból a maghéj alakul ki (összenőhet a termésfallal, pl. a szemtermésben).
A zárvatermőknél megjelenő evolúciós újítások: virágtakarólevelek, termő, zárt magház, termés, szállítócsövek, gyökérszőrök tovább tökéletesítették a szárazföldi életmódhoz történő alkalmazkodást.
20. tétel: Kétéltűek, hüllők, madarak |
Négylábúak
A szárazföldi négylábú gerincesek törzsfejlődése az ősi bojtosúszójú halak ágától vált le, kb. 350 millió évvel ezelőtt. A kövületek alapján az ősi bojtosúszójúak páros úszóinak belső, csontos felépítése megegyezik a magasabb rendű gerincesek végtagvázának alapszabásával, ezért a halaktól a kétéltűekhez vezető átmeneti formának tartják őket. Páros úszóik a szárazföldi ötujjú végtagtípus előfutárai, mert minden irányban izmokkal mozgatható, belső csontos vázzal rendelkeztek.
A ma élő bojtosúszójúak osztályába tartozó maradványhalak, ennek az ősi csoportnak ma is élő leszármazottjainak tekinthetők, élő kövületek.
 |
Kétéltűek osztálya
A szárazföldi életmódra való áttérés szükségessé tette:
- a szárazföldön való mozgáshoz az ötujjú végtagtípus kialakulását,
- a kiszáradás ellen a bőr hámrétegének elszarusodását,
- a légköri levegőből a gázcsere lebonyolítását végző tüdő kialakulását.
Virágkorukat a karbon és a perm időszak során élték (350-250 millió évvel ezelőtt), a mai fajok széles elterjedésűek, de hiányoznak a pólusok közeléből és az óceáni szigetekről.
A kétéltűek a két világ – víz és szárazföld - határán élnek, idejük legnagyobb részét a szárazföldön töltik, ugyanakkor testfelépítésük és főleg szaporodásuk miatt életük vízhez, nedvességhez kötött.
Főbb jellemzőik
- Szárazföldi életmódhoz részben alkalmazkodott, állkapcsos gerincesek.
- Változó testhőmérsékletű állatok, a kedvezőtlen időszakokat – szárazságot vagy hideget -nyári álommal vagy téli hibernációval vészelik át.
- Kialakult az ötujjú végtagtípus.
- 2 pár, szárazföldi közlekedésre alkalmas, a törzshöz általában oldalról kapcsolódó tolólábuk van, amely tartósan nem emeli a testet a talaj fölé.
- Kivétel nélkül ragadozók.
- Tüdővel lélegeznek, ugyanakkor jelentős a bőrlégzésük.
- 2 vérkörük, 3 üregű szívük van.
- A szaporodásuk vízhez kötött, a kifejlett egyedek pedig szárazföldön, de többnyire nedves környezetben élnek.
- Átalakulással fejlődnek.
Vízhez kötött életmódjuk oka, hogy
- bőrük még nem véd tartósan a kiszáradástól,
- szaporodásuk vízhez kötött.
 |
Vázrendszer, mozgás
- Nincs csontos szemüregük.
- Koponyájuk a halakéval szemben mozgathatóan kapcsolódik a nyakcsigolyákhoz, azonban a békák fejüket nem tudják mozgatni, nyakuk nincs.
- Kevés csigolyájuk van.
- Bordáik csökevényesek, mellkasuk nincsen.
- Ötujjú végtagjuk tolóláb, békáknál a hátsó láb ugróláb.
- Alkarjuk, lábszáruk egyetlen csont.
Kültakaró
Nyálkás bőrük mirigyekben gazdag. Kissé véd a kiszáradástól, de nem akadályozza a gázcserét.
A felhám gyengén elszarusodó többrétegű laphám.
- Véd a mechanikai hatások ellen.
- Nem véd a kiszáradástól.
Az irha vérerekkel gazdagon átszőtt (bőrlégzés), kötőszövetes réteg, amely tartalmaz:
- nyálkamirigyeket, fajtól függően előfordulnak
- méregmirigyek, a nyílméreg békák curare nevű izombénító anyagát termelő mirigyek,
- pigmentsejteket, amelyek a hőszabályozásban és a rejtőzködésben, a figyelemfelkeltésben (riasztószín) játszanak szerepet (nyílméreg békák).
A bőralja csak néhány ponton kapcsolja össze a bőrt az izomzattal. Békáknál sok nagy méretű nyirokzsák található az irha és az izmok között, amelyekbe levegőt képesek préselni.
Táplálkozás
Lárváik növényevők vagy mindenevők, a felnőtt egyedek ragadozók.
- Elöl lenőtt, kicsapható, ragadós végű nyelvük van.
- Apró ránőtt fogazattal rendelkeznek.
- A nyelést egyes esetekben a szájüregbe süllyeszthető szemfenék segítheti.
- Gyomruk izmos szerv, itt történik a táplálék mechanikai roncsolása.
- Kloákájuk van.
Légzés
- Jelentős a bőrlégzésük (60%).
- Lárva állapotban (külső vagy belső) kopoltyúval lélegeznek.
- A légcső – farkos kétéltűeknél - kezdete a gégefő, amely a benne elhelyezkedő hangszalagoknak köszönhetően hangadásra képes. Mikor a tüdőből kiáramló levegő a hangszalagokat rezgésbe hozza, hang keletkezik. A hím békák a hangot a szájgarat üregből nyíló hanghólyagokban még felerősítik.
- A békáknak légcsövük nincs, mivel nincs nyakuk, így a tüdők közvetlenül a gégéből nyílnak.
- A fejletlen tüdők páros, zsákalakú képződmények, belső felületükön felület megnagyobbító lécek vannak. A tüdők hidrosztatikai szervként is szolgálnak.
Légcseréjük során „nyelik” a levegőt (nincs mellkas, nincs rekeszizom).
Belégzéskor
- az állat lesüllyeszti a szájfenekét, a szájüreg térfogata megnő, benne a levegő nyomása lecsökken, az orrnyílásokon keresztül levegő áramlik be,
- majd az orrnyílásait egy billentyűvel elzárja, a szájfenekét felemeli és így a szájüreg szűkítésével a nagynyomású levegő a tüdőbe préselődik.
Kilégzésnél a tüdők rugalmas fala összehúzódik és a megnyíló orrnyílásokon keresztül kiszorítja a levegőt.
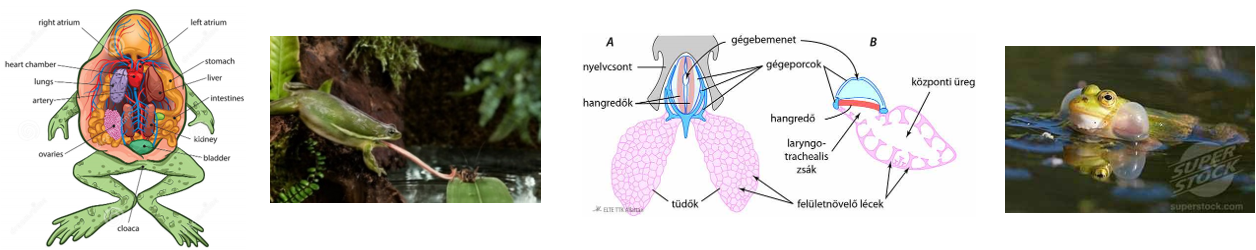 |
Keringés
- A lárvák keringése hasonló a halakéhoz, egyvérkörös.
- Kifejlett állatok keringése két vérkörös, szívük háromüregű - két pitvarral és egy kamrával.
- Két vérkörük a kisvérkör (tüdő vérköre) és a nagyvérkör.
- A kamrában az oxigén szegény és az oxigéndús vér részben keveredik, ugyanakkor mély zsebek gátolják a vér teljes keveredését.
- A kisvérkör a kamra jobb oldali zsebeiből indul, és vénás vért szállít a tüdők, valamint a bőr alatti hajszálerek felé (tüdő-bőr artéria, bőrlégzés).
- Az oxigéndús vér a bal pitvarba ömlik a tüdőből, innét kerül a kamra bal zsebeibe, ahonnét a nagyvérkörbe lépve bejárja az állat testét, ellátva oxigénnel és tápanyagokkal a sejtjeit.
- A testvénák a vénás öbölbe szállítják az enyhén kevert vért (a bőr alatti hajszálerekben a vér frissül, oxigénnel telítődik). A vénás öbölből a vér a jobb pitvarba kerül, innét pedig a kamra jobb oldali mélyedéseibe.
- Az aortában húzódó spirális billentyű a vér egy részét a nagy-, másik részét pedig a kisvérkörbe tereli.
Szaporodás
A megtermékenyítésük külső, a vízben zajlik.
- Tápanyagokban gazdag, kocsonyás burokkal körülvett nagyszámú petéiket általában vízbe rakják, és a lárvák fejlődése is itt történik (kivétel álelevenszülők, habfészket készítők).
- Fejlődésük közvetett, a lárvák átalakulással (metamorfózis) fejlődnek.
- A vízben fejlődő, halszerű lárvák (ebihal)
- kopoltyúval lélegeznek,
- kezdetben nincsenek végtagjaik,
- farkuk van,
- oldalvonalszervük van,
- először a hátsó, majd a mellső végtagjaik fejlődnek ki (békáknál).
Érzékszervek
A szárazföldi élethez való részleges alkalmazkodás az érzékszervek szintjén is megnyilvánul.
- A szemgolyó felépítése hasonló a többi gerinces osztály szeméhez (hólyagszem).
- Kialakultak a szemet a kiszáradástól megvédő szemhéjak és könnymirigyek.
- A szemek már képesek távoli tárgyakat is látni.
- A hallás és a helyzetérzés szerve a belső fülben található. Kialakul a középfül, benne egy hallócsonttal.
- A dobhártya, amely felfogja a levegő rezgéseit, a fej két oldalán a bőrfelszínnel egy síkban helyezkedik el.
- A szaglás helye az orr belsejében elhelyezkedő szaglóhám.
- A szájban érzőbimbók vannak, de ezek inkább tapintó, mint ízlelőszervek.
- A tapintás és hőérzékelés képletei a bőrben elszórt érzéksejtek, idegvégződések.
 |
Magzatburkos gerincesek
A hüllők, a madarak és az emlősök tartoznak ebbe a rendszertani kategóriába. A magzatburkok az embrió védelmét és táplálását biztosító képződmények, megvédik a fejlődő embriót a kiszáradástól és így lehetővé teszik a szaporodás, egyedfejlődés víztől történő elszakadását.
A három csoport közös sajátsága a szárazföldi életmódhoz való alkalmazkodás, az elsődleges vízi életmódtól való teljes elszakadás, amely egyrészt
- a kültakaró hámrétegének erőteljes elszarusodásának (amely gátolja a kiszáradást),
- másrészt a szaporodás víztől függetlenné válásának köszönhető (magzatburkok, belső megtermékenyítés).
Hüllők osztálya
A hüllők kb. 320 millió éve jelentek meg, ősi kétéltűekből alakultak ki a szárazföldi életmódhoz való alkalmazkodás eredményeképpen. Virágkorukat kb. 200 millió évvel ezelőtt a jura időszakban élték, az egész Földet meghódították, a szárazföldektől a vizeken át a levegőig.
Legsikeresebb csoportjuk a dinoszauruszok voltak, amelyek kb. 65 millió évvel ezelőtt a kréta időszakban máig vitatott körülmények között – legvalószínűbb elképzelés szerint egy meteorit Földdel való ütközése miatti klímaváltozás következtében – kipusztultak.
 |
Vázrendszer, mozgás
- A gerincoszlop első két csigolyája
- a fejgyám (atlasz) és
- a forgó (episztrófeusz).
- Az első két, speciális nyakcsigolyának köszönhetően fejüket jól forgatják.
- Fejlett a gerincoszlop, nyaki, háti, ágyéki keresztcsonti, farok szakaszokra tagolódik.
- A nyak és a farok kivételével majdnem mindenhol fejlett bordáik vannak.
- Zárt mellkasuk van, mivel a mellkasi bordákat elöl a szegycsont kapcsolja össze (kígyók kivételével).
- Ötujjú végtagtípus jellemző,
- az alkar (orsó, singcsont),
- és a lábszár (síp-, szárkapocscsont) dupla csont.
- Tolólábuk van.
Kültakaró
Bőrüket rendesen 3 réteg alkotja; a felhám, az irha, a jelentéktelen bőralja.
A felhám száraz, erősen elszarusodó laphám;
- melynek felületén elhalt sejtek alkotta, kiszáradás ellen védő, vastag szaruréteg található, ami szarupikkelyeket vagy szarupajzsokat hoz létre, amelyek az egész testet beborítják, jelentős mechanikai védelmet biztosítva,
- a növekedés miatt vedleniük kell,
- a kígyók egyben (kígyóing),
- a teknősök és krokodilok „folyamatosan vedlenek”,
- a gyíkok és a kaméleonok szakaszosan vedlik le külső szarurétegeiket, többnyire nagyobb darabokban.
Az irhában
- a kötőszövet elcsontosodásával létrejövő bőrcsontok lehetnek, melyek a teknősöknél összenőhetnek a gerincoszloppal, a bordákkal és a szegycsonttal, ez a teknő,
- továbbá sok pigmentsejt van, egyesek (kaméleonok) gyorsan változtatják a színüket.
A hüllők bőre mirigymentes (estleg előfordulhatnak illatmirigyek).
 |
Táplálkozás
Lehetnek
- növényevők - leguánok, egyes teknősök,
- ragadozók – krokodilok, kígyók, gyíkok stb.
- dögevők – varánuszok.
- Szájüregben gyökértelen, ránőtt fogazat található.
- A krokodiloknál gyökeres,
- kígyóknál méregfogak találhatók.
- Teknősöknek szarukávájuk van.
- Kloákájuk van.
Légzés
- A tüdő bonyolítja a gázcserét, bőrlégzésük nincs a vastag szaruréteg miatt.
- A tüdő egyszerű, zsákszerű képződmény, amelyet válaszfalak tagolnak kamrákra.
- A kígyóknál többnyire csak a jobb tüdőfél fejlődik ki.
- A légcsere során a mellkasuk térfogatát változtatják bordaközi izmok segítségével. Belégzéskor bordaközi izmok összehúzódnak, a mellkas tágul, a tüdőben levő levegő nyomása lecsökken, a levegő beáramlik. Kilégzés fordítva.
- Krokodiloknál megjelenik a rekeszizom.
Keringés
- Két vérkörös, zárt keringésük van.
- A legtöbb hüllőnek háromüregű szíve van, amelyet két pitvar és egy válaszfallal szinte teljesen kettéosztott kamra
- A tüdőből jövő, bal pitvarba kerülő, oxigéndús és a testből jövő, jobb pitvarba jutó oxigénszegény vér a kamrában alig keveredik, mivel azt egy válaszfal két kamrafélre tagolja.
- A kamrához egy pár aorta csatlakozik, melyben kissé kevert vér áramlik, melynek oxigéntartalma alacsony az állandó, magas testhőmérséklet kialakításához, ezért változó testhőmérsékletűek (nem hidegvérűek!).
- Egyes krokodiloknál megjelenik a kétkamrás szív.
Szaporodás
- Váltivarúak,
- belső megtermékenyítésűek,
- közvetlen fejlődésűek, embrionális fejlődésük tojásban zajlik, melyeket többnyire talajba ásnak, és a Nap melege költi ki azokat.
- Lehetnek álelevenszülők (egyes kígyóknál, pl. viperáknál), mikor a nőstények a tojásokat nem rakják le, hanem testükben, a petevezetőben tartják, melegítve, védve, majd mikor az utódok kikelnek, a kloákán keresztül távoznak az anya szervezetéből, látszólag a szüléshez hasonlóan. Azért álelevenszülés, mert az anya, ill. az embriók szervezete között semmiféle anyagcserekapcsolat nincsen, az utódok a tojásban felhalmozott, ún. szikanyagból növekednek.
- Párzás a farok tövénél található kloáka segítségével történik, párzószervek nincsenek.
- Tojásaikat bőrszerű vagy meszes (krokodilok), de lágy héj védi, az embriót magzatburkok veszik körül.
Érzékszervek
Megjelenik a külső fül, a külső hallójárat, azaz a dobhártya a mélybe süllyed.
Érzékszerveik rendkívül kifinomultak, különösen a kígyóké, amelyek Jacobson-féle szerve járulékos szaglószervként segíti a nyelv öltögetésével a szájba juttatott illatanyagok érzékelését.
Némelyik kígyófaj (pl. óriáskígyók, csörgőkígyók) a zsákmány felderítését megkönnyítő infravörös sugarakat (hősugarakat) érzékelő gödörszervvel is rendelkezik.
 |
Madarak osztálya
A tollas dinoszauruszokból alakult ki a legkorábbi ismert madár, a jurakori (150 millió éve), galamb nagyságú Archaeopteryx. Testfelépítése átmenetet mutat a mai madarak és az egykori hüllők között, mindkét csoportra jellemző sajátságokat mutat (egyesek szerint a madarak a hüllők egyik csoportja, másképp; a túlélő dínók egy csoportja.)
Hüllőbélyegek:
- fogazat,
- gerincoszlopot is tartalmazó fejlett farok,
- mellső végtagon karomban végződő, szabadon álló ujjak.
Madárbélyegek:
- tollazat,
- szárny,
- csűdcsont.
A madarak főbb jellemzői
Testük felépítése jól tükrözi a repülő életmódhoz való alkalmazkodást:
- mellső végtagjaik szárnyakká módosultak,
- farkuk csökevényes, a test súlyát csökkentve,
- csontjaik üregesek, légzsákokat tartalmaznak, könnyűek, szegycsontjuk tarajos,
- testüket tolltakaró borítja,
- fogak nincsenek, a szájnyílást csőrkávák veszik körül, melyeket szaruképletek, az ún. szarutokok borítanak (súlycsökkentés),
- fejük kis méretű - ezért és a fogazat hiányának köszönhetően -, kis tömegű,
- az állatvilág legfejlettebb tüdejével rendelkeznek,
- tüdejükhöz légzsákrendszer kapcsolódik, testük fajsúlya kicsi,
- nagy, négyüregű szívük, szeparált két vérkörük van, ahol az oxigén-dús és az oxigén-szegény vér nem keveredik,
- ezért testhőmérsékletük viszonylag magas és állandó (magas alapanyagcsere szint).
Ezenkívül jellemzően
- meszes héjú tojásokkal szaporodnak,
- fejlett az ivadékgondozásuk,
- a Föld valamennyi tengerén és szárazföldjén előfordulnak, közel 9700 madárfaj ismert.
- A madarak csontjai - szemben az emlősök csontvelőt tartalmazó csontjaival - levegő tartalmúak, aminek köszönhetően a madártest fajsúlya csekély.
- Általánosan jellemző a fogatlan csőr és a nagy méretű szemüreg.
- Számos (11-24) nyakcsigolyája igen lazán kapcsolódik egymáshoz, így a madárnyak rendkívül mozgékony.
- Ezzel szemben az utolsó hátcsigolyák, az ágyék- és keresztcsonti csigolyák összecsontosodtak (álkeresztcsont, far-hát), ami a repüléshez és a két lábon járáshoz szilárd alapot biztosít.
- A bordák horgokkal kapcsolódnak össze, ami stabilizálja a mellkast a repüléshez.
- A futómadarak kivételével az összes madárfaj szegycsontján jellegzetes tarajt találunk, amely nagy eredési felületet eredményez a repülőizmoknak (futómadaraknál nincs).
- A mellső végtagban (szárny) a kéztőcsontok a kézközépcsontokkal nagyrészt összenőnek és megnyúltak. Három ujjuk közül az első az ún. fiókszárnyat hordozza, a többi ujj pedig csökevényes.
- A hátsó végtagban a combcsontot, a sípcsontot és a vékony szárkapocscsontot, a csüdcsontot (lábközép és lábtőcsontok összenövéséből) és a lábujjperceket különíthetjük el.
- A legtöbb madárfajnak 4 lábujja van (3 előre, 1 hátra áll), de a futómadarak esetében ez redukálódhat 3 vagy 2 lábujjra.
- Gázlóláb – fehér gólya
- Markolóláb – réti sas
- Evezőláb tőkés réce
- Futóláb – túzok
- Kapirgálóláb – vadászfácán
- mechanikai védelmet nyújt,
- fontos szerepe van a hőháztartásban,
- csökkenti a párologtatást,
- zsíros, vízhatlan bevonata révén megakadályozza a bőr felázását.
- Szaruképleteik: tollak, karmok, csüdön pikkelyek, a szájnyílás körül a csőrkávákat borító szarutokok.
- Irha alatt változó vastagságú zsírszövet található, amely tápanyagraktár, hőszigetelő szerepet tölt be.
- A kültakaró általában mirigyeket nem tartalmaz, egyetlen bőrmirigyük – a tollazat vízhatlanná tételére szolgáló zsíros anyagot termelő – a farkcsík mirigy, a faroktollak tövében található.
- bonyolult felépítésű, könnyű szaruképletek,
- vízhatlanná teszik az állat felületét,
- jó hőszigetelők,
- mechanikai védelmet nyújtanak,
- nagy felületet biztosítva lehetővé teszik a repülést.
- fedő- (testen),
- evező- (szárnyon),
- kormánytollak (farok).
- A pehelytollak a kontúrtollak alatt vannak, hőszigetelők.
- A fiókák első tollazata.
- a toll irhában levő része a cséve,
- a hámból kiemelkedő része: tollszár + tollágak + sugarak, melyeket horgok tartanak össze egy síkban, az ún. zászlót alkotva.
- Vésőcsőr – fakopáncs
- Magevő csőr - pintyek
- Horgas csőr - sólyomalkatúak
- Lemezes csőr – lúdalkatúak
- Vágócsőr – gólyaalkatúak
- a táplálékot puhítja, raktározza,
- szaporodási időszakban termeli a zsírokban, fehérjékben gazdag begytejet, ami a fiókák táplálására szolgál.
- a mirigyes gyomorban megkezdődik az emésztés,
- az izmos zúzógyomorban lenyelt kövek segítségével történik a táplálék felaprítása (fogazat hiánya).
- Kerigési rendszerük zárt, két vérkörük van (természetesen).
- Fokozott anyagcsere folyamatainak alapja a 4 üregű szív, munkavégzése rendkívül gyors.
- Teljesen elkülönült a 2 vérkör, mivel az artériás és a vénás vér nem keveredik a szívben a két kamrát elválasztó falnak köszönhetően, így a test szöveteihez magas oxigéntartalmú vér jut, ami az egyik feltétele az állandó testhőmérséklet kialakulásának (a másik, az agyi hőszabályozó központ kifejlődése).
- A gyors anyagcserével összefügg a madarak magas testhőmérséklete (40-44 C fok), amely az állatvilágban csaknem egyedülálló.
- Váltivarúak, ivari kétalakúsággal
- belső megtermékenyítésűek,
- közvetlen fejlődésűek.
- Embrionális fejlődésük meszes héjú tojásban zajlik.
- Ivadékgondozók, fészeklakók vagy fészekhagyók.
- petesejtjéből, a tojássárgájából, az azt körülvevő
- fehérjéből,
- kettős héjhártyából és
- meszes héjból épül fel.
- felfogja a különféle mechanikai hatásokat, tompítja a hőmérséklet-változások hatásait,
- fehérjeforrás,
- víz- és ionraktár.
- Fülkagylójuk nincs. Középfülükben egy hallócsont van.
- Belső fülük, így hallásuk - bár csigavezetékük nem felcsavarodott, hanem csak görbült - és egyensúlyérzékelésük rendkívül fejlett.
- Szaglásuk, ízlelésük fejletlen.
- Szemük valamennyi élőlény közül a legtökéletesebb.
- Belső szilárd váz,
- a test alakját és méretét határozza meg,
- részt vesz a látás, a szaglás, valamint a hallás és az egyensúlyozás érzékszervei, továbbá az agy védelmében,
- a tüdő működéséhez külső támaszt szolgáltat,
- helyet biztosít a vörös csontvelőnek és így részt vesz a vérképzésben,
- a szervezet kalciumraktára.
- Az egyik legfontosabb szabályozó a parathormon, amely a mellékpajzsmirigy sejtjeiben termelődik. A parathormon fő hatása a vér-, ill. az extracelluláris kalciumszint növelése. Elválasztását a vér alacsony kalciumszintje serkenti. A csontra hatva - a csontfaló sejtek aktiválásán keresztül - fokozza a csontok kalciumleadását, valamint a vesében serkenti a kalcium visszaszívását. Mindezek mellett serkenti az aktív D-vitamin szintézisét a vesében, így közvetve elősegíti a bélből a Ca2+ felszívódását.
- A kalcitriol a D-vitamin aktív alakja. A kalcitriol a bélben növeli a kalcium-, valamint a foszfátfelszívódást, továbbá fokozza a csontok Ca2+-felvételét.
- A kalcitonin a pajzsmirigyben termelődik. Fő szerepe a vér kalciumszintjének csökkentése a csontbontás gátlása és a kalciumionok csontokba való beépülésének elősegítése révén. Termelődését a vér magas Ca2+-szintje serkenti.
- Az ösztrogének gátolják a csontfaló sejtek működését, miáltal biztosítják a csontok megfelelő állapotát, idősebb korban az ösztrogénhiány csontritkulást eredményez.
- szerves (40%) és
- szervetlen (60%) állományra,
- sejtekre és
- sejtközötti állományra
- csontsejtek,
- kollagénrostok,
- erek, idegek építik fel.
- a sejtközötti állomány szervetlen kalciumsói (40%), mint pl. Ca5(PO4)3 X, ahol X= F-, OH- (apatitok), CaCO3 és
- víz alkotják (20%).
- Amennyiben a szervetlen állományt sósavas kezeléssel kioldjuk, a csontok elvesztik keménységüket, gumiszerűen hajlíthatók lesznek.
- A szerves állományt égetéssel távolíthatjuk el, ekkor a csontok, elvesztve rugalmasságukat, ridegek és törékenyek lesznek.
- rostos kötőszövetből áll,
- innen erednek a csontszövetet ellátó erek és idegek.
- A csont védelmét, táplálását biztosítja,
- innen indul ki a csontregeneráció,
- lehetővé teszi a csontok vastagodását.
- a csöves csontok végein,
- továbbá a lapos és a köbös csontok belsejében található.
- foszfát-, kollagén- és víztartalma csökken,
- karbonáttartalma nő.
- a végtagokban találhatók, hosszú, belül üreges csontok,
- végükön kiszélesedők, ízületek képzésében vesznek részt.
- Középső részükben, a velőüregben sárga csontvelő, végükön a szivacsos állományban vörös csontvelő található.
- Hosszú csöves csontok pl. a felkarcsont, a combcsont, rövid csöves csontok pl. a kézközépcsontok, lábközépcsontok.
- kétdimenziós csontok,
- belsejüket szivacsos állomány és vörös csontvelő tölti ki,
- vérképzés folyik bennük.
- Ilyen a lapockacsont, az agykoponyacsontok, a medencecsont, a bordák.
- háromdimenziós csontok,
- szivacsos állománnyal és vörös csontvelővel, a vérképzés fő helyei,
- ilyenek pl. a csigolyatestek, a kéz- (8 db) és lábtőcsontok (7 db).
- A csontos összenövéseknél az eredetileg önálló csontok az életkor előrehaladásával egybeforrnak, ilyen fordul elő pl.
- a keresztcsontnál (5 csigolyából),
- a medencecsontnál (csípőcsontból, szeméremcsontból, ülőcsontból, 18 éves korra),
- idősebb korban az arc- és agykoponyacsontoknál.
- A porcos összeköttetéseknél a csontokat a közéjük ékelődő porc köti össze, ilyen kapcsolat van pl.
- a csigolyák (porckorongok) és a szeméremcsontok között.
- A szalagos, kötőszövetes összeköttetések esetén mikroszkopikus kötőszöveti szalagok kapcsolják össze az érintkező csontokat, ilyenek pl.
- a koponya varratai.
- felszíne az ízületi felszín, melyet üvegporc borít.
- Az egyik csontvég domború (ízületi fej),
- a másik homorú (ízületi árok, vápa) felszínű.
- a keresztcsont és a csípőcsont közötti ízület, ill.
- a szeméremcsontok közötti kapcsolat (szülés), mely egyes források szerint átmenetet képez a folytonos összeköttetés és az ízületes kapcsolódás között.
- az agykoponyából és
- arckoponyából.
- homlokcsont,
- ékcsont,
- nyakszirtcsont,
- falcsontok (páros),
- halántékcsontok (páros).
- Oldalnézetben kétszeres S görbületű, ez a koponya rugalmas alátámasztását teszi lehetővé, ami pedig a két lábon járás miatt fontos.
- Szelvényezett felépítésű, csigolyákból (32-35) áll.
- A koponyával érintkező első két nyakcsigolya, a fejgyám (atlas) és a forgó (axis) különleges alakja teszi lehetővé a fej nagyfokú mozgékonyságát a gerinchez képest.
- A mozgékony csigolyák porckorongokkal, kötőszöveti szalagokkal, ízületekkel vannak összekapcsolódva.
- a nyaki 7,
- a háti 12,
- az ágyéki 5,
- a keresztcsonti 5,
- a farokcsonti 3-6 csigolyából áll.
- a jelentős szivacsos állományt – benne vörös csontvelőt - tartalmazó csigolyatestet,
- a csigolyaívet, ami a csigolyalyukat határolja,
- a hátsó tövisnyúlványt, mely a hátizmok tapadási felületét adja,
- a harántnyúlványokat, amikhez a bordák ízesülnek,
- az ízületi nyúlványokat, amelyek a csigolyák egymáshoz ízesülésének a helyei.
- A csigolyalyukak összessége a gerinccsatornát képezi, ahol a gerincvelő foglal helyet.
- A 7 pár valódi borda külön porccal ízesül a szegycsonthoz,
- a 3 pár álborda közös porccal kapcsolódik egymáshoz és a 7. borda porcán át szegycsonthoz,
- a 2 pár repülőborda nem kapcsolódik egyáltalán a szegycsonthoz, végük a hasfal izomzatába ágyazódik.
- a kulcscsont a szegycsonthoz és a lapockához ízesül,
- a lapocka a hátizomba ágyazódik.
- a felkarcsont a lapockához ízesül,
- az orsócsont (hüvelykujj felé),
- a singcsont,
- a kéz csontjai,
- kéztőcsontok (8db) köbös csontok,
- kézközépcsontok,
- ujjpercek, ujjanként jó esetben 3, a hüvelykujj esetén 2.
- a keresztcsontból, ami 5 csigolyából nő össze és
- a páros medencecsontból.
- a csípőcsontból,
- a szeméremcsontból és
- az ülőcsontból nő össze.
- a combcsont,
- a lábszárban a sípcsont (hüvelykujj felé) és a szárkapocscsont,
- a láb csontjai pedig
- a lábtőcsontok (7 db),
- a lábközépcsontok és
- az ujjpercek.
- zárt, amikor a bőr felszíne nem sérül, a törtvégek a bőr alatt vannak,
- illetve nyílt, ha a bőr átszakad és a törtvégek láthatóvá válnak.
- a csontrendszer passzív módon,
- a vázizomrendszer aktívan vesz részt a mozgások kivitelezésében.
- az elmozduló csonton van az izom tapadási helye,
- a másik csonton található az eredési hely.
- orsó alakúak a végtagokon,
- laposak a törzsön,
- gyűrűszerűek a testnyílások körül.
- mimikai izmok, rágóizmok, fejbiccentő izom,
- nagy mellizom, deltaizom, egyenes hasizom, ferde hasizom, bordaközi izmok, rekeszizom, nagy farizom, gátizmok (gát: a végbélnyílástól a külső nemi szervekig terjedő terület, alátámasztja a kismedencei és hasüregi szerveket)
- bicepsz, tricepsz, négyfejű combizom, hátulsó lábszárizom (vádli, lábikra).
- a gerincesek és az ízeltlábúak vázát mozgató izmokban,
- a nyelvben, a garatban, a nyelőcső felső szakaszában, a végbélnyílás és a húgycső kezdete körül,
- továbbá ilyenek a légzőizmok (rekeszizom, bordaközi izmok).
- többnyire akaratunktól függően működik (kivétel pl. vázizomreflexek (térdreflex)),
- összehúzódása gyors,
- nagy erőkifejtésre képes,
- de fáradékony.
- A harántcsíkolt vázizomszövet szöveti egysége a hajszálszerű, akár több ezer sejtmagot is tartalmazó, óriás sejteknek tekinthető izomrost (muscle fiber).
- Az izomrostokat membrán határolja, mely helyenként betüremkedve eléri a SER-t.
- Sok izomrost köteggé – izomnyalábbá – szerveződik, amelyet kötőszövetes hártya (perimysium) tart össze,
- az izomrostkötegek tovább rendeződve alkotják az izmokat, amelyeket szintén egy kötőszövetes lemez, az ún. izompólya (epimysium) határol.
- izomfonalkötegeket (miofibrillumokat),
- amelyek izomfonalakból (miofilamentumokból) állnak.
- Az izomfonalkötegek között nagy mennyiségű mitokondrium helyezkedik el.
- Az ugyancsak nagyszámú sejtmag perifériásan az izomrost membránja alatt található.
- A Ca-raktárként működő endoplazmatikus retikulum(SER) harisnyaszerűen veszi körbe az egyes izomfonalkötegeket.
- a vékony fonal csak a világos szakaszokat építi fel,
- a vastag fonal nagyrészt a sötét sávokban található.
- két globuláris feji részből,
- mely tartalmazza az aktinkötő centrumot,
- ATP-bontó aktivitása van,
- és a fejhez kapcsolódó fibrilláris részből.
- Az izomrostokat beidegző idegrostok végződéseiből felszabaduló ingerületátvivő anyagok hatására az izomrostok citoplazmájában Ca2+-ok szabadulnak fel (SER-ből), aminek hatására a miozinmolekulák feji részei hozzákötődnek az aktinfonalakhoz.
- A kötődést követően a miozin feji részei bólintó mozgást végeznek (konformációváltozás), aminek köszönhetően a vékony fonalak beljebb csúsznak a vastag fonalak közé, egységnyi távolságot megtéve.
- Ezután az ATP és Mg2+ megkötésekor a miozinfejek leválnak az aktinfonalakról.
- ATP bontásakor felszabaduló energia hatására a miozin fejek kiegyenesednek, majd már egy másik kötőhelyen ismét hozzákötődnek a vékony fonalakhoz és kezdődik minden elölről.
- izotóniás, ha az izom megrövidül,
- izometriás, ha a rostok feszülnek, azonban az izom nem rövidül meg.
- Az első 7 másodpercben az ATP-kreatin-foszfát rendszer a domináló,
- míg a 7-60 másodperc közötti időszakban a glikolízis,
- majd a biológiai oxidáció veszi át a főszerepet.
- a szénhidrátraktárak kimerülése és az ebből fakadó energiahiány,
- az izomrost pH-jának csökkenése,
- a sejten kívüli tér K+-tartalmának megemelkedése,
- az izomrost plazmájában levő Ca2+-mennyiségnek, vagy egyes molekulák Ca2+-érzékenységének megváltozása.
- lassú, tartós összehúzódásra képes, magas mioglobin tartalmú ún. vörös izmokat, ilyenek a testtartásért felelős izmok, pl. hátizmok, ill.
- gyors, fáradékonyabb ún. fehér izmokat, ilyenek a végtagok izmai.
- akaratunktól függetlenül működik,
- gyors,
- nagy erőkifejtésre képes,
- nem fáradékony.
- egymagvú izomsejtekből áll, melyek elnyújtott térrácsot alkotnak, a sejtek helyenként elágaznak,
- a sejtmagvak a sejt közepén helyezkednek el.
- a gerincesek zsigeri izomzata: erek falában, bélcső falában, légcső falában stb.,
- bőrben,
- egyes gerinctelenek - férgek, puhatestűek - bőrizomtömlőjében.
- akaratunktól függetlenül működik,
- lassú,
- kis erőkifejtésre képes,
- nem fáradékony.
- az emésztőcsatorna üregébe elválasztott enzimek indítják el, pl. a gyomorban termelődő pepszin (luminalis emésztés),
- majd a monomerekre való végső bontást a bélhámsejtek felszínéhez kötött enzimek végzik, pl. a vékonybél nyálkahártyájának hámfelületéhez kötött membránpeptidázok (celluláris emésztés).
- nyaki szakaszában találjuk a mucinózus nyálkát termelő melléksejteket,
- a középső régióiban helyezkednek el a H+ és Cl- ionokat termelő, ún. fedősejtek,
- az alsó részben vannak a pepszinogént (és gyomorlipázt) termelő, ún. fősejtek.
- a fültőmirigy,
- a nyelv alatti mirigy,
- az állkapocs alatti mirigy.
- a nyálamiláz enzim, melynek feladata a keményítő, glikogén maltózzá bontása,
- a mucin, melynek feladata a táplálékpép összeragasztása, a falat képzés elősegítése, a nyál viszkozitásának növelése.
- A nyál ezen kívül még anorganikus sókat is tartalmaz: Na+, K+, Ca2+, Cl-, PO43-.
- egyrészt aktiválják a zsírbontó lipázokat,
- másfelől a bélmozgások miatt felaprózódott zsírcseppek felületén elhelyezkedve csökkentik az emulgeált zsírgolyócskák felületi feszültségét, stabilizálják azokat, és fajlagos felületüket nagyobbítva kedvezőbb feltételeket teremtenek a lipáz emésztés számára (hatásuk a szappanokéhoz hasonló).
- testépítő anyagok, pl. fehérjék,
- energiaforrások, pl. szénhidrátok, zsírok,
- enzimek koenzimjeinek összetevői, pl. vitaminok,
- enzimek prosztetikus csoportjai, pl. egyes ásványi anyagok.
- A fiziológiás éhségérzet kialakulása – a hipotalamuszban – alapvető biológiai szükséglet kielégítését teszi lehetővé, ettől meg kell különböztetnünk
- a mentális éhség fogalmát, amit étvágynak nevezünk, mely létrejöttének hátterében inkább külső tényezők – stresszhatások, ingerszegény környezet (unalomevés), különféle beidegződések-társítások (TV, moziéhség, egy kedvenc étel látványa, szaga, íze, emléke) stb. – keresendők.
- az éhségközpont, amelynek aktivitása éhségérzetet kelt és ez a táplálék felvételét eredményezi, ill. ugyanitt van
- az ellentétes hatású jóllakottsági központ.
- elsősorban az alacsony vércukorszint
- és olyan peptidhormonok – pl. ghrelin szöveti hormon – váltják ki, amiket a
- gyomor üressége esetén elsősorban a tápcsatorna fala választ el.
- mivel hat a májra és a hasnyálmirigyre, elősegíti a tápanyagok glikogén, ill. zsír formájában történő raktározását,
- a központi idegrendszerben serkenti az agy jutalmazó központjában a dopaminfelszabadulást, aminek hatására fokozódik az evéssel együttjáró örömérzet.
- a gyomor teltsége (mechanoreceptorok révén),
- a magas vércukorszint és inzulinszint, továbbá
- a zsírszövet sejtjeiben termelődő leptin szöveti hormon.
- az élelmiszer szénhidráttartalmának emészthetősége
- és a felszabaduló egyszerű cukrok felszívódásának a sebessége határozza meg.
- lebonyolítja a külső gázcserét, azaz
- biztosítja a biológiai oxidációhoz szükséges oxigén felvételét,
- a lebontó anyagcserében keletkezett szén-dioxid leadását,
- lehetővé teszi a hangképzést,
- intenzív légcsere során részt vesz a hőleadásban,
- a légzőmozgások – belégzés – segítik a vér szív felé történő áramlását az ún. negatív mellűri nyomás – a légköri nyomásnál kisebb – kialakítása révén.
- gázcseréről, ill.
- légcseréről.
- külső, ha a gázok kicserélődése a testfolyadék – pl. vér – és a levegő között zajlik a légzőszerv – tüdő – felületén.
- Belső gázcseréről akkor beszélünk, ha a légzési gázok a testfolyadék (vér) és a szövetek között cserélődnek ki a kapillárisok határfelületén.
- a légutak,
- a tüdő,
- a mellhártyák,
- a kisvérköri erek,
- a mellkas, a légzőizmokkal együtt, ill.
- a légzésszabályozásban résztvevő idegrendszeri képletek, receptorok.
- az orrüreg, a szájüreg,
- a garat, ami a légzőrendszer és a tápcsatorna közös szakasza,
- a gége,
- légcső,
- főhörgők, egyre kisebb hörgők, hörgőcskék alkotják,
- melyek a léghólyagokba torkollva végződnek.
- csontos – orrcsont –,
- porcos,
- részben tömör kötőszövet alkotja,
- kívülről bőr, belső részét nyálkahártya borítja, sok faggyúmiriggyel és szőrtüszővel.
- két nagyobb üregből és
- az azokból nyíló melléküregekből áll.
- az arcüreg,
- a homloküreg,
- a rostacsontok üregei,
- az ékcsonti üreg.
- a nyálkahártya bő vérellátása révén az áthaladó levegő felmelegítése,
- nagyszámú mirigye miatt a párásítása,
- a szőrzete, ill. a csillózata pedig a pormentesítést biztosítja.
- Rezonátorüregként részt vesz a hangképzésben,
- ellátja a szaglóműködést.
- porcokból,
- kötőszövetes lemezekből,
- izmokból álló,
- nyálkahártyával bélelt üreges szerv.
- a pajzsporc,
- a gyűrűporc,
- a kannaporcok és
- a gégefedőporc.
- laphámsejtek, ezek képezik a légzőfelületet, továbbá
- fagocitáló sejtek (makrofágok), melyek a léghólyagokig eljutott porszemcséket kebelezik be, ill.
- felületaktív anyagot termelő sejtek, ezek váladéka csökkenti a léghólyagocskákat borító folyadékfilm felületi feszültségét, s megakadályozza, hogy a nagyon kis átmérőjű (10 μm) léghólyagocskák felszíne kilégzéskor összetapadjon.
- a tüdő hajszálerei,
- továbbá olyan kötőszöveti sejtek, amik a tüdő falában található nagy mennyiségű
- rugalmas rostot termelik.
- bordaközi izmok és
- a rekeszizom működésének köszönhető.
- a külső bordaközi izmok felemelik a mellkast, ill.
- a kupolaszerű rekeszizom összehúzódva ellaposodik.
- a mellüreg térfogata megnő,
- a tüdő kitágul,
- s benne a nyomás lecsökken,
- így a levegő kívülről a tüdőbe áramlik.
- a mellkas visszatér alaphelyzetébe,
- térfogata csökken,
- a tüdőben a levegő nyomása megnő,
- aminek következtében kiáramlik a tüdőből.
- a belső bordaközi izmok, amelyek összehúzzák a mellkast, ill.
- a hasizmok, amelyek a belekre gyakorolt nyomásuk révén erősebben bedomborítják a rekeszizmot a mellüregbe (hasprés).
- Nyugodt légzés esetén az egyszeri légvétellel ventillált 0,5 l (dm3) levegőt légzési térfogatnak nevezzük.
- A légvételenként kicserélt levegő térfogatát megszorozva a percenkénti légvételek számával, megkapjuk a légzési perctérfogatot. Nyugodt légzés esetén a perctérfogat 8 l, mivel percenként 16-szor veszünk levegőt, légvételenként 0,5 l levegőt kicserélve.
- Fizikai, ill. pszichés terhelés hatására a perctérfogat jelentősen fokozódhat, ami egyrészt a megnövekedett légzésszámnak, ill. a megnövekedett légzési térfogatnak köszönhető.
- Erőltetett belégzéskor további 2,5 l levegőt tudunk beszívni, amit belégzési tartaléknak hívunk.
- Erőltetett kilégzés során 1,5 l levegőt tudunk még kipréselni a tüdőből, ezt kilégzési tartaléknak tekintjük.
- Ekkor még mindig marad levegő a tüdőben, amit semmilyen körülmények között nem tudunk kifújni, ez a maradék levegő, kb. 1 l.
- Az egy légvétellel maximálisan beszívott, majd erőteljesen kifújt levegő térfogatát vitálkapacitásnak nevezzük, kb. 4,5 l.
- az edzett szervezet a légzésszám növekedése mellett a légzések mélységének növelésével,
- az edzetlen szervezet inkább a légzésszám fokozásával biztosítja.
- egy olyan harang alakú üvegbúrával (mellkas) végezzük,
- amit alulról egy gumimembrán (rekeszizom),
- felülről pedig egy kétfuratú gumidugó zár el.
- Az egyik furatban egy elzárható üvegcső található (melynek nyitásával a légmellt demonstrálhatjuk),
- a másikban egy olyan üvegcső (légcső), amelyre
- egy rugalmas léggömb (tüdő) csatlakoztatható.
- A Donders harang és a léggömb közötti tér a mellhártyák közötti teret,
- a léggömb belső tere a tüdő belső terét modellezi.
- örökletes tényezők,
- különféle fizikai-kémiai környezeti ártalmak, ill.
- kórokozó szervezetek.
- az allergiás eredetű, ún. szénanátha és
- a heveny fertőzéses, hurutos, közönséges nátha.
- növényi pollenek vagy különféle penészek, ill. háziporok – poratkák ürüléke – okozhatják.
- Tünetei az orrfolyás, szemkönnyezés, tüsszögés, viszketés.
- hurutos – intenzív váladéktermeléssel járó – gyulladása, a leggyakoribb megbetegedések egyike, legtöbbször egyszerű meghűlésből ered,
- bakteriális, ill. vírusfertőzés okozza.
- bakteriális vagy virális eredetű.
- Jellemző, hogy a garatot bélelő nyálkahártya gyulladt, továbbá láz, és a nyaki nyirokcsomók duzzanata tapasztalható.
- Kezelése leginkább antibiotikumokkal történik.
- az emberi beszédhang színezete megváltozik, a beteg nem tudja teljesen kontrollálni a hangképzést és fokozni a hangerejét.
- A rekedtség közvetlen oka a hangszálak megvastagodása, melynek a hátterében egyszerű gége- és hangszálgyulladás áll. A rekedtség társulhat felső légúti fertőzéshez, de okozhatja túlzott mértékű dohányzás vagy a hangszalagok túlerőltetése is.
- a hörgők fokozott váladéktermelésével járó gyulladásos megbetegedése.
- Leggyakrabban valamilyen vírus vagy baktérium okozza, általában a nátha szövődménye. Ha nem gyógyul, a folyamat előrehaladtával tüdőgyulladás alakulhat ki.
- A leggyakoribb tünetek a köhögés, rossz közérzet, hátfájás, váladékképződés, hőemelkedés.
- az alsó légutak krónikus gyulladásos betegsége,
- amit rohamokban jelentkező nehézlégzés kísér. Asztmás roham alatt a hörgőfal simaizmai összehúzódnak, a hörgők keresztmetszete beszűkül, sűrű, tapadós váladék zárja el a levegő útját, ami köhögést, sípoló kilégzést vagy mellkasi szorító érzést, légszomjat eredményez.
- A tüneteket leggyakrabban allergia okozza, de kiválthatja légúti vírusfertőzés, levegőszennyezés, fizikai terhelés és pszichés tényezők egyaránt.
- okozója a Koch-féle baktérium. A kór cseppfertőzés útján terjed.
- A betegség elsődlegesen a tüdőt támadja meg, de ha a baktériumok a vérárammal szétszóródnak, megtámadhatják a központi idegrendszert, a nyirokrendszert, a keringési rendszert, az ivarszerveket, a húgyutakat, a csontokat, az ízületeket és még a bőr felületét is. A tbc-s beteg tüdejében rögök, gümők A gümő latinul tuberculum, innen ered a betegség neve (gümőkór).
- Tünetei: mellkasi, ill. hátfájás, vér felköhögése, nem szűnő köhögési rohamok, valamint láz, testsúlycsökkenés és gyakori fáradtság.
- A tuberkulózis megelőzésére szolgáló BCG-oltás Magyarországon kötelező, kezelése antibiotikummal történik. A kórokozó fertőzőképességétől, ill. a szervezet immunrendszerének állapotától függ, hogy a szervezetbe jutott baktérium képes-e betegséget okozni. Általában a legyengült immunrendszerű, rossz egészségügyi állapotban lévő, nem megfelelő higiéniás viszonyok között élő emberek vannak jobban kitéve a fertőzés veszélyének.
- a léghólyagokat és a környező szöveteket érinti. A tüdőgyulladás nem egyféle betegség, hanem sok fajtája van, amelyeket különböző mikroorganizmusok, vírusok, baktériumok, gombák és egyéb környezeti ártalmak okoznak.
- A tüdőgyulladást elsődlegesen a belégzéssel bejutott, az ép tüdő felületén megtapadt kórokozók váltják ki. A másodlagos tüdőgyulladás az influenza egyik leggyakoribb szövődménye. Az influenza miatt a légúti hám sérülései teszik könnyebbé a gyulladást okozó baktériumok behatolását. Ritkábban maró gáz, füst vagy por, permet is okozhat tüdőgyulladást.
- Leggyakoribb tünetei a köpettel járó köhögés, mellkasi fájdalom, láz és nehézlégzés. Tüdőgyulladás gyanúja esetén a mellkast sztetoszkóppal hallgatják meg. A tüdőgyulladás rendszerint megváltoztatja a sztetoszkópban hallható légzési hangokat, a tüdőből szörtyögésre emlékeztető hang hallható. A tüdőgyulladás diagnózisát a mellkasröntgen erősíti meg.
- az agy bizonyos területein az idegsejtek nikotinikus acetilkolin-receptoraihoz kötődik,
- fokozza a szívműködést, emeli a vérnyomást,
- oldja a szorongásokat, javítja a kedélyállapotot.
- a tüdő-, a gége-, a szájüreg-, a nyelőcső- és a húgyhólyagrák egyik fő rizikófaktora.
- A tüdőrákos megbetegedések 80%-áért felelős.
- A dohányfüsttel bekerült szén-monoxid, mivel jóval erősebben kötődik a hemoglobinhoz, mint az oxigén, megakadályozza a vörösvértestek oxigénszállítását.
- A dohányfüst a légutak nyálkahártyáját fokozott nyálkatermelésre serkenti, ami pedig köhögésre ingerel, krónikus hörghurutot okoz, továbbá gátolja a légutak csillóinak működését, ezért a szennyeződések eltávolításának hatékonysága csökken.
- A léghólyagocskák közötti válaszfalak felszívódnak tüdőtágulatot eredményezve (COPD), aminek következtében jelentősen csökken a légzőfelület.
- Csökken a tüdőben a felületaktív váladék termelődése is, ami a léghólyagocskákat borító folyadékfilm felületi feszültségének növekedését és a légzőfelszínek összetapadását eredményezi.
- Megnő az agyvérzés, a szívkoszorúér betegség, az infarktus, az érszűkület kialakulásának kockázata.
- A várandósság alatti dohányzás növeli a magzati halálozás, a koraszülés, a fejlődés visszamaradásának a valószínűségét, a szoptatás alatt károsítja a csecsemő egészségét, mivel az anyatejbe bekerülnek a dohányfüst méreganyagai.
- A dohányfüstös környezet a nemdohányzók egészségét is veszélyezteti (passzív dohányzás).
- akkor beszélünk, ha egészséget károsító vagy életveszélyt okozó gázok – pl. szén-monoxid, szén-dioxid, nitrogén-oxidok, klórgáz stb. – kerülnek a szervezetbe.
- Ezek egy része tüdővizenyőt okoz, ami által nehezítik az oxigén vérbe történő bejutását, mások a hemoglobinhoz erősen kötődve megakadályozzák az oxigénszállítást.
- A segítségnyújtás első lépése a mérgezés okának a megszüntetése, a sérült szabad levegőn történő elhelyezése.
- egy globin fehérjerészből és
- egy hem nem fehérje természetű részből áll. A hem 4 pirrol gyűrűből összekapcsolódott porfirinváz, melynek közepén Fe2+ helyezkedik el.
- a szén-dioxid a szövetekből a vérplazmába diffundál, a vérplazmában a szén-dioxid hidrát alakban van jelen CO2.H2O,
- onnan a vörösvértestekbe jut, ahol egy enzim hatására szénsavvá alakul,
- a szénsav elbomlik H+ és hidrokarbonát-ionra (HCO3-),
- a protonokat a hemoglobin - oxigén leadása mellett – felveszi,
- a felesleges HCO3- kidiffundál a vérplazmába, ott szállítódik, helyette a töltések egyensúlya miatt a sejt Cl--t vesz fel.
- megindul a vérplazmából a CO2 felszabadulása a tüdő légterébe,
- a vérplazmában a csökkenő mennyiségű CO2 pótlása a vörösvértest felől történik.
- A vörösvértestben mivel csökken a CO2 ezért a szénsav elbomlik vízre és szén-dioxidra, ami kidiffundál.
- A szénsav H+ + HCO3- = H2CO3 reakcióból pótlódik.
- A H+ a hemoglobinhoz kötött H+-ból származik, így a hemoglobin újra oxigént vehet fel.
- A sejtben tehát fogy a HCO3-, ezért a vérplazmából mennyisége Cl- cserével pótlódik.
- a globin láncok N-terminálisához is kötődnek karboxil-csoportot alkotva. Ezen az úton jut a tüdőbe a CO2 mintegy 20%-a,
- ezenkívül a vérplazmában is oldódik – 10% -, és hidrokarbonát-ionok formájában szállítódik.
- a sejtek elsődleges energiaforrásai (pl. glükóz),
- tartaléktápanyagok (pl. glikogén),
- más vegyületekkel összekapcsolódva fontos makromolekulák – nukleinsavak – építőkövei (ribóz, dezoxiribóz).
- A GLUT-1 a legtöbb szövetben – pl. agy, vörösvértestek, vastagbél - kis mennyiségben található meg, inzulintól független biztosítja a glükóz felvételét.
- Az inzulinfüggő GLUT-4 többek között izom- és zsírszövetben fordul elő.
- vagy energiát szolgáltatva lebomlanak,
- vagy májsejtekben, ill. zsírsejtekben felépítő folymatokban vehetnek részt.
- a vérből szénhidrátokat – glükózt – vesz fel (magas vércukorszint esetén)
- és ad le (alacsony vércukorszint esetén) a szükségleteknek megfelelően.
- Továbbá szénhidráthiány esetén a máj nemszénhidrát előanyagokból (tejsav, glicerin, bizonyos aminosavak (pl. alanin és glicin)) glükózt tud előállítani kortizol hatására.
- Táplálkozást követően, amikor a vércukorszint megnő, a vérből felvett szénhidrátok
- a májban vagy lebomlanak energiát szolgáltatva,
- vagy átalakulnak pl. zsírokká,
- vagy átmenetileg raktározódnak glikogénként.
- Éhezéskor, vagy hosszan tartó izommunka esetén, ha a vércukorszint csökken, a raktározott glikogén lebontása útján a keletkezett glükózt a vérbe juttatja.
- Az inzulin - magas vércukorszint esetén - serkenti a máj glükózfelvételét, a glikogén, ill. zsírok szintézisét.
- Az adrenalin, glukagon – ill. szimpatikus aktivitás - hatására pedig fokozódik a máj glikogénbontása, a keletkező glükóz leadása.
- Egyetlen hormonként csökkenti a vércukorszintet, mivel segíti a szövetek, sejtek cukor felvételét, segíti a cukor bejutását a sejtekbe, (elsősorban a máj-, zsír-, izomszövetekre hat, az idegsejtek glükóz felvételét nem befolyásolja),
- serkenti a máj glikogén szintézisét, csökkenti a glükóz leadását, gátolja a glükoneogenezist,
- növeli az izom glikogén szintézisének mértékét,
- fokozza a szövetek - főleg a zsírszövet - cukor oxidációját, serkenti az ebből meginduló zsírszintézist, ami biztosítja, hogy a feleslegben felvett szénhidrátok (csoki) zsírok formájában raktározódjanak.
- Inzulin hatására az addig a sejt belsejében, hólyagocskákban tárolt GLUT-4 transzporterek beleolvadnak a külső membránba, s megnövelik a zsírsejtek és az izomrostok glükóz felvételét.
- a vizelet mennyisége is megnő,
- mivel a szűrlet koncentrációja a benne maradó cukortól magas,
- így kevesebb víz tud visszaszívódni,
- a betegek vízfogyasztása a vízvesztés következtében megnő.
- emeli a vércukorszintet, csökkenti a máj glükóz raktározását, serkenti a glikogén bontását (cAMP) fokozza a keletkezett glükóz leadását,
- a sejtekben – főleg az izomzatban - serkenti a glükóz oxidációját.
- emeli a vércukorszintet,
- serkenti az izomfehérjék bontását, a májba jutott aminosavak cukrokká alakítását (glükoneogenezis),
- a májba jutott tejsavból elősegíti a glükóz képződését (glükoneogenezis, Cori-kör).
- mert a szénhidrátok állatokban másodlagos tartalék tápanyagok,
- legfontosabb energiaszolgáltatók,
- a reakcióút köztes termékei - intermedierjei - több bioszintetikus útnak részei.
- a biológiai oxidáció,
- és az erjedés.
- a glikolízis,
- a citrátkör,
- a terminális oxidáció.
- A glükózlebomlás első szakasza.
- Nem kell hozzá oxigén, ezért
- az erjedési folyamatok részét is képezi.
- A sejtek citoplazmájában játszódik le.
- 2 molekula piroszőlősav (piruvát),
- nettó 2 molekula ATP,
- 2 molekula NADH + H+ (H-leadás miatt oxigén nélküli oxidáció történik!).
- Amennyiben a lebomlás anaerob körülmények között zajlik, a piroszőlősav az erjedés folyamatában alakul tovább (lásd később).
- Azonban, ha a disszimiláció oxigén mellett folyik, a reakcióút a citromsavciklusba
- érösszehúzódás
- vérlemezke fázis (kitapadás, összecsapódás, aktiválódás)
- véralvadási szakasz
- fibrinolízis
- a vérlemezkék kitapadnak a hámréteg alatti kötőszövet kollagén rostjaihoz, majd kifejlődő nyúlványaikkal összekapcsolódva elfedik a szabaddá vált kollagént.
- A kitapadás hatására a hámsejtekből olyan anyagok szabadulnak fel, melyek fokozzák a vérlemezkék további kitapadását, ill. nagymértékű összecsapódásra késztetik őket, melynek eredményeképpen egy vérlemezkékből álló tömítés, a fehér trombus jön létre.
- A kitapadás továbbá aktiválja a vérlemezkéket, aminek hatására különféle faktorokat (ADP, adrenalin, szerotonin) és tapadást segítő anyagokat bocsátanak ki.
- Fibrinogén
- Protrombin
- Szöveti faktor (tromboplasztin)
- Kalcium
- A trombin egy enzim, amely az oldott fibrinogénből lehasít egy oligopeptidet, létrehozva a polimerizálódó fibrint.
- Az oldhatatlan fibrinpolimer térhálós szerkezetű, amelybe beletapadnak a vér alakos elemei. Az így kialakult kocsonyás képződményt vérlepénynek vagy vörös trombusnak nevezzük, mely méretének növekedésével teljesen elzárja a nyílást.
- ér-eredetű rendellenesség,
- a vérlemezkékkel kapcsolatos problémák,
- valamint véralvadási faktorokkal összefüggő zavar miatt.
- Egyrészt képződésük elégtelenné válik (bizonyos gyógyszerek károsító hatása nyomán, vírusfertőzésben (pl.: parvovírus), általános csontvelő-elégtelenség részjelenségeként, pl. leukémiákban),
- másrészt a fokozott vérlemezke-pusztulás miatt (vírusfertőzésekben, gyógyszerhatás miatt, fokozott lépbeli raktározás következtében stb.).
- A vérlemezke-funkció zavart okoz pl. az aszpirinkezelés, amely gátolja a vérlemezkék kitapadását az érfalakhoz.
- hemofília A: VIII. faktor hiánya, ill.
- hemofília B: IX. faktor hiánya, az X-kromoszómához kötött öröklődő betegségek.
- A láb duzzanata,
- fájdalom az érintett lábban,
- az érintett terület melegsége.
- A véráramlás lelassulása. Tartós ágyhoz kötöttség, sérült vagy operált végtag rögzítése, jelentős visszértágulat, szívelégtelenség stb. kapcsán a véráramlás lassul, ami segíti a véralvadást.
- Az érfal legbelső rétegének sérülése, pl. visszérbetegség esetén, melynek kapcsán az érfalból véralvadási faktorok szabadulnak fel és a sérülés helyén a vérlemezkék összetapadnak.
- A vér alvadási készségének fokozódása miatt, pl. fogamzásgátló tabletták, dohányzás, veleszületett tényezők stb. kapcsán.
- elhízás,
- dohányzás,
- szívelégtelenség,
- sérülések,
- műtét utáni állapot, tartós fekvést igénylő súlyos betegségek, illetve a nagyon hosszas ülés (repülőutakon).
- a terhesség és szülés körüli időszak,
- fogamzásgátló tabletta hosszan tartó szedése.
- felül két pitvar,
- alul két kamra.
- A pitvarok és a kamrák között: a vitorlás billentyűk.
- A kamrák és az artériák között: a zsebes billentyűk.
- Az első egy tompa, mély hang (vitorlás billentyűk),
- a második magasabb hang (zsebes billentyűk).
- A szív elernyedt állapota – diastole
- Az összehúzódás állapota – sistole
- A pitvarok összehúzódnak,
- térfogatuk csökken,
- bennük a nyomás emelkedik, majd csökken,
- a vitorlásbillentyűk nyitottak, s a vér az ernyedt kamrákba áramlik. A pitvarok ürülését a táguló kamrák szívóhatása elősegíti.
- A kamrák vérrel teltek, a pitvarok felől jövő összehúzódás átterjed a kamrák falára, a kamrák megkezdik összehúzódásukat.
- A kamra fala ráfeszül a benne lévő vérre, a vérnyomás meredeken nő, s mikor eléri, ill. meghaladja a pitvari nyomást, a vitorlás billentyűk becsapódnak (első szívhang).
- A zsebes billentyűk még zárt állapotúak.
- A kamratérfogat változatlan, a vér nem áramlik semerre.
- A kamrák összehúzódása (szisztole) fokozódik,
- s mikor a növekvő vérnyomás eléri, ill. meghaladja a nagyartériákban mért nyomást (pl. az aortában 80 Hgmm-t), az eddig zárt zsebes billentyűk megnyílnak, s a kamrák vértartalmának kb. 60%-a az artériákba ömlik.
- A kamratérfogat csökken, a kiáramló vér miatt a nyomás elkezd csökkenni.
- A kamrák összehúzódásával párhuzamosan a pitvarok elernyednek, s kezdenek vérrel telítődni (tágulásuk szívóhatást gyakorol a nagyvénákra).
- A kamrák elernyednek, a vérnyomás meredeken csökken és amikor a kamrai nyomás az artériákban mért nyomás alá esik (aortában kb. 105 Hgmm), a zsebes billentyűk becsapódnak (második szívhang), a vitorlás billentyűk még zártak.
- A kamrák vértérfogata nem változik, a vér nem áramlik semerre.
- A kamra vértérfogata nő, vérnyomása tovább csökken.
- Amikor a nyomás a kamrákban a pitvari érték alá csökken, a zárt zsebes billentyűk mellett a vitorlás billentyűk megnyílnak, a vér passzívan beáramlik a kamrákba, a vérbeáramlás miatt a kamrai nyomás enyhén nő.
- A pitvarok ürülését elsősorban a kamrák térfogatnövekedése, ill., ami ezzel együtt jár a kamrai nyomás csökkenése teszi lehetővé, ami erős szívóhatást gyakorol a pitvarokra.
- a felnőtteknél 72,
- 12 éves kor körül kb. 80,
- újszülötteknek 140.
- Szinuszcsomó
- Pitvar-kamrai csomó
- His-köteg
- Tawara-szárak
- Purkinje-rostok
- His-köteg
- Tawara-szárak
- Purkinje-rostok viszik.
- P-hullám (pitvari hullám): az ingerület pitvari terjedésének felel meg.
- P- Q távolság: átvezetési idő a pitvar és kamra között.
- QRS-komplexum (kamrai hullám): a kamrák depolarizációját jelöli, ez idő alatt megy végbe a kamra teljes munkaizomzatának összehúzódása.
- ST-szakasz: a kamrák lassú repolarizációs szakasza.
- A pressor központnak önálló, állandó aktivitása van, érszűkítő hatású, növeli a vérnyomást és a szívfrekvenciát (szimpatikus hatás).
- A depressor központnak önálló aktivitása nincs, a receptorok felől aktiválódik szükség esetén. Gátolja a pressor kp-ot, a gerincvelői kp-okat, továbbá közvetlenül a szívet (paraszimpatikus hatás X. agyidegen keresztül).
- Inger: vérnyomás emelkedés.
- Receptor az aortaív falában,
- depressor kp. aktiválás, pressor kp. gátlás,
- értágulás, vérnyomás csökken,
- X. paraszimpatikus agyideg közvetlenül gátolja a szinuszcsomót,
- szívfrekvencia csökken,
- perctérfogat csökken.
- a genetikai hajlam,
- az életkor,
- a férfi nem,
- magas vérnyomás,
- a vérzsírok emelkedése (LDL),
- az elhízás,
- a dohányzás,
- a cukorbetegség,
- a mozgásszegény életmód,
- a stressz.
- nyomásérzés a mellkas közepén, esetleg csak furcsa teltségérzés, vagy szorító mellkasi fájdalom (angina pectoris), ami több mint néhány percen keresztül fennáll,
- kisugárzó fájdalom a mellkasból a váll, a kar, a hát vagy akár a fogak, állkapocs irányába,
- légszomj,
- izzadás,
- ájulás,
- hányinger, hányás,
- megsemmisülés érzés, halálfélelem,
- sápadtság, szürkeség.
- gyorsabb szívverést tachikardiának nevezzük a szívfrekvencia 100/percnél magasabb,
- míg a túl lassút bradikardiának hívjuk, a szívfrekvencia 60/perc alatt van.
- A szívből indulnak ki a verőerek vagy artériák.
- A szív felé vezetik a vért a gyűjtőerek vagy vénák.
- A legkisebb artériákat - arteriolákat - és a legkisebb vénákat - venulákat - a hajszálerek vagy kapillárisok kapcsolják össze.
- A nagyvérkör a test vérköre, a bal kamrából eredő aortával kezdődik, egyre kisebb erekre ágazva ellátja a test szöveteit, szerveit oxigéndús vérrel, majd a kapillárisok után egyre nagyobb vénákba szedődve a fő vénákkal, az alsó és a felső testvénával a szív jobb pitvaránál végződik.
- A kisvérkör a tüdő vérköre, a jobb kamrából az oxigénben szegény vért szállító tüdőartériával kezdődik, majd a tüdő kapillárisaiban az oxigénnel telítődött vért a tüdő vénái juttatják a bal pitvarba.
- külső réteg (kötőszövet),
- középső réteg (simaizomszövet),
- belső réteg (egyrétegű laphám).
- A külső réteg főleg rugalmas rostokat tartalmazó lazarostos kötőszöveti réteg, amely a nagyobb artériák esetén tartalmazhat ereket és zsírszövetet.
- A középső réteg simaizomból épül fel.
- A belső réteg egyrétegű laphámból áll.
- Az artéria-véna kapillárisok általában ív alakban, rövid úton kötik össze az arteriolákat a venulákkal.
- A valódi kapillárisok az artéria-véna kapillárisok különböző pontjait kötik össze bonyolult hálózatot alkotva.
- Általában az intenzíven működő, élénk anyagcseréjű szövetekben a vér a valódi kapillárisok felé áramlik.
- Nyugalomban levő szövetekben az izomgyűrűk zártak, a vér az artéria-véna kapillárisokon keresztül rövid úton távozik.
- Magasnyomású rendszerre
- Alacsony nyomású rendszerre
- Amikor a pitvar szisztole, ill. az egyidejű kamra diasztole következtében a pitvarban a vér nyomása legalacsonyabb kamrai nyomás - kb. 5 Hgmm - fölé megy, a vitorlás billentyűkön keresztül a vér a bal kamrába áramlik.
- A meginduló kamra szisztolének köszönhetően a nyomás meredeken emelkedik, majd 80 Hgmm-es nyomás értéknél megnyíló zsebes billentyűn keresztül a kamra tartalmának mintegy 60%-át az aortába juttatja. Ezt a kb. 70-80 ml vér a vérmennyiséget pulzustérfogatnak nevezzük.
- A zsebes billentyű megnyílása után a kamra és az aorta nyomása együtt nő, elérve a max. 120 Hgmm-t.
- Az aortában mért legkisebb nyomás érték ugyanakkor kb. 80 Hgmm, mivel mikor a kamara diasztolé következtében zuhanó kamrai nyomás értéke az aortai nyomás alá esik a zsebes billentyűk becsapódása – kb. 110 Hgmm-es értéknél - megakadályozza a vér visszafolyását.
- legnagyobb nyomást – 120 Hgmm - szisztolés nyomásnak hívjuk, amely a bal kamra összehúzódásának eredményeképpen alakul ki,
- a legkisebb nyomást – 80 Hgmm - diasztolés nyomásnak nevezzük, ami a bal kamra elernyedése következtében jön létre.
- A legnagyobb nyomásesés az artériás rendszerben az arteriolák területén történik, így ez a szakasz már az alacsony nyomású rendszer kezdete.
- A kapillárisok területén további 20 Hgmm-es nyomásesés következik be, 35-15 Hgmm nyomásérték között.
- A vénás rendszer elején mérhető 15 Hgmm-es nyomás érték a jobb pitvar közelében szinte 0 értéket mutat.
- Az egész kisvérkör az alacsony nyomású rendszerhez tartozik, sokkal kisebb nyomásingadozásokkal. Pl. a jobb kamra szisztolés nyomása csupán 30 Hgmm.
- az artériás rendszerben a szívtől távolodva egyre nő.
- Az arteriolák területén az oszlások során az összkeresztmetszet jelentősen nő,
- míg a legnagyobb értéket a kapillárisok területén észleljük.
- A vénák összkeresztmetszete a szív irányába fokozatosan csökken,
- a vérnyomásesés a kamráktól a pitvarokig, melynek fő oka a vénák falának tágulékonysága,
- a végtagok vénáinak billentyűi, melyek a gravitáció ellenében megakadályozzák a vér visszaáramlását,
- az izompumpa,
- a negatív mellűri nyomás, amelynek során a mellkasban belégzéskor kialakuló alacsony nyomás szívóhatást gyakorol a gyűjtőerekre, segítve a vénás visszaáramlást.
- Bizonyos szervek - agy - vérellátása egyenletesnek mondható,
- ugyanakkor másoké - pl. vázizmok, bőr, zsigerek - a szervezet mindenkori állapotának megfelelően széles határok között változhat.
- egyrészt a kevésbé működő szervek vérellátása csökken,
- másrészt a vénás rendszer tónusának fokozódására jelentős mennyiségű vér kerül át az artériás rendszerbe.
- Intravazális (vér)
- Intersticiális (szövetnedv, nyirok)
- Intracelluláris (sejten belüli tér)
- a szövetek alapállományát képezi,
- közeget biztosít a sejtek számára,
- a vérből szűrődik ki,
- közvetít a vér és a sejtek között,
- felesleges részét a nyirokkeringés szállítja vissza a vérbe, mint nyirokfolyadékot.
- átbocsátja a vizet, az ionokat, kisebb szerves molekulákat, pl. glükózt, aminosavakat,
- azonban visszatartja a makromolekulákat és az alakos elemeket.
- Tehát a kapillárisok artériás szakaszán nettó 10 Hgmm-es nyomás a vérplazmát kifelé préseli.
- A kapillárisokban a vénás oldal felé haladva a folyadék kilépése következtében a vér vér nyomása lecsökken.
- Amikor a vérnyomás egyenlővé válik a kolloidozmotikus nyomással, a folyadék kiszűrődése megszűnik.
- A kapillárisok vénás szakaszán a vérnyomás tovább csökken kb. 15 Hgmm-re és a kolloidozmotikus nyomás alá esik. Ennek következtében a vénás szakaszon a vérfehérjék kolloidozmotikus szívóhatásának köszönhetően a szövetnedv egy része visszalép az érpályába, elszállítva a szövetekben keletkezett bomlásterméket.
- a felesleges szövetnedv visszajuttatása az érpályába,
- zsírok szállítása,
- immunsejtek termelése, tárolása, szállítása.
- Nő a vénás rendszerben a nyomás, ekkor több folyadék lép ki, ill. kevesebb lép vissza. Ez történik az alsó végtagokban, ha a szív teljesítménye csökken (idősebb korban), továbbá terhesség, ill. trombózis miatt.
- Csökken a plazmafehérjék koncentrációja - éhezéskor -, így csökken a kolloidozmotikus szívóerő.
- Vizenyő alakulhat ki akkor is, ha a nyirokerek eltömődése miatt azok elvezetése csökken.
- Az hajszálerek fala átjárhatóságának megnövekedése - gyulladás miatt - szintén a szövetnedv felszaporodását eredményezi.
- a nyirokérrendszer két végpontja közötti nyomáskülönbség határozza meg,
- továbbá a nyirokerekben található zsebes billentyűk, melyek az izmok összehúzódásakor fellépő izompumpa következtében összepréselődő nyirokerekből a nyirkot a szív felé irányítják.
- Nyirokerek
- Nyirokszervek
- nyirokcsomók
- mandulák, féregnyúlvány (nyiroktüszők)
- lép
- csecsemőmirigy, vörös csontvelő
- Összekapcsolják a nyirokereket.
- Megszűrik, a különféle sejttörmelékektől megtisztítják a nyirkot.
- Nyiroksejteket tárolnak.
- Vérraktározás.
- Kiszűri és eltávolítja az elöregedett vörösvértesteket.
- Nyiroktüszőinek köszönhetően nyiroksejteket raktároz és termel.
- elsősorban ozmoreguláció, azaz a szervezet optimális ozmotikus koncentrációjának a fenntartása,
- másodsorban méregtelenítés.
- a feleslegben felvett és a szervezet belső egyensúlyát veszélyeztető (víz, sók stb.),
- valamint az anyagcsere során feleslegessé vált (NH3, karbamid, húgysav, tejsav, bilirubin stb.), nagyobb mennyiségben mérgező anyagokat távolítja el a szervezetből.
- a tüdő, mivel közreműködik az anyagcserében keletkezett szén-dioxid eltávolításában.
- A máj szintén kiválaszt, amikor a hemoglobin bomlásából származó bilirubint az epével a bélcsatornába üríti.
- A bőr verejtékmirigyeinek működése révén a víz, konyhasó, illatanyagok és egyes gyógyszerek kiválasztását végzi.
- A tápcsatorna – végbél – elsősorban a sókiválasztásban működhet közre.
- ozmotikus koncentráció,
- ionösszetétel,
- testfolyadékok kémhatása,
- testfolyadékok térfogata.
- a páros vesék,
- a páros húgyvezetékek,
- a páratlan húgyhólyag,
- a páratlan húgycső.
- Nemkívánatos anyagok eltávolítása:
- anyagcsere-végtermékek,
- feleslegben felvett anyagok.
- Benne termelődő hormonok révén
- a vérnyomás szabályozása: renin,
- vörösvértest-képződés serkentése: eritropoetin.
- H+-kiválasztás miatt a pH-szabályozás, sav-bázis egyensúly biztosítása.
- Vitaminszintézis: a D-vitamin aktív formájának előállítása.
- a kb. 2,5 cm széles, külső tömör állomány
- egy nagyobb központi üreget, a vesemedencét veszi körül.
- perifériásan a szemcsézett kéregállományt és
- az üreg felé eső sugaras lefutású velőállományt különböztethetjük meg.
- a vesetestecskére és
- az elvezető csatornára tagolható.
- hajszálérhálózatból – érgomolyag – és az azt körülvevő
- kettős falú Bowman-tokból áll.
- a közeli kanyarulatos csatornára (proximális tubulus),
- a Henle-kacsra és
- a távoli kanyarulatos csatornára (disztális tubulus).
- a szűrőműködés,
- a visszaszívó működés és
- a kiválasztó működés eredményeként jön létre.
- vizet, sókat,
- tápanyagokat – glükózt, aminosavakat stb. –,
- bomlástermékeket – karbamidot, urobilinogént, húgysavat stb. –,
- hormonokat – szteránvázas ivari hormonokat, kisebb peptidhormonokat, mint pl. hCG-t, a terhességi hormont.
- a glükóz, aminosavak, sók aktív transzporttal,
- a víz pedig passzív transzporttal, ozmózissal.
- méretüknél fogva nem szűrődnek át a vérből, mint pl. egyes gyógyszerek,
- vagy nagyobb mennyiségük eltávolítása szükséges, pl. H+ (vér pH-szabályozása).
- A vérnyomás (Pv) az érpályából kifelé préseli a vérplazmát,
- ennek ellene hat a vérplazmafehérjék kolloidozmotikus nyomásából (Pcoll) adódó szívóerő
- és a Bowman-tok üregében található szűrlet nyomása (PB).
- Na⁺ + Cl⁻ + HCO₃⁻ stb.→ kb. 280–290 mOsm
- Fehérjék hozzájárulása:→ ~1–2 mOsm
- Belépő onkotikus nyomás: ~25 Hgmm
- Kilépő onkotikus nyomás: ~35 Hgmm
- 100%-a
- a proximális tubulusban szívódik vissza,
- aktív transzporttal (Na+-glükóz kotranszport),
- A sók – elsősorban a NaCl, mivel ez az ion teszi ki a szűrlet ionjainak 90%-át – elsősorban aktív transzporttal,
- a víz az oldott anyagok mozgását követve vízcsatornákon (aquaporinokon) keresztül passzív transzporttal – ozmózissal – szívódik vissza.
- az átszűrődött víz további 15%-a szívódik vissza,
- ugyanakkor a csatorna fala a sók számára szinte átjárhatatlan.
- a víz számára átjárhatatlan,
- míg a sók aktív transzporttal kilépnek a csöveket körülvevő szövet közötti folyadékba, létrehozva e tér magas ozmotikus koncentrációját.
- a felszálló ágból kilépett sók növelik a csövek közötti tér (szövetnedv) koncentrációját (2),
- mivel a leszálló ágból víz szívódik vissza, a szűrlet NaCl-koncentrációja emelkedik (3),
- mire a felszálló ág kezdetéhez kerül (4), ebből a magasabb NaCl-koncentrációjú szűrletből jut ki a NaCl aktív transzporttal a szövetnedvbe tovább emelve annak töménységét. (5)
- A töményebb szöveti tér hatására fokozódik a leszálló ágban a víz kilépése, ami tovább növeli a leszálló ág koncentrációját (6) és így tovább.
- fokozódik az ADH felszabadulása,
- melynek hatására a gyűjtőcsövek fala áteresztővé válik a víz számára, így az passzív transzporttal a csatornákat körülvevő nagyobb koncentrációjú szövetnedvbe, ill. a vérplazmába áramlik.
- Ennek eredményeképpen a vizelet mennyisége csökken, koncentrációja nő.
- vizet, ionokat (Na+, K+, Ca2+, CI-, NH4+, PO43- ),
- karbamidot (az aminosavak N-tartalmú bomlásterméke),
- húgysavat (a purinbázisok N-tartalmú bomlásterméke),
- urobilinogént (a hemoglobin bomlásterméke),
- tejsavat (megerőltető izommunka esetén),
- szteroidhormonokat,
- terhesség esetén a hCG terhességi hormont, gyógyszereket, méreganyagokat.
- a táplálék és az elfogyasztott folyadék minősége és mennyisége,
- hőmérséklet, fizikai aktivitás,
- különféle élettani állapotok, mint pl. terhesség.
- Amennyiben a hipotalamuszban csökken vagy megszűnik a víz visszaszívását serkentő ADH (vazopresszin) hormon termelődése, akár napi 15 liter híg vizelet is keletkezhet, folyamatos gyötrő szomjúság mellett.
- Inzulin hiányában – I. típusú cukorbetegség esetén - is megnő a vizelet mennyisége, mivel a szűrlet koncentrációja a benne maradó cukortól magas lesz, így az ozmotikus egyensúly eltolódása miatt kevesebb víz tud visszaszívódni a vesében.
- környezet különféle vegyületei hatnak a receptorokra – kemoreceptorok az ingerfelvevők -,
- ősiek,
- informálják a szervezetet a táplálékról, annak potenciális kedvező és kedvezőtlen hatásairól (mérgezés),
- olyan fiziológiás változásokat is kiváltanak a szervezetben, melyek az emésztéshez kellenek, pl. megindul az emésztőnedvek elválasztása, fokozódik a perisztaltika,
- a magatartásbeli, hangulatbeli változást is okoznak (undor),
- nagy a felbontóképességük (több ezer szag megkülönböztetése),
- alacsony az ingerküszöbük,
- könnyen adaptálódnak.
- Limbikus rendszerrel való szoros kapcsolat miatt régi emlékek felidézője lehet.
- A szagok meghatározásában nagy egyéni különbségek tapasztalhatók.
- A nők érzékenyebbek.
- Az életkorral változik, 20-as évek közepétől a 40-es évek végéig a legnagyobb az érzékenység.
- Dohányosoknál, gőzölgő vegyi anyagokkal dolgozóknál, nagyvárosokban lakóknál csökkent érzékenység, vakoknál nagyobb érzékenység alakul ki.
- A szaglóhám az orrüreg felső részén helyezkedik el,
- az itt található receptorok – primer receptorok - saját axonnal rendelkező valódi idegsejtek.
- A receptorok között támasztósejtek vannak.
- Kb. 1000 különböző szaglóreceptort ismerünk, melyek a nyálkahártyában oldott szaganyagokat, nyúlványaikkal érzékelik.
- A receptorok axonjai átkapcsolás – szaglógumóban, Bulbus olfactoriusban - után a halántéklebeny és a homloklebeny alsó felszínén található, igen ősi szaglókérgéhez futnak, itt tudatosul a szaglásérzet. A talamusz szintén kap szaglórostokat.
- A szaglópálya kapcsolatban van más kéregalatti struktúrákkal is, pl.: az érzelmekért és a hosszú távú memóriáért felelős limbikus rendszerrel (hippocampusz, amigdala), aminek köszönhetően különféle magatartási reakciókat, érzelmeket váltanak ki, emlékeket idéznek fel az egyes szagingerek. Ez a rendszer felelős a szagok viselkedésünkre gyakorolt tudat alatti befolyásoló hatásáért.
- A szaglóhagyma idegsejtjeinek axonjai képezik a szaglóideget, az I. agyideget, amely nem valódi agyideg.
- A szaglórendszer gyorsan adaptálódó rendszer, amely elsősorban a szaglóközpont működési sajátossága.
- A nyelv hátának elülső 2/3-án szemölcsök találhatók.
- Egyes szemölcsökben ízérző sejtcsoportok, ún. ízlelőbimbók vannak, amelyekben különböző anyagokra érzékeny kemoreceptorok
- A savanyú íz érzete a H+-koncentráció függvénye,
- a sós íz a kisebb pozitív töltésű kationok hatására (Na+) alakul ki,
- míg ugyanezek az ionok nagy koncentrációban keserű ízt okoznak, de a nagyobb töltésű kationok, pl. Ca2+, Mg2+ szintén keserűek,
- édes ízt a legkülönfélébb szerkezetű vegyületek okozhatnak, de természetes módon a sok -OH csoportot tartalmazó vegyületek felelősek az édes íz kialakításáért.
- Az ötödik alapíz az umami – ami japánul finom ízt jelent - előfordul pl. a paradicsomban, a parmezán sajtban és a japán konyhában használt kombu nevű barnamoszatban. Ezt az ízt kiváltó mesterséges adalékanyag a nátrium-glutamát, E 621.
- A csípős ételekben lévő anyagok, pl. a kapszaicin a szájüreg fájdalomérző receptoraira hatnak.
- Csecsemőknél viszonylag kevés receptor található, ezután számuk gyorsan növekszik, 40. életév tájékán maximális, később mennyiségük csökken.
- Az ízlelőbimbók állandóan pusztulnak és újak lépnek a helyükbe, átlagéletkoruk kb. 10 nap.
- A kemoreceptorok módosult hámsejtek – érzékhám -, nem igazi idegsejtek, nincs saját axonjuk.
- A receptorokból az ingerületet bipoláris neuronok vezetik el, amely a nyúltvelőn, a talamuszon keresztül a fali lebeny alsó részébe jut, ahol tudatosul az ízérzet. Természetesen egyes rostok a limbikus rendszerbe futnak, aminek köszönhetően a különféle ízek hatást gyakorolnak az érzelmeinkre, viselkedésünkre, emlékekkel kapcsolódhatnak össze, undort válthatnak ki.
- kémiai – a vér és az idegszövet sejtközötti állományának CO2 és O2 tartalma – és
- a tüdőből származó mechanikai ingerek játszanak szerepet.
- részben a szívhez közeli erek falában található kemoreceptorok,
- részben maguk a nyúltvelői szabályozó központok képesek felfogni.
- a testfolyadékok - vérplazma - ozmotikus koncentrációja (ozmotikus nyomása) növekszik (hiperozmózis),
- ugyanakkor térfogata csökken (hipovolémia).
- A pressor központ érszűkítő hatású, növeli a vérnyomást és a szívfrekvenciát (szimpatikus hatás).
- A depressor központ gátolja a pressor kp-ot, a gerincvelői kp-okat, továbbá közvetlenül a szívet (paraszimpatikus hatás X. agyidegen keresztül).
-
- helyük: szívhez közeli erek falában,
- inger: O2 hiány (CO2 növekedés),
- hatás: pressor kp. aktiválás, vérnyomás nő, szívfrekvencia nő.
- A nyúltvelőben, maga a pressor központ neuronjai.
- Inger a vér CO2 növekedése (a liquor H+-koncentrációjának növekedése).
- Hatás: pressor kp. aktiválás, vérnyomás nő, szívfrekvencia nő.
- az éhségközpont, amelynek aktivitása éhségérzetet kelt és ez a táplálék felvételét eredményezi, ill. ugyanitt van
- az ellentétes hatású jóllakottsági központ.
- alacsony vércukorszint
- és olyan peptidhormonok – pl. ghrelin szöveti hormon – váltják ki, amiket a gyomor üressége esetén elsősorban a tápcsatorna fala választ el.
- a magas vércukorszint és inzulinszint, továbbá
- a zsírszövet sejtjeiben termelődő leptin szöveti hormon.
- Morfogenetikai hatás
- Az anyagcserére tett hatás
- Csökkenti a máj glükóz raktározását,
- emeli a vércukorszintet,
- serkenti a sejtek cukor oxidációját.
- alacsony alapanyagcsere,
- alacsony testhőmérséklet,
- lelassult ideg- és izomműködés.
- az alapanyagcsere szintje megnő, nő az oxigénfogyasztás,
- a testhőmérséklet emelkedik,
- a testsúly annak ellenére csökken, hogy a beteg sokat eszik,
- ideg- és izomrendszer túlműködik, emiatt jellemző a fáradékonyság, remegés, izzadás.
- Csökkenti a máj glükóz raktározását,
- emeli a vércukorszintet,
- serkenti a sejtek cukoroxidációját.
- emeli a vércukorszintet,
- serkenti az izomfehérjék bontását, a májba jutott aminosavak cukrokká alakítását (glükoneogenezis),
- a májba jutott tejsavból elősegíti a glükóz képződését (glükoneogenezis, Cori-kör).
- emeli a vércukorszintet,
- csökkenti a máj glükóz raktározását,
- serkenti a glikogén bontását (cAMP) fokozza a keletkezett glükóz leadását,
- a sejtekben – főleg az izomzatban - serkenti a glükóz oxidációját.
- Egyetlen hormonként csökkenti a vércukorszintet, mivel segíti a szövetek, sejtek cukor felvételét, segíti a cukor bejutását a sejtekbe, (elsősorban a máj-, zsír-, izomszövetekre hat, az idegsejtek glükóz felvételét nem befolyásolja),
- serkenti a máj glikogén szintézisét, csökkenti a glükóz leadását, gátolja a glükoneogenezist,
- növeli az izom glikogén szintézisének mértékét,
- fokozza a szövetek - főleg a zsírszövet - cukor oxidációját, serkenti az ebből meginduló zsírszintézist, ami biztosítja, hogy a feleslegben felvett szénhidrátok (csoki) zsírok formájában raktározódjanak.
- Inzulin hatására az addig a sejt belsejében, hólyagocskákban tárolt GLUT-4 transzporterek beleolvadnak a külső membránba, s megnövelik a zsírsejtek és az izomrostok glükóz felvételét.
- Táplálkozást követően, amikor a vércukorszint megnő, a vérből felvett szénhidrátok a májban
- vagy lebomlanak energiát szolgáltatva,
- vagy átalakulnak zsírokká,
- vagy átmenetileg raktározódnak glikogénként.
- Éhezéskor, vagy hosszan tartó izommunka esetén, ha a vércukorszint csökken, a raktározott glikogén lebontása útján a keletkezett glükózt a máj a vérbe juttatja.
- Az inzulin - magas vércukorszint esetén - serkenti a máj glükóz felvételét, a glikogén, ill. zsírok szintézisét.
- Az glukagon hatására pedig fokozódik a máj glikogén bontása, a keletkező glükóz leadása, ami a vércukorszint emelkedését eredményezi. Továbbá fokozza a glükoneogenezist és a zsírok bontását. Összeségében hatása az inzulinéval ellentétes.
- 1-es típusú cukorbetegség: a betegség oka a hasnyálmirigy Langerhans-szigeteiben az inzulintermelő béta-sejtek pusztulása. Ez a szigetsejtek elleni autoimmun-reakció következménye. Bármely életkorban kialakulhat. A betegség csak inzulinpótlással kezelhető. Gyermekkori diabetes elsődleges oka.
- 2-es típusú cukorbetegség: a csökkent inzulinérzékenység – ún. inzulinrezisztencia - következtében alakulhat ki. Az inzulinrezisztencia az a jelenség, amikor a sejtek nem érzékelik az inzulin jelenlétét, aminek hátterében az inzulinreceptorok számának csökkenése, ill. szerkezetük megváltozása áll. A betegség kezdetén még vércukoreltérés nem jelentkezik, a hasnyálmirigy fokozott inzulintermelése miatt. A betegség előrehaladtával a hasnyálmirigy inzulin-elválasztása a béta-sejtek kimerülése miatt elégtelenné válik és emiatt kialakulnak az 1-es típusú diabetes tünetei. Kialakulásának hátterében örökletes hajlam, túlsúly, inaktív fizikai életmód, egészségtelen étrend (magas cukor- és szénhidráttartalommal) áll.
- Másodlagos cukorbetegség: kialakulhat alkoholizmus vagy epeúti kövesség következtében.
- A terhességi cukorbetegség: oka, hogy a terhesség alatt a méhlepényben termelődő hormonok csökkenthetik az anyai szervezetben termelődő inzulin hatását.
- a nagy mennyiségű édes vizelet (mézédes húgyár), tehát cukorürítés,
- a vízvesztés miatt kialakuló szomjúságérzet, sok folyadék fogyasztása,
- acetonos lehelet,
- gyakori eszméletvesztés,
- testsúlycsökkenés.
- magas a vércukorszint, a cukor a szövetekből vizet vonva el a vizelettel távozik, (a vizelet mennyisége megnő, mivel a szűrlet koncentrációja a benne maradó cukortól magas, így kevesebb víz tud visszaszívódni, a betegek vízfogyasztása a vízvesztés következtében megnő),
- a sejtek glükóz hiányában fokozott zsírbontásra kapcsolnak, aminek következtében számos a vér pH-ját savas irányba mozdító anyagcseretermék ún. ketontestek halmozódnak fel, mint pl. acetecetsav, aceton stb. Ennek eredménye az acidózis, ami eszméletvesztést, súlyosabb esetben kómát okoz.
- a fertőzésekkel szembeni csökkent ellenállóképesség,
- elhúzódó sebgyógyulás, sportsérülések valószínűsége megnő,
- érelmeszesedés kockázatának növekedése várható.
- A betegek látása romlik, a retinán vérömlenyek alakulnak ki.
- A vese kisereinek károsodása, mely fehérjevizelést, magas vérnyomást, végül veseelégtelenséget eredményezhet.
- Érzéskiesések, fájdalom, vegetatív működési zavarok alakulnak ki. A vegetatív zavarok közé tartozik az impotencia, hasmenés vagy székrekedés, fokozott izzadás, szapora szívverés.
- elhízás,
- sok cukor fogyasztása,
- mozgásszegény életmód,
- stressz,
- alkoholizmus.
- a csontok szervetlen alapállományának egyik alkotója (Ca5(PO4)3 X, X=OH-, F-),
- elengedhetetlen az izomműködésben,
- nélkülözhetetlen a véralvadásban,
- lehetővé teszi egyes enzimek működését,
- fontos az idesejtek aktivitásában, az ingerküszöböt emeli, fokozza az izom-ideg ingerlékenységet, hiánya izomgörcsökhöz vezet,
- szerepet játszik a szívizom ingerlékenységében,
- idegsejtekben citoplazmatikus koncentrációjának emelkedése fokozza a szinaptikus hólyagok exocitózisát,
- másodlagos hírvivő.
- a csontrendszer leadással és felvétellel egyaránt,
- a vesék szükség esetén visszaszívással,
- a bélcsatorna a táplálék Ca2+-tartalmának felszívásával D-vitamin jelenlétében.
- a kalcitonin, amely a pajzsmirigy kötőszövetében,
- a parathormon, amely a mellékpajzsmirigyben termelődik,
- kalcitriol, a D-vitamin aktív alakja, mely inaktív alakjából a vesében képződik.
- a csontok Ca2+ leadását serkenti (fokozza a csontfaló sejtek aktivitását),
- a vesében a Ca2+ kiválasztását csökkenti, visszaszívódását serkenti,
- a bélcsőben a Ca2+ felszívódását serkenti, melyhez a D-vitamin nélkülözhetetlen.
- A mellékpajzsmirigyek eltávolítása azonnal a vér Ca2+-tartalmának 30-40%-os csökkenéséhez vezet, ami súlyos izomgörcsöket, súlyosabb esetben fulladást eredményez,
- azonban a parathormon túltermelődése csontritkulást, csonttörékenységet okoz.
- A gerincvelő az idegrendszer ősi része, a csigolyaívek alkotta gerinccsatornában helyezkedik el.
- Kisujjnyi vastagságú, fehér, hengeres szerv, agyhártyák borítják.
- Nyaki, háti, ágyéki, keresztcsonti szakaszokra osztjuk.
- 0,5 m hosszú, az öreglyuktól kezdődően az első ágyékcsigolya magasságáig húzódik, mivel hossznövekedése nem tart lépést a gerinc növekedésével (lásd kép).
- Ettől lefelé a gerinccsatornát már csak a gerincvelőből ki- és belépő idegrostok kötegei – gerincvelői gyökerek - töltik ki (lófarok).
- A felső, ill. az alsó végtagot ellátó idegek kilépési helyei:
- a nyaki szakaszból kilépő karfonat,
- az ágyéki-keresztcsonti szakaszt elhagyó lábfonat.
- Kívül fehérállomány,
- belül szürkeállomány.
- Pillangó alakú.
- Neuronok sejttestei alkotják.
- Közepén van a központi csatorna.
- Hátsó szarv: interneuronok.
- Oldalsó szarv: központi vegetatív interneuronok.
- Mellső szarv: szomatikus mozgatóneuronok.
- Pályák alkotják.
- Elülső köteg: leszálló – mozgató - pályák.
- Hátulsó köteg: felszálló – érző - pályák.
- Oldalsó köteg: vegyesen.
- Közvetíti az agy utasításait a periféria felé és vissza, felszálló érzőpályákat és leszálló mozgatópályákat tartalmaz.
- Reflexközpont, önálló működése van.
- Izomeredetű.
- Bőreredetű.
- Szimpatikus
- Paraszimpatikus
- amely a combfeszítő izomban található,
- az izomrostokra csavarodott idegvégződés,
- mechanoreceptor, mivel adekvát ingere az izom megnyúlása (inakban az ínorsó van).
- Védi az izmot a túlzott passzív megnyúlástól, így az izomszakadástól, hiszen nyújtásra az izom összehúzódással reagál.
- Jelentős a testtartásban, azaz az antigravitációs izmok optimális feszítettségi állapotának beállításában, ezáltal testtartási reflexek alapját képezi.
- Alapvető az izomtónus fenntartásában (izomtónus: az izmok állandó kismértékű feszülése, a testtartásban alapvető a jelentősége).
- A fájdalomérző receptorok a bőrben vannak,
- a hátsó gyökérben levő csigolyaközti dúcban található az érzőidegsejt sejtestje (1),
- a gerincvelő hátsó szarvában ülő interneuronok kapcsolják össze az érző- és a mozgatóidegsejteket (2, 4),
- mellső szarvi, azonos oldali mozgatóneuronok működése kiváltja a hajlítóizmok összehúzódását, a veszélyeztetett végtag elhajlítását a veszélyforrástól (3),
- ugyanakkor gátló neuronoknak köszönhetően ugyanezen a végtagon a feszítőizmok elernyednek.
- Ezzel egyidejűleg, átkereszteződve az ellenkező oldali végtagon a mozgatóneuronok ingerülete a feszítőizmok összehúzódását eredményezik, ami lehetővé teszi az egyensúlyi helyzet megtartását (5).
- Az érzőidegsejt egyik elágazása egy felszállópályán az agyba küldi az érzőinformációt.
- védelem a károsító (fájdalmas) ingerek ellen,
- fontos testtartási reflexek alapja.
- A receptorok a zsigerekben – pl. bélfalban – találhatók,
- A hátsó gyökér csigolyaközti dúcában van az érzőneuron,
- a hátsó szarvban az interneuron,
- az oldalsó szarvban van a központi vegetatív neuron, melynek axonja
- elülső gyökéren keresztül lép ki, majd
- a perifériális vegetatív dúcban levő mozgatóneuronnal képez szinapszist,
- aminek a nyúlványa a végrehajtó belső szervre - mirigyek, erek, bél, bőr, nemi szervek simaizmaira - hat.
- lokális hideghatás esetén a bőr ereinek reflexes szabályozása,
- vizelet- és székletürítés szabályozása,
- szexuális reflexek, erekció, ejakuláció,
- béltartalom nyomásának hatására a bélmozgások szabályozása.
- nem diffundáló, nagy méretű anionokat tartalmaz, pl. fehérjéket, RNS-eket,
- magas a K+-koncentráció,
- alacsony a Na+-koncentráció,
- alacsony a Cl--koncentráció.
- magas a Na+-koncentráció,
- alacsony a K+-koncentráció,
- magas a Cl--koncentráció.
- Magas Ca2+-koncentráció, melynek főleg a szívizomban van jelentősége.
- Hipopolarizáció esetén a membránpotenciál értéke csökken (pl. –50 mV), pl. Na+-beáramlás miatt a serkentő szinapszisokban.
- Depolarizációkor a membrán eredeti polarizációja megszűnik.
- Hiperpolarizáció során a gátló szinapszisokban membránpotenciál értéke megnő, a potenciálváltozást a K+-kiáramlása, ill. a Cl--beáramlása okozza (-100 mV).
- Eltávolítja a sejt belsejéből az enzimműködést gátló Na+-t.
- Szabályozza a sejt ozmotikus nyomását (ellensúlyozza a sejtből kijutni nem tudó fehérje anionok ozmotikus szívóerejét, 3:2 ioncsere-aránnyal eléri, hogy a sejt belsejében kevesebb ozmotikusan aktív részecske legyen), ezáltal biztosítja a sejt optimális alakját.
- A pumpa blokkolásakor a sejt vizet vesz fel, duzzadni kezd.
- kismértékben állandóan nyitva lévő, ún. szivárgási csatornák,
- ligandkötődésre nyíló, adott feszültségértéknél áteresztő csatornák (lásd később),
- mechanikai ingerre nyíló csatornák (mechanoszenzitív ioncsatornák), melyek nyitási ingere a sejthártya deformálódása (mechanoreceptorokon).
- Az egyes ionoknak külön csatornája van, ismertek Na+-, K+-, Cl-- csatornák, melyek zárhatók és nyithatók.
- Endogén módon a sejtek anyagcseréjének változása okán. Ilyenkor külső inger hiányában is megváltozik az ionáramok erőssége. Ezek az oszcilláló (periódikusan változó) membránpotenciálú sejtek automatikus működésűek, endogén ritmusúak, pl.: szívizomsejtek (szinuszcsomó), egyes idegsejtek, simaizomsejtek. Ezeknek a sejteknek a működése az alapja a biológiai időmérésnek, a biológiai órák működésének, az élettani ciklusok – alvás-ébrenléti ciklus, női nemi ciklus, szívciklus (pl. a szív nyugalomban egy perc alatt 72-szer húzódik össze) stb. szabályozásának.
- A nyugalmi potenciál megváltozhat külső inger hatására is.
- Küzdelem esetén endogén opiátok termelődnek, melyek ingerküszöbemelő hatásuk révén csökkentik a fájdalomérzékenységet a sérülések iránt.
- Ha pl. alacsony a testfolyadékok – vér - kalciumszintje, csökken az izomrostok membránjának ingerküszöbe, s így fokozódik az ideg-izom ingerlékenység, ami miatt a vázizmok görcsösen összehúzódnak.
- Hipopolarizáció (2)
- Depolarizáció (3) (Na+- vagy/és Ca2+-beáramlás)
- Csúcspotenciál
- Repolarizáció (4) (K+-kiáramlás)
- Hiperpolarizáció (5) (K+-kiáramlás)
- Y tengely: membránpotenciál értéke mV-ban.
- X tengely: idő ms-ban mérve.
- Inger: a serkentő neurotranszmitter – ligand - megkötődik a fogadósejt receptorain.
- A receptorok ún. ligand-függő Na+-csatornák - melyek a sejttesten és a dendriteken találhatók meg - a kötődés hatására megnyílnak.
- Az ECF felől Na+-ok passzív transzporttal áramolnak be a sejtbe.
- A fogadó sejt membránjának (nyugalmi) potenciálja (a szinapszis körül) csökkenni kezd, a membrán hipopolarizálódik, ami a pozitív töltésű nátriumionok és/vagy kalciumion fokozódó beáramlásával áll kapcsolatban.
- Ekkor a sejttesten vagy a dendriten helyi/lokális potenciálváltozás alakul ki, melynek értéke/amplitúdója arányos az inger erősségével (a felszabaduló átvivőanyag mennyiségével).
- Amennyiben a helyi potenciál nagysága nem éri el az ingerküszöböt, a membránpotenciál mértéke újra növekszik, visszaáll a nyugalmi potenciál, mert helyreáll az eredeti ioneloszlás.
- Ugyanakkor a sejttesten egyidejűleg több helyi potenciál alakulhat ki, azok hatása az axoneredésénél (axondombnál) összegződhet (térbeli szummáció).
- Ha a helyi potenciálváltozások eredője eléri az ingerküszöb értékét, az axoneredésen ún. feszültség függő Na+-csatornák tömegesen nyílnak meg és az axonon (tovaterjedő) akciós potenciálhullám (sorozat) alakul ki (a folyamat pozitív visszacsatolás).
- A membrán depolarizálódik, a passzív transzporttal beömlő + töltésű Na+-ok átmenetileg + töltésűvé változtatják a membrán belső felszínét (+30 mV értékű csúcspotenciál).
- Ezt követően az axonon feszültségfüggő K+-csatornák nyílnak meg, K+-ionok tömegesen ömlenek diffúzióval az ECF felé, így a membrán repolarizálódik (negatív visszacsatolás).
- Mivel nagy mennyiségű + ion (K+) hagyja el a sejtet, ill. kis mennyiségű Cl- is beáramlik a sejtbe, a membrán hiperpolarizálódik.
- A nyugalmi potenciálnak megfelelő ioneloszlást és az ennek megfelelő potenciálértéket a Na+-K+ pumpa állítja helyre.
- A folyamat rendkívül gyors, kb. 3-4 ms múlva a nyugalmi potenciál az adott membrán szakaszon helyreáll.
- gyenge ingerek esetén olyan kevés átvivőanyag található meg a szipaptikus résben, hogy a posztszinapszisban a potenciálváltozás nem éri el az ingerküszöböt és csak lokális potenciál marad.
- Erősebb ingerek olyan nagy mennyiségű neurotranszmitter ürülését eredményezik, hogy a posztszinapszison rövid idő alatt sűrű akciós potenciálok jönnek létre.
- A digitális jelek meghatározott értékekből álló jellemzőkkel bírnak (0 vagy 1),
- az analóg jelek az időben folytonosan változó értéket vehetnek fel.
- A helyi potenciálok analóg,
- az akciós potenciálok digitális jelként értelmezhetők.
- A bejutó Na+-ok a még nyugalomban levő negatív töltésű helyek felé áramolnak a sejt belsejében,
- míg a sejt felszínén fordítva, a + ionok a depolarizált (negatívabb) helyek felé áramolnak.
- kis méretű a szinaptikus rés (2 nm),
- a csatornák nyithatók és zárhatók,
- az ingerületet többnyire ionok közvetítik,
- de átjuthatnak ATP, cAMP,
- a transzport – ingerületátadás - kétirányú,
- az ingerületvezetés gyorsabb.
- agy, retina,
- simaizomsejtek között (ábra),
- gerinctelenekben.
- Ingerületátvivő anyagok, neurotranszmitterek (ligand) közvetítik az ingerületet egyik sejtről a másikra.
- Pre- és posztszinaptikus membrán módosulatok alkotják.
- A kettő között szinaptikus rés (30-50 nm) található.
- Neurotranszmitterek szinaptikus hólyagokban tárolódnak a preszinapszisban, exocitózissal ürülnek.
- Az átvitel egyirányú, mivel a szinaptikus hólyagok csak a szinapszis előtti sejtvégződésben keletkeznek.
- Vannak serkentő és gátló szinapszisok, az átvivőanyag típusától és a receptorfehérje fajtájától függően.
- Pl. az acetilkolin a vázizmokon és szívizmon eltérő receptortípusokhoz kötődik,
- a vázizmokon Na+ csatornát nyit meg, tehát serkent,
- de a szívben K⁺ csatornát nyit meg, tehát gátol.
- Preszinapszis többnyire axonvég.
- Posztszinapszis többnyire sejttest, dendritek.
- A preszinaptikus idegsejt aktivitásakor – ingerületi állapotakor – a szinaptikus hólyagok tartalma Ca2+-beáramlás miatt exocitózissal a szinaptikus résbe ürül.
- Mennyiségük a preszinaptikus sejt akciós potenciáljának frekvenciájával arányos.
- A szinaptikus résben az ingerületátvivő anyag diffúzióval jut el a posztszinaptikus membránig, ott megkötődik specifikus fehérje receptorokon.
- A receptorok ioncsatornák, amelyek különféle ionok,
- a gátló szinapszisokban Cl- be-, ill. K+-kiáramlását,
- a serkentő szinapszisokban Na+ vagy/és Ca2+ -beáramlását teszik lehetővé a sejtbe.
- A beáramló ionok megváltoztatják a sejthártya töltésviszonyait (membránpotenciálját), egyúttal a sejt aktivitását.
- A serkentő szinapszisok csökkentik (-50 mV-ra),
- a gátló szinapszisok, melyek főleg az axoneredésénél – axondombon – vannak, növelik a membránpotenciált (-100 mV-ra).
- axonja elágazhat és több másik neuron aktivitását befolyásolhatja,
- ugyanakkor egy neuronhoz több különböző helyről érkezhet információ.
- acetilkolin: az ideg-izom szinapszisok átvivőanyaga, tanulásban, emlékezésben fontos,
- dopamin: mozgatópályák, jutalmazási – örömérzetet kiváltó - központok transzmittere,
- szerotonin: alvást, hangulatot, fájdalomérzetet szabályozó központokban termelődik,
- endorfinok: csökkentik a fájdalomérzetet, boldogságérzetet okoznak, fokozzák a dopamin felszabadulását,
- adrenalin, noradrenalin: pl. szimpatikus aktiváló rendszerek átvivőanyaga,
- gamma-amino-vajsav (GABA), gátló átvivőanyag (Cl--csatornákat nyit).
- részben enzimek bontják le őket (c),
- részben kimosódnak (b), ill.
- visszakerülnek aktív transzporttal, ill. endocitózissal a preszinapszisba (a).
- felszabadulásának fokozása (serkentők, pl. amfetamin), ill. gátlása (lassító drogok, pl. opiátok),
- visszavételének gátlása (serkentők, pl. kokain, amfetamin),
- lebontását végző enzimek hatásának módosítása útján.
- gátolják az átvivőanyagok visszavételét (kokain a dopaminét),
- lebomlását (pl. enzimgátlók, dopamin, noradrenalin, szerotonin),
- serkentik az ürülésüket (amfetamin). Az extasy pl. növeli a szerotonin, dopamin és a noradrenalin felszabadulását.
- gátolják a transzmitterek ürülését (opiátok: morfin és társai),
- vagy GABA-receptorokat serkentenek, melyek Cl--ioncsatornák, hiperpolarizációt okoznak (nyugtatók pl. benzodiazepinek (pl. xanax), a barbiturátok, ill. az alkohol).
- természetes hormonok, neurotranszmitterek,
- de lehetnek kívülről a szervezetbe juttatott vegyületek, mérgek, drogok.
- A gátló morfin K+-csatornák nyitása révén hiperpolarizációt eredményez.
- A THC (tetrahydrocannabinol) a kannabinoid-receptorokat aktiválja (CB1-receptorok), amelyek szerepet játszanak a hangulat, a fájdalomérzet, az étvágy és más funkciók szabályozásában.
- A nikotin serkentő hatását úgy éri el, hogy a központi idegrendszerben az idegsejtek acetilkolin receptoraihoz kötődik, aktiválva a működésüket (hatására a vérnyomás, szívfrekvencia nő, az idegsejtek fokozzák dopamin leadásukat, ami örömérzet és így a függőség kialakulásában játszik szerepet).
- szomatikus idegrendszerre, amely többnyire akaratlagos és vázizmokkal összefüggő, ill.
- vegetatív (autonóm) idegrendszerre, amely többnyire akaratunktól független, simaizmokkal, mirigyekkel, szívizommal kapcsolatos.
- Vérkeringési rendszer: szív, erek, vérnyomás szabályozása.
- Légzőműködések irányítása.
- Táplálkozással, emésztéssel kapcsolatos működések összehangolása.
- Kiválasztás, húgyhólyag működésének irányítása.
- Só- és vízháztartás szabályozása (ADH).
- Hőszabályozás lebonyolítása.
- Szexuális reflexek működtetése.
- Szem belső izmainak mozgatása (pupillareflex, akkomodáció).
- Hormonális rendszer egy része – mellékvesevelő, közvetlenül agyalapi mirigy és azon keresztül egyéb mirigyek, - működésének szabályozása.
- székletürítést, vizeletürítést,
- a bélmozgásokat,
- az erekció, ejakuláció reflexek működését,
- lokális hideghatásra a bőr ereinek reflexes összehúzódását.
- a keringés, légzés (vitális központok a nyúltvelőben),
- köhögés, tüsszentés, nyelés, hányás, nyálelválasztás (nyálkahártyareflexek központjai a nyúltvelőben),
- a szem alkalmazkodásának (pupillareflex, akkomodáció) reflexközpontjai találhatók a középagyban.
- Táplálkozás, folyadékfelvétel (ozmoreguláció), hőszabályozás, hormonális rendszer irányító központjai helyezkednek el.
- Vegetatív rostot is tartalmazó gerincvelői idegek, a nyaki szakasz kivételével.
- Vegetatív rostot is tartalmazó agyidegek.
- Lehetnek közvetlenül a gerinc mellett, mint pl. a szimpatikus határlánc dúcai,
- vagy a szervek előtt, azok falában, mint pl. a paraszimpatikus rendszer dúcai.
- Belső szervek (belek, erek, tüdő) falában,
- vagy maga a központ a receptor
- az agytörzsben a CO2-ra érzékeny légzőközpont, vagy a presszor központ,
- a hipotalamuszban a vér ozmotikus koncentrációjára érzékeny szomjúságközpont, ill. a vércukorszintre érzékeny éhség- és jóllakottsági központ.
- nyomásreceptorok: szív körüli nagyerek falában,
- feszítési receptorok: tüdő, bélfal, húgyhólyag falában,
- kemoreceptorok: szív körüli nagyerek falában, központi idegrendszerben,
- termoreceptorok: bőrben, májban, gyomorban.
- a hólyag-, a végbélürítő reflex,
- az erekció, ejakuláció szabályozása,
- bélmozgások,
- bőr ereinek reflexes működtetése.
- A mellsőgyökéren keresztül hagyják el a gerincvelőt, majd
- gerincvelőidegekben futnak a szervekhez.
- A mozgatóneuronok nem a központi idegrendszerben, hanem a testben található vegetatív dúcokban vannak.
- általános aktivitását fokozza, az erőtartalékait mozgósítja, testünket készenléti állapotba hozza.
- Felkészíti a szervezetet a nagyobb terhelések elviselésére.
- a mellékvesevelőből az adrenalin, a noradrenalin felszabadulás fokozódik,
- a légzés gyorsul, a légutak tágulnak,
- a vázizmok erei tágulnak,
- a vérnyomás, szívfrekvencia nő.
- a vércukorszint emelkedik (máj glikogén lebontása nő),
- a lebontó folyamatok túlsúlyba kerülnek,
- a fűtőközpont aktivitása nő, bőr erei szűkülnek,
- a pupilla tágul.
- A bélműködés – elválasztás, perisztaltika - csökken, a bélcső erei szűkülnek.
- részben agyidegek, pl. X. bolygóideg,
- részben keresztcsonti gerincvelői idegek útján lépnek ki.
- ürítés: hányás, székelés, vizelés,
- bélműködés serkentése: emésztőnedvek elválasztásának serkentése, bélmozgások fokozása,
- felépítő folyamatok túlsúlyának elősegítése,
- közelnézéshez akkomodáció,
- hűtőközpont működésének fokozódása.
- Erekció.
- idegi – szimpatikus idegrendszer, lásd 46. tétel -,
- részben hormonális mechanizmusok – adrenalin, kortizol - felelősek.
- A felgyorsult életvitellel kapcsolatos mindennapi események (például tömegközlekedés, munkavégzés stb.).
- Az egyén életének nagyobb fordulatai: vizsgák, költözés, állásváltoztatás, elbocsátás, betegség, egy személy halála stb.
- Belső konfliktusok, pl. párkapcsolati problémák, elfojtott érzelmek, gondolatok, vágyak okozta belső feszültség stb.
- Háborúk, katasztrófák.
- Akut: hirtelen előállt stresszhelyzetben. Az enyhébb akut stressznek lehetnek akár pozitív hatásai is, mivel segítheti a nehézségekkel való megbirkózás képességének, a megküzdési technikák fejlődését.
- Krónikus stressz: tartós, hosszabb ideig fennálló megterhelés hatására alakulhat ki.
- Magas stresszhormon szint: adrenalin, kortizol.
- Stresszhelyzetben fokozódik a hipotalamusz (CRF) faktorának elválasztása,
- ami az agyalapi mirigy mellékvesekéreg-serkentő hormonjának (ACTH) elválasztását fokozza,
- aminek hatására a mellékvesekéregből kortizol,
- továbbá idegi hatásokra mellékvesevelőből adrenalin szabadul fel.
- Magas vércukorszint.
- Emelkedett vérnyomás, magas koleszterinszint, fokozódik az érelmeszesedés.
- Immunrendszer működése gyengül, gyakoriak a betegségek, daganatos betegségek kockázata nő.
- Izomfeszülés (nyaki, háti, mellkasi izmokban).
- Hányás, hasmenés.
- Fokozott verejtékezés, hideg kéz-láb, remegés, rángatózás, dadogás.
- Menstruációs zavarok, szexuális vágy megszűnése.
- Szorongás.
- Pánikrohamok.
- Depresszió, ingerlékenység, alvászavar.
- Soványság, vagy éppen elhízás elhízás.
- Tanulási, koncentrálási és térbeli tájékozódási zavarok.
- adrenalint,
- noradrenalint, amelyek az idegrendszerben is előforduló ingerületátvivő anyagok.
- emeli a vércukorszintet, csökkenti a máj glükóz raktározását, serkenti a glikogén bontását (cAMP) fokozza a keletkezett glükóz leadását,
- a sejtekben – főleg a vázizomzatban - serkenti a glükóz oxidációját,
- serkenti a zsírok lebontását, emeli a vér zsírsavszintjét,
- emeli a vérnyomást,
- növeli a szívfrekvenciát, az összehúzódások erejét,
- tágítja a hörgőket,
- tágítja a vázizmok ereit, továbbá a koszorúereket,
- a bélcső, bőr ereit ugyanakkor szűkíti.
- A vázizmokban futó erek simaizomsejtjeinek felszínén található ún. β2 receptorok ingerlésével az erek simaizmai ellazulnak, aminek hatására az erek átmérője megnő.
- Ugyanakkor a bélfalban elhelyezkedő simaizomsejtek felületükön α1 típusú receptorokat hordoznak, melyek ingerlése a simaizomsejtek összehúzódását eredményezi, aminek következtében az erek szűkülnek.
- emeli a vércukorszintet,
- serkenti a zsírok lebomlását,
- serkenti az izomfehérjék bontását, a májba jutott aminosavak cukrokká alakítását (glükoneogenezis),
- a májba jutott tejsavból elősegíti a glükóz képződését (glükoneogenezis, Cori-kör).
- Összességében az inzulin antagonistája.
- Stresszhormon, a stresszhelyzetekben döntő szerepet játszik az anyagcsere szabályozásában, pl. a zsíroxidáció fokozásával energiát biztosít a szervezet számára,
- ugyanakkor gátolja az immunrendszer működését, ezért gyulladáscsökkentő, az allergiás reakciókat mérsékeli, antihisztamin hatású.
- Granulociták
- Limfociták
- Monociták
- B limfociták, melyek a vörös csontvelőben termelődnek – érnek be - és az ún. szerzett (adaptív, specifikus) humorális immunválasz kialakításában vesznek részt.
- T limfociták, amelyek előalakjai szintén a vörös csontvelőben jönnek létre, azonban a sejtek kivándorolnak a tímuszba (csecsemőmirigy) és fejlődésüket – érésüket - ott fejezik be. A T-sejtek az ún. szerzett (adaptív, specifikus) sejtes, azaz celluláris immunválaszért felelősek.
- A veleszületett – természetes - immunrendszer elemeként legfontosabb feladatuk a kórokozók és apoptotikus sejtek azonnali felismerése, bekebelezése és feldolgozása.
- A gyulladás kialakulásában is közreműködnek.
- Antigénbemutató sejtként képesek a T- és B-sejteket aktiválni.
- a testbe bekerült idegen anyagokat, parazitákat, ill.
- a potenciális veszélyforrást jelentő megváltozott saját sejteket - vírussal fertőzött sejteket és a képződött tumorsejteket - felismerje és eltávolítsa, megsemmisítse.
- Szervezetünk elsődleges fizikai védelmi vonalát a bőr és a nyálkahártya szövetei képezik. Ezek a jól záródó határrétegek megakadályozzák a káros anyagok, a baktériumok és a vírusok bejutását a testbe.
- Továbbá a légutak nyálkahártyáinak felületén található csillók folyamatosan eltávolítják a nyákba beletapadt kórokozókat, ill. idegen anyagokat pl. port, pollent.
- A gyomor erősen savas kémhatása fizikai-kémiai védelmet nyújt a táplálékkal bekerült kórokozókkal szemben. A savas kémhatás többnyire megakadályozza ezeknek a szervezeteknek a túlélését és a szaporodását.
- A nők hüvelyében szimbiózisban élő tejsavat termelő baktériumok savas kémhatás kialakításával védik a női ivarutakat, elsősorban a különféle gombás fertőzésektől.
- leginkább az immunrendszer sejtjei által termelt különféle vegyületek – antitestek -
- a máj által termelt komplementrendszer tagjai,
- a légző- és emésztőrendszeri váladékokban megtalálható antibakteriális anyagok (pl. lizozim, mely a baktériumok sejtfalát bontja) biztosítják.
- egyrészt az emberi szervezettel együtt élő ún. mikrobiom,
- másfelől a saját immunrendszerük teszi lehetővé.
- segítenek az emésztésben, az emészthetetlen növényi rostokat a vastagbélben erjedéssel bontják és rövid láncú zsírsavakat termelnek, amelyek segítik a bélhámsejtek egészségének fenntartását, csökkentik a vastagbélrák kialakulásának a kockázatát,
- szintén a vastagbélben B- és K-vitaminokat termelnek,
- antibiotikumok termelésével, tápanyagokért történő versengéssel, ill. helyfoglalással távol tartják a káros mikroorganizmusokat,
- erősítik az immunrendszerünket,
- befolyásolják az anyagcserét, részt vesznek a testtömeg-szabályozásban.
- A bőr faggyúrétegében szimbióta baktériumok tapadnak meg, savas váladékot (pH 5,5) termelve védik a bőrt a gombás fertőzésektől. Ebből is látszik, hogy sok esetben a fizika-kémiai védelem nem különíthető el élesen a biológiai védekezéstől.
- Jelentős szerepet játszanak az idegrendszer működésében is, fontos szabályozóanyagokat termelnek szerotonint, melatonint, adrenalint.
- Az immunogén olyan antigén, mellyel szemben immunválasz indul meg.
- A tolerogének olyan antigének, mely természetes módon saját sejtjeink felületén találhatók meg, ezeket az immunrendszer sajátként ismeri fel, normálisan nem indul immunválasz ellenük, pl. ilyenek az MHC-molekulák, melyek a sejtek felszínén helyezkednek el.
- a szervezeten kívülről származik heteroantigénről,
- ha a saját szervezetből származik autoantigénről beszélünk.
- Vírusok burokfehérjéi.
- Baktériumok, egysejtű kórokozók sejtfelszíni molekulái.
- Baktériumok, gombák citoplazmatikus toxinjai, melyek kikerülve a sejtekből válnak antigénné.
- Szervezetidegen makromolekulák, elsősorban fehérjék, szénhidrátok.
- Szervátültetésnél a szervezetbe került idegen sejtek felszíni molekulái. Az ún. transzplantációs antigének miatt a beültetett szerv általában kilökődik, amit úgy próbálnak lassítani vagy megakadályozni, hogy a befogadó szervezet immunrendszerének működését gyógyszeresen korlátozzák.
- Vérátömlesztésnél a vörösvértestek felületén található ún. vércsoportantigének, melyek a különféle vércsoportok kialakításáért felelősek.
- nem specifikus (veleszületett vagy természetes), amely az antigén fajtájától függetlenül hasonlóan játszódik le.
- specifikus (adaptív vagy szerzett).
- különböző vegyületekhez kötött humorális (humor = folyadék)
- és az igen sokféle immunsejttel kapcsolatos sejtes (celluláris) immunválasz.
- Az aktiválódási lánc utolsó elemei fehérjékből csatornaszerű pórust alakítanak ki a megtámadott sejt sejthártyájában, az ozmotikus egyensúly megbomlik, a sejt elpusztul.
- Fontos szerepet töltenek be a gyulladás mechanizmusában is.
- Hozzákötődve antigénekhez megjelölhetik azokat a fagociták számára.
- fagocitái a mikrofágok és a makrofágok
- és a természetes ölősejtek (NK) vesznek részt.
- ún. perforinok segítségével a sejthártyán lyukakat „fúrnak”, ami ozmotikus sejthalált eredményez,
- a patogének számára egyéb mérgező vegyületeket termelnek és ürítenek,
- vagy a sejtekben programozott sejthalál (apoptózis) beindításával fejtik ki védő hatásukat.
- vagy olyan molekuláris mintázatokat – baktériumok felületén előforduló lipopoliszacharidokat - ismernek fel receptoraik segítségével, amelyek tipikusan patogének felszínén jelennek meg, de a saját sejtekre nem jellemzőek,
- vagy az antigénekhez kapcsolódott antitestek bizonyos jellemző részeit azonosítják, ami beindítja a fagocitózist.
- A B-limfociták a humorális,
- a T-limfociták a sejtes (celluláris) immunfolyamatokat bonyolítják le.
- plazmasejtekké alakulnak, melyek nagy mennyiségben állítanak elő ellenanyagot, amit a kibocsájtanak a sejten kívüli térbe, ill.
- hosszú életű memóriasejtté differenciálódnak, melyek egy későbbi fertőzés során azonnali másodlagos immunválaszt tesznek lehetővé. Az immunmemória alapját képezik.
- A segítő (helper) sejtek (Th) szabályozóanyagokat ún. citokineket termelnek, amelyek osztódásra késztetik a már aktiválódott B-limfocitákat, makrofágokat, és egyéb T-sejteket. A citokinek felelősek többek között a gyulladás és a láz kialakulásáért.
- A memória T-sejtek hosszú ideig élnek és tartós védettséget biztosítanak.
- Az elnyomó, szupresszor T-sejtek (Treg) visszaállítják a fertőzés előtti állapotot úgy, hogy gátolják az immunsejtek osztódását.
- A testnedvekben – pl. vérben, nyirokban - szabadon keringve hozzákötődhet a neki megfelelő antigénhez kicsapva, ill. megjelölve azt.
- Az antigénhez kötődve fagocitózisra készteti a falósejteket, pl. a makrofágokat.
- Az antigénhez kötődve mozgósítja a komplementrendszert.
- A B-sejtek a maguk által termelt sejtfelszínre kihelyezett antitestekkel.
- A T-limfociták a makrofágok által feldarabolt és "bemutatott" peptideket ismerik fel, méghozzá a bemutató sejt MHC-molekuláinak kíséretében. Más szóval, a T-sejtek egy módosított ("saját nyelvükre lefordított") formában képesek csak az antigén felismerésére.
- a petefészkek,
- a petevezetők,
- a méh, és
- a hüvely.
- egyrészt ivarsejteket hoznak létre, ill. érlelnek,
- másrészt szteránvázas nemi hormonokat termelnek.
- közepén egy diploid, éretlen „petesejt” helyezkedik el,
- amelyet egyetlen sejtréteg, a tüszőhámsejtek rétege vesz körül.
- az érett petesejtet (a másodrendű oocytát), amely már haploid
- és a sarki sejtet, a polocitát.
- A „petesejtek” (I. rendű oocyták) már a magzati élet során létrejönnek, számuk már születéskor adott. Érési osztódások a pubertástól csak a menopauzáig vannak.
- Ezzel szemben gyakorlatilag a spermatogenezis összes szakasza – a spermatogóniumok keletkezésének kivételével, mely az embrionális korban zajlik - a pubertáskor kezdődik és az élet végéig tart.
- Az oogenezis eredménye 1 teljes értékű petesejt és 3 sarkisejt.
- A spermatogenezis során 4 teljes értékű spermium jön létre.
- A petesejtek mindig csak X ivari kromoszómát tartalmaznak,
- a spermiumok fele X, másik fele Y ivari kromoszómát.
- megtermékenyülés esetén a terhesség kb. 3. hónapjáig fennmarad, folyamatosan ösztrogént termel, ill. a főleg az embrió beágyazódását és fejlődését segítő progeszteront.
- Azonban, ha a megtermékenyítés elmarad, csak kb. 10 napig működik, utána elsorvad, ezért a progeszteronszint a vérben újra lecsökken.
- közvetlenül az agyalapi mirigy szabályozza az FSH és az LH elválasztásán keresztül,
- közvetve pedig a hipotalamusz a kissejtes magvakban keletkező GnRH révén.
- elsősorban a fejlődő tüszőben jön létre, kisebb mennyiségben a sárgatestben és a zsírszövetekben is keletkezik,
- elválasztása a pubertáskorban kezdődik (az ösztradiol kis mennyiségben a férfiak zsírszövetében is keletkezik), a menopauza után megszűnik.
- Elősegíti az elsődleges és a másodlagos nemi jellegek kialakulását, fejlődését,
- hatására érik el végleges alakjukat és méretüket a külső és a belső nemi szervek,
- kiváltja az emlő fejlődését, a méhfal erőteljes vastagodását,
- jellegzetes női zsírfelhalmozódást eredményez,
- kialakítja a nőies pszichét.
- A ciklus alatt biztosítja a méhnyálkahártya regenerálódását, vastagodását.
- Termelődésének maximumát az ovuláció előtt éri el a 12.-13. napon, elválasztását
- közvetve a hipotalamusz GnRH-ja,
- közvetlenül pedig a tüszőserkentő hormon növeli (mivel serkenti a tüszőérést).
- Az FSH-, LH-elválasztást
- nagy koncentrációban a ciklus közepén serkenti (pozitív visszacsatolással),
- a ciklus második felében alacsonyabb koncentrációban a progeszteron jelenlétében pedig már gátolja (negatív visszacsatolás).
- a ciklus második felében a sárgatestben keletkezik.
- Megtermékenyítés esetén a sárgatest fennmarad és kb. a terhesség 3. hónapjáig termeli a progeszteront (később a méhlepény választja el a terhesség végéig),
- ellenkező esetben a 24. naptól kezdve szintje néhány napon belül minimumra csökken.
- Előkészíti a méh falát az embrió befogadására, hatására a nyálkahártyája eléri maximális vastagságát,
- bezárja a méhszájat,
- akadályozza a méhösszehúzódásokat,
- 0,5 fokkal emeli a testhőmérsékletet.
- Hatására fejlődésnek indul az emlő mirigyes állománya, előkészíti a tejelválasztást, de gátolja a tejtermelést.
- Gátolja az FSH és az LH termelődését, ezért megakadályozza terhesség esetén az újabb tüszőérést. Ezen a hatáson alapszik a hormonális fogamzásgátlás, melynek során az ösztrogén- és progeszterontartalmú tablettákkal megakadályozzák a tüszőérést és a tüszőrepedést.
- kívülről vékony hashártya borítja,
- középen vastag simaizomréteg (myometrium) helyezkedik el,
- az üreg felőli részét pedig nyálkahártya (endometrium) béleli.
- A ciklus 1. napjától a kb. 4-5. napig tartó szakasz a menstruációs fázis. A menstruáció során az előzőleg megvastagodott, vérbő nyálkahártya 2/3-a vérzés kíséretében lelökődik.
- A menses után, ösztrogén hatására a helyenként megmaradt nyálkahártyafoltokból megindul a regeneráció.
- Ezt követően a burjánzási fázisban a ciklus közepéig a nyálkahártya egyre magasabb lesz, erekben, mirigyekben gazdagodik. A helyreállítási és a burjánzási fázisok kialakulásáért a petefészek ösztrogén hormonja felelős, ezért ezt a két szakaszt együtt az ösztrogénhatás fázisának is nevezzük.
- Az ovulációt követően az elválasztási fázisban a nyálkahártya eléri a maximális vastagságát (4-5 mm), abszolút vérbő, mirigyei váladékot termelnek, mely az odaérkező embrió beágyazódását biztosítja. A nyálkahártya maximális vastagságának a kialakításáért a sárgatest progeszteron hormonja felelős (progeszteronhatás fázisa).
- a petesejt megtermékenyül, a fejlődő embrió egy hCG nevű hormont termel, aminek a hatása, hogy fenntartja a sárgatestet kb. 3 hónapig, miáltal biztosítja a folyamatos progeszterontermelést, ami lehetővé teszi a vastag nyálkahártya fennmaradását, az embrió befogadását, táplálását.
- Azonban, ha nem történik megtermékenyítés, a sárgatest sorvadásnak indul, a progeszteron koncentrációja a vérben erősen csökken, aminek következtében a megduzzadt nyálkahártya vérzés kíséretében (menstruáció) lelökődik.
- Az egyedfejlődés a megtermékenyítéssel veszi kezdetét.
- Ezt követően megindul a barázdálódás, mialatt gyors, egymást követő osztódásokkal kialakul az ún. szedercsíra.
- Létrejönnek a csíralemezek, majd
- a szervtelepekből kialakulnak az embrió szervkezdeményei.
- Az embrionális szervkezdemények később funkcionáló szervekké alakulnak.
- egy haploid petesejt (másodrendű oocyta) és
- egy sarki sejt, tüszőhámsejtekkel (corona radiata) körülvéve.
- A hímivarsejtek akroszómális enzimek segítségével a petesejtet körülvevő sejtek közötti kapcsolatokat fellazítják, s így hozzáférhetnek a petesejtburokhoz (zona pellucida).
- A fellazított fénylő rétegen (zona pellucidán) a hímivarsejt keresztülfúrja magát, és a külső membránján található kötőfehérjék a petesejt ZP3 fehérje receptoraihoz kapcsolódnak.
- A spermiumok hiperaktív mozgásúak lesznek, az ostor erőteljes csapásokkal mozgatja előre a sejteket.
- Az első hímivarsejt feji részének bejutását követően a petesejt sejthártyája alatt a citpolazmában található hólyagokból többféle, nagy mennyiségű bontó enzim ürül exocitózissal a petesejt felszínére, melyek felszámolják a ZP3 receptorokat és megkeményítik a petesejt burkát (kortikális reakció).
- A petesejt és spermiummembrán egyesülését követően befejeződik a II. meiotikus osztódás.
- A zigóta 24 óra múlva mitózisokkal osztódni kezd (barázdálódás), majd 3-4 nappal a megtermékenyítés után 16 sejtes ún. szedercsíra (morula) állapotba kerül.
- A szedercsírának van egy ún. belső és egy külső sejttömege.
- A szedercsíra belső sejtjeinek egy része apoptózissal elhal, a megmaradó sejtek pedig egy ponton gyűlnek össze, létrehozva az embriócsomót.
- A külső nagyobb sejtek később a külső magzatburkot hozzák létre. Ezt a belül üreges képződményt hólyagcsírának(blastula) nevezzük.
- A hólyagcsíra megtapad a méh nyálkahártyáján, elkezd bontóenzimeket termelni és besüllyed a méhnyálkahártya kötőszöveti állományába. Ez a folyamat a beágyazódás, az implantáció.
- az embrió rögzítése és táplálása,
- részt vesz a méhlepény kialakításában,
- hCG hormont (human Chorion Gonadotrop hormon) választ el, amely a 10. naptól fenntartja a sárgatestet (így az tovább termeli a progeszteront).
- véd a külső mechanikai hatásoktól,
- a kiszáradástól és
- a hőmérséklet-változásoktól,
- a magzat szabad, szinte lebegő mozgását biztosítja.
- a herék,
- a mellékherék,
- az ondóvezetékek,
- az ondóhólyagok,
- a prosztata, és
- a Cowper-féle mirigyek.
- egyrészt hímivarsejteket termelnek,
- másrészt nemi hormonokat – tesztoszteron – választanak el, ezért belső elválasztású mirigyeknek is tekinthetjük őket.
- az aktív hímivarsejteket,
- az ondóhólyag fruktóz- és prosztaglandintartalmú termékeit,
- a prosztata lúgos kémhatású, mennyiségét tekintve legjelentősebb folyadékát.
- a csatornácskák közötti kötőszövetben elhelyezkedő Leydig-féle köztes sejtek tesztoszteront (ösztrogént),
- a Sertoli-féle sejtek inhibint termelnek.
- egyrészt serkenti a Sertoli-féle sejtek működését, a spermiumok képződését,
- másrészt felelős magzati korban az elsődleges, serdülőkorban a férfiak másodlagos nemi jellegeinek kialakításáért,
- a zsírszövetekben ösztrogénné alakulhat.
- a vállöv és a mellkas erőteljes, a medence szűk és magas,
- kifejezett arc és testszőrzet,
- a gégefő erősebb, a hang mélyebb,
- a psziché férfias.
- A tüszőserkentő hormon a Sertoli-sejtekre hat, serkenti a spermiumok képződését,
- a sárgatestserkentő hormon a Leydig-sejtek működését, s ezáltal a tesztoszteron termelődését fokozza.
- A Sertoli-sejtek hormonhatású inhibinje a tüszőserkentő hormon,
- a Leydig-sejtek tesztoszteronja a sárgatestserkentő hormon termelődését gátolja.
- azonos fajhoz tartozó egyedek,
- amelyek adott helyen és időben együtt élnek,
- egymás között szaporodnak, a faj folytonosságát fenntartó tényleges szaporodási közösséget alkotnak (térben és időben is elkülönülnek a faj más populációitól).
- születések és a halálozások számától, azaz a szaporodási rátától,
- a ki- és a bevándorlások számától,
- az egyedek koreloszlásától,
- az ivararánytól.
- a környezet eltartóképességétől, a populáció sűrűségétől,
- környezeti tényezők optimális értékeitől való eltérés mértékétől,
- más populációktól (ragadozók, élősködők, versenytársak számától).
- emberi populációk esetén az 1000 lakosra jutó születések száma.
- Állatoknál a populáció női ivarú egyedeinek egységnyi idő alatt létrehozott utódainak a száma.
- Értéke fajonként eltérő.
- a koreloszlástól (a populáció szaporodásra képes egyedeinek számától),
- az ivararánytól,
- szaporodásbiológiai sajátosságoktól (magok, termés mennyisége, peteszám, generációk száma évente stb.).
- A potenciális (korlátlan) szaporodóképesség a lehető legmagasabb elérhető utódszám. Ideális feltételek között a populációk korlátlanul növekedhetnének. Az egyedszámváltozást az idő függvényében ábrázolva egy exponenciális görbét
- A reális (korlátozott) szaporodóképesség a ténylegesen megvalósuló szaporodás mértéke. A potenciális szaporodóképességet korlátozza a környezet eltartóképessége (Kapacitás).
- megmutatja, hogy időegység alatt mekkora a populáció létszámának növekedése,
- általában új élőhelyek benépesítésekor a legmagasabb a táplálékbőség miatt,
- kis egyedszám (sűrűség) esetén a legnagyobb (exponenciális). Ez persze felveti a beltenyészet veszélyét, ami viszont az egyedek genetikai leromlásához vezet. Ezért populációknak van egy kritikus egyedszáma – ún. kihalási küszöb -, amely alatt már a szaporodóképesség rohamosan csökken (genetikai okok, lásd 6.3.1. fejezet).
- születési rátája b) csökken,
- a halálozási ráta d) pedig emelkedik.
- A két ráta metszéspontjában az állomány egyensúlyba kerül (K, egyensúlyi állományméret vagy eltartóképesség).
- E feletti egyedsűrűség – túlnépesedés - esetén a populáció mérete csökken. Okok:
- a táplálék megfogyatkozása,
- az élősködők elszaporodása, járványok,
- a ragadozók nagyobb számban jelennek meg,
- tömegstressz következményei,
- élőhely-beszűkülés.
- függőleges tengelyen az egyes korcsoportokat,
- a vízszintes tengelyen az y tengelytől jobbra a nők, balra a férfiak arányát tüntetjük fel.
- az egészen fiatal egyedek száma meghaladja a középkorú és az idős egyedek számát. A természetben az állatokra,
- ill. a harmadik világ elmaradott népességeire jellemző.
- a különböző korosztályokhoz tartozó egyedek aránya nemzedékről nemzedékre változatlan marad.
- Fejlett ipari társadalmakra jellemző.
- Élőhelyeik környezeti feltételei állandóak,
- szaporodási rátájukat az egyedsűrűség, az egyedsűrűségüket a környezet eltartóképessége (K) határozza meg,
- egyedei hosszú életűek, általában nagy testűek,
- hosszú idő után válnak ivaréretté,
- utódaik száma csekély,
- fejlett az ivadékgondozás, az utódok felnevelésébe sok energiát fektetnek,
- a szaporodásuk során magas az utódok túlélési rátája,
- ilyenek a nagy testű gerincesek és a fák.
- Élőhelyeik környezeti feltételei gyorsan változhatnak,
- többnyire időszakosan megjelenő élőhelyeken (pl. kidőlt fa helyén, pocsolyák, bolygatott élőhelyek, árterek) fordulnak elő,
- elsőként jelennek meg egy társulás kialakulásánál (pionírtársulás-alkotók),
- utódaik száma jelentős, szaporodási rátájuk magas (exponenciális), K-értékük gyakorlatilag nincs, szaporodási rátájuk független az egyedszámtól,
- egyedei rövid életűek, kis termetűek,
- rövid idő után válnak ivaréretté,
- nagyszámú utódjuk nevelésére kevés energiát fektetnek, az ivadékgondozás sokszor nem jellemző,
- a szaporodásuk során az utódok túlélési rátája alacsony,
- nem versenyképesek.
- Ilyenek lehetnek pl. a rovarok, gyomnövények, többnyire a rágcsálók.
- abiotikusak (élettelen):
- talaj,
- levegő,
- fény,
- hőmérséklet,
- víz,
- biotikusak, amelyek az élőhelyen együtt élő populációk egymásra hatásából következnek.
- Megmutatja, hogy egy életközösség – pl. populáció - hogyan reagál a környezeti tényezők változásaira.
- A környezeti tényezők azon tartománya, amelyen belül a populáció előfordulhat,
- tehát a tűrőképesség meghatározza a populáció elterjedését, előfordulását.
- Az evolúció során kialakult, a populáció egyedeinek öröklődő alkalmazkodóképessége az adott környezeti tényező változásaihoz.
- x tengelyen a környezeti tényező értékeit,
- az y tengelyen a szaporodóképességet, egyedszámot stb. tüntetjük fel.
- Az éppen elviselhető alsó, ill. felső érték minimum és a maximum.
- Az optimum a legkedvezőbb, a pesszimum még elviselhető értéktartomány.
- A grafikon megmutatja, hogy a populáció milyen mértékben alkalmazkodott az adott környezeti tényező értékeihez és annak változásaihoz.
- Az élettani tűrőképesség a többi fajjal (populációival) való kölcsönhatás nélkül,
- az ökológiai tűrőképesség e kölcsönhatások figyelembevételével mutatja meg a faj életlehetőségeit.
- Növények esetében a fényért, vízért, tápanyagokért, élőhelyért stb.
- Állatok esetében a táplálékért, a vadászterületért, búvóhelyért stb.
- abiotikus (hőmérséklet, páratartalom stb.) környezeti tűrőképessége,
- az elfogyasztott táplálék típusa,
- ragadozói,
- viselkedési szokásai,
- stb.
- legkisebb mennyiségben van jelen, vagy
- amelyik mennyisége leginkább eltér az optimálistól.
- Jelentősebb változás,
- egyes populációi egy ideig feltűnő módon, mások pedig rejtetten vannak jelen.
- Például a lomberdők tavaszi, nyári, őszi és téli változásai, aszpektusai.
- Lombfakadás előtt nyílnak a fényigényes, többnyire gyöktörzses vagy hagymás évelő növények, termést és magot érlelnek, majd föld feletti részeik elszáradnak (a kora tavaszi ún. geofitonok, az év nagy részét a talajban töltik hagymák, gyöktörzsek, gumók formájában, ilyen pl. az odvas keltike).
- majd a lombosodást követően az árnyéktűrők jelennek meg.
- Állatok egy része az egyes aszpektusokban nincs jelen, pl. vándormadarak, rovarok egy része.
- az alföldön tatárjuharos-lösztölgyes (már csak foltokban (mezőgazdaság)),
- cseres-kocsánytalan tölgyesek (cseres-tölgyesek) 250 - 450 m,
- gyertyános-tölgyesek 450 - 600 m,
- bükkösök 600 m-től,
- elegyes fenyves erdők (az ország legnyugatibb részein, csak kis területen).
- Középhegységeinkben és dombvidékeinken tszf. 250-450 méteres magasságban jellemző. A leggyakoribb erdőtípus hazánkban.
- Laza a lombkorona, amely a fényt jól átengedi.
- Jellemző a csertölgy és a kocsánytalan tölgy (barkócafa (szórtan), virágos kőris szárazabb helyeken).
- A fejlett cserjeszint és fejlett gyepszint fény- és melegkedvelő fajokban gazdag.
- Cserjeszintben egybibés galagonya, kökény, fagyal, vörösgyűrű som fordul elő,
- a gyepszintben egyvirágú gyöngyperje, tavaszi lednek, nyáron baracklevelű harangvirág, pillangósvirágúak, vitéz bükköny található.
- Középhegységeinkben 450-600 méter magasságában fordulnak elő.
- Kétszintű lombkorona alakul ki, ahol:
- a felső szintet a kocsánytalan vagy kocsányos tölgy,
- az alsó szintet gyertyán alkotja.
- A kevesebb fény miatt jelentéktelen a cserjeszint és a gyepszint.
- Kora tavaszi aszpektusa gazdag geofitonokban, amelyek tavasszal virágzanak, nyárra mire a lombkorona záródik magjaikat elszórják, visszahúzódnak földalatti raktározó szerveikbe (gumó, hagyma, gyöktörzs) és az év nagy részét föld alatt töltik.
- Fajai: odvas keltike, medvehagyma, bogláros szellőrózsa, salátaboglárka.
- Középhegységeink 600 méternél magasabban fekvő területein jellemzőek.
- Sűrű erdő, a lombkorona teljesen záródó, cserjeszint nincs, gyepszint csak kora tavasszal (geofitonok, hagymások, gumósok) fordul elő.
- Uralkodó fája a bükk.
- függőlegesen
- és vízszintesen.
- A legalsó szint, amely a termőtalaj felszíni rétegétől a legmélyebbre lehatoló gyökerekig tart,
- termelői kemotróf nitrifikáló baktériumok,
- fogyasztói a pl. vakond és ízeltlábúak,
- lebontói felaprózást végzők (talajlakó atkák, rovarok, gyűrűsférgek, ikerszelvényesek), továbbá gombák, baktériumok stb.
- Bomlásnak indult és a frissen lehullott levelek szintje.
- Termelő szervezetek nem jellemzők rá.
- Fogyasztói gerinctelenek (csigák és ízeltlábúak),
- lebontó szervezetek: ízeltlábúak, gombák és baktériumok.
- A társulás lágyszárú szintje.
- Néhol alig éri el a 20-30 cm-es magasságot, másutt akár két méteres is lehet.
- Fontos termelői szint.
- Fogyasztói főleg ízeltlábúak.
- A bokrok lombozatát jelenti.
- Gazdag az ízeltlábú faunája.
- A bokrok lombozatában sok madár fészkel (feketerigó, a tövisszúró gébics, ökörszem).
- A társulások legfelső szintje, a fák lombozatából áll.
- Több alszintre tagolódhat, pl. kettős lombkoronaszint jellemző a gyertyános tölgyesekre, melyekben az alsó szintet a gyertyán, a felső szintet a kocsánytalan tölgy lombozata jelenti.
- Madarak, egyes emlősök előfordulásának jellemző szintje.
- víznek és a tápanyagoknak a talajban való mennyisége és egyenlőtlen eloszlása, s az értük folyó versengés, ill.
- a populációk közötti kölcsönhatások – pl. kommenzalizmus - alakítják ki.
- Csupán látszólagos változás, amely az egyes élőlények – főleg állatok – napi aktivitásával függ össze.
- Vannak nappali és éjszakai állatok, egyes növények virágnyílása is napszakokhoz köthető.
- Jelentősebb változás,
- egyes populációi egy ideig feltűnő módon, mások pedig rejtetten vannak jelen.
- Például a lomberdők tavaszi, nyári, őszi és téli változásai, aszpektusai.
- Lombfakadás előtt nyílnak a fényigényes, többnyire gyöktörzses vagy hagymás évelő növények, termést és magot érlelnek, majd föld feletti részeik elszáradnak (a kora tavaszi ún. geofitonok, az év nagy részét a talajban töltik hagymák, gyöktörzsek, gumók formájában, ilyen pl. az odvas keltike),
- majd a lombosodást követően az árnyéktűrők jelennek meg.
- Állatok egy része az egyes aszpektusokban nincs jelen, pl. vándormadarak, rovarok egy része.
- természetes
- és az emberi hatásnak – zavarás, erdőégetés - köszönhető másodlagos formája.
- Földtörténeti léptékű.
- Több ezer évet átfogó szukcesszió.
- Egyirányú, nem ismétlődő folyamat, az éghajlat megváltozása következtében lejátszódó.
- Nagy területet érintő változások (pl. Kárpát-medence).
- Ilyen pl. az utolsó jégkorszakot követően Magyarországon a tundrai társulások átalakulása.
- Rövidebb (száz években számolható),
- azonos éghajlati körülmények között zajló folyamat.
- Első lépése egy növényzet nélküli, kopár terület benépesítése.
- Az első betelepedő élőlények alkotják az ún. nyitó- vagy pionír társulást.
- A pionír társulások
- fajban szegények, alkotó populációik pedig egyedszámban viszonylag gazdagok,
- r-strategisták, fajaik apró, könnyen terjedő magvú és sokáig csírázóképes növények.
- Ezek a fajok nem viselik el az erős kompetíciót, pl. árnyékolást, más fajok gyökereinek elszívó hatását.
- Az első betelepedő növények a termőtalaj fokozatos kialakulásának, valamint új és új fajok megjelenésének teremtik meg a lehetőségét.
- Diverzitás egyre nő, de a végén csökkenhet (pl. bükkerdő), a K-stratégiájúak a folyamat végén vannak jelen nagyobb arányban, melyek leárnyékolják a szukcesszió korábbi fázisainak fényigényes fajait. A folyamat során nagyobb magvú, nehezebben terjedő fajok nyernek tért.
- A folyamat addig tart, amíg az adott éghajlati viszonyok mellett lehetséges legbonyolultabb, legnagyobb produktivitású, egyensúlyban levő társulás, az ún. zárótársulás (klimaxtársulás) ki nem alakul (általában erdő).
- Planktoni szervezetek,
- lebegő hínárok (békalencse),
- gyökerező hínárok (békaszőlő, tündérrózsa, vízitök),
- nádas (káka, gyékény),
- magassásos társulások (zsombékosok),
- mocsárrétek,
- fűzbokor ligetek,
- többnyire fűz-nyár ligeterdő.
- A magas tápanyag-koncentráció, ami a túlzott műtrágyázás, a háztartási szennyvizek bemosódásának eredménye, nagymérvű fitoplankton burjánzáshoz vezethet (vízvirágzás).
- Ez az egész vízfelszínt elborítja, és meggátolja, hogy a fény az alsóbb vizeket is elérje.
- Ez megállítja a mélyebb rétegekben a növények növekedését és pusztulásukhoz vezet.
- Amikor a fitoplanktonok és a növények elpusztulnak, baktériumok lebontják őket. Ez a folyamat a vízben lévő oxigént felhasználja és nem marad elegendő az állatok lélegzésre, így azok elpusztulnak.
- A többlet tápanyagok néha hozzájárulhatnak olyan fitoplankton fajok elszaporodásához, amelyek káros méreganyagokat termelnek. Ezek sok faj pusztulását okozhatják. A kagylókban, rákokban felhalmozódó mérgek továbbkerülnek az emberbe.
- Az elpusztuló élőlénymaradványok a fenékre lesüllyedve anaerob, bomló szerves iszapot hoznak létre.
- A tó mélysége csökken, a nádasok egyre beljebb terjednek, elborítják a szabad vízfelszínt.
- Erdőirtás, fakitermelés,
- tavak kotrása,
- rétek legeltetése, kaszálása.
- Nyáras-borókás társulások létrejötte is emberi beavatkozásnak köszönhető.
- A legelés során az állatok a szúrós levelű borókát nem legelik le, a kialakuló borókák mellett a homokot kedvelő nyár is megtelepedhet.
- A degradáció legsúlyosabb esetben egy adott rendszer teljes kipusztulásához vezet, ilyen pl. erdőkben a tarvágás.
- A talaj leromlása helytelen talajművelés, túllegeltetés, túlzott öntözés, szikesedés, ill. eróziós vagy deflációs pusztulás következtében szintén degradációhoz vezet.
- A túlzott műtrágyázás miatt nitrofil gyomok megjelenése és az ennek nyomán fellépő növénytakaró-változás.
- amíg az anyagok – kémiai elemek - a körfolyamatban újra és újra felhasználódhatnak,
- a belépő energia csak egyszer haladhat át a rendszeren.
- Az élettelen környezet elemeit – C, H, O, N - az autotrófok veszik fel szervetlen anyagok formájában – CO2, H2O, ásványi anyagok - alakítják át szerves anyagokká és építik be a testükbe.
- egyrészt minden táplálkozási szint – még a növények is - egyedei lebontják – oxidálják – energianyerés céljából és visszaalakítják szervetlen anyagokká,
- másrészt az elpusztult élőlények megmaradt szerves anyagait a lebontók alakítják vissza szervetlen anyagokká.
- A szén minden szerves vegyület alapvázát képezi. A szén a levegőből szén-dioxid formájában kerül növényekbe (autotrófokba), ahol szén-dioxidból és vízből szerves anyagok keletkeznek. (1) A szerves anyag:
- a növényekben is lebomlik, szén-dioxidot felszabadítva (2),
- az elpusztult növényi részekkel a humusz alkotója lesz (3),
- továbbkerül valamely növényt fogyasztó állatba (4), abból pedig még tovább egy ragadozóba (5).
- Előbb-utóbb valamennyi - növényevő, ragadozó vagy lebontó - szervezet elbontja és felszabadítja a szerves vegyületekből a szén-dioxidot (6).
- A szén a humuszban igen sokáig kötött formában lehet jelen.
- Régi, földtörténeti időkben oxigénmentes környezetben keletkeztek a leülepedett szerves anyagokból a kőolaj-, a földgáz- és a kőszénkészletek, ezáltal hosszabb ideig kikerülhet a szén a körforgásból. Ezen fosszilis energiahordozókból égetéssel jut vissza a szén-dioxid a légkörbe (7).
- Továbbá a tengerek jelentős mennyiségű CO2-t nyelnek el, kivonva a légkörből, alapját képezve a karbonátos kőzetek – CaCO3 - képződésének.
- A vulkáni tevékenység is jelentős mennyiségű szén-dioxidot juttat a légkörbe.
- kis részét a fotoszintézishez használják fel, mely során elbontják hidrogénre és oxigénre.
- A hidrogén beépül a szerves vegyületekbe, az oxigén elemi formában (O2) a légkörbe kerül.
- A légköri oxigént az élőszervezetek a biológiai oxidációban használják fel, melynek során a szerves vegyületek H-tartalmát ismét vízzé alakítják.
- A főleg pillangósvirágú növények gyökérgümőiben – Rhizobium - szimbiózisban, és a talajban szaprofita módon élő aerob heterotróf nitrogénkötő baktériumok a légköri nitrogént megkötik és ammóniává alakítják. A folyamat energiaigényes.
-
- szerves vegyületekbe – aminosavakba – épül be, melyet a gazdanövény részben felhasznál,
- másrészt a talajba kerül (2).
- Az ammóniát kemoautotróf nitrifikáló baktériumok nitritionná, majd a nitritionokat nitrátionokká oxidálják. (3).
- Az ammónium- és a nitrátion vízben oldódik, és így a növények könnyen felveszik, s a nitrogént beépítik szerves vegyületekbe, pl. aminosavakba (4).
- A heterotróf szervezetek a növények által előállított szerves nitrogénvegyületek – pl. fehérjék - elfogyasztásával jutnak nitrogénhez (5).
- Az elpusztult élőlények nitrogéntartalmú szerves anyagaiból a lebontó baktériumok és gombák ammóniát állítanak elő, amelyből ismét ammóniumion és nitrátion keletkezhet (6).
- Oxigénmentes talajokban az anaerob vagy fakultatív anaerob denitrifikáló baktériumok hatására a nitrátionokból elemi nitrogén képződik, amely elszökik a levegőbe (nitrát-légzők). A denitrifikáló bacik többnyire anaerob heterotrófok, de ismertek kemoautotrófok is. A denitrifikáló baktériumok jelenléte káros a talaj nitrogéntartalmának szempontjából ezért a talaj szellőztetésével – kapálás – lehet ellenük védekezni (7).
- élettelen anyagok (víz, szén-dioxid, nitrogén, foszfor, különböző sók stb.),
- termelő vagy producens szervezetek, ezek szervetlen anyagokból szerves vegyületet állítanak elő (autotrófok),
- fogyasztók vagy konzumensek (heterotrófok),
- lebontók vagy reducensek (főleg baktériumok és gombák), az elhalt szervezetek összetett szerves anyagait bontják le (heterotrófok).
- a napenergia megkötését és áramlását szerves anyagok formájában,
- a táplálékláncokat,
- alakítja ki a bio-geo-kémiai ciklusokat.
- egy tó táplálékhálózatában egy édesvízi kagyló közvetlenül fogyaszthat zöldmoszatokat, amikor is ő egy elsődleges fogyasztó.
- Azonban táplálkozhat állati egysejtűekkel is, amelyek maguk elsődleges fogyasztók, ebben az esetben a kagyló másodlagos fogyasztó.
- termelő,
- a különböző szintű fogyasztó,
- valamint a lebontó szervezetek alkotják.
- környezeti problémát okoz, mivel a táplálékláncba bekerülve felhalmozódnak,
- a hasznos élőlényeket is elpusztítják,
- a legtöbb kártevőnél rezisztenciát alakítanak ki a vegyszerekkel szemben.
- rövidtávú hatás magában foglalja a heveny mérgezéseket, főleg májkárosodást,
- hosszútávú hatás, amelyről feltételezik, hogy szerepet játszik a rákos megbetegedések, a születési hibák, az immunológiai problémák, a Parkinson-kór és más krónikus elváltozások kialakulásában.
- Az autotrófok, elsősorban a növények fotoszintézissel történő szerves anyag előállítása az elsődleges vagy primer produkció.
- A fogyasztók és a lebontók – heterotrófok - által átalakított szerves anyag a másodlagos vagy szekunder produkció.
- egy része a szervezetekben végbemenő lebontó folyamatok révén elbomlik, energiát szolgáltatva az életműködésekhez,
- a másik része felhalmozódik az élőlényekben (beépül).
- az adott területen,
- adott időpontban megtalálható,
- élőlényekben előforduló szerves anyagok összes tömege.
- A fototrófok a szükséges energiát a Nap sugárzó energiájából nyerik,
- a kemoautotrófok – kemoszintetizálók - termelőként nem jelentősek, különböző szervetlen vegyületeket alakítanak át, és az átalakítás során keletkező kémiai energiát hasznosítják. Ilyenek pl. a nitrifikáló baktériumok, melyek ammóniát oxidálnak nitritté, ill. nitráttá, a nitrogén-körforgalomban van szerepük.
- részben energiaigényes folyamatokra fordítódik,
- részben hő formájában a környezetbe jut (~35%),
- kémiai energia – szerves anyagok - formájában áramlik a táplálkozási szinteken és
- hőenergia formájában hagyja el a rendszert minden szinten.
- amíg az anyagok – kémiai elemek - a körfolyamatban újra és újra felhasználódhatnak,
- a belépő energia csak egyszer haladhat át a rendszeren.
- ami nyílt anyagi rendszer, azaz amelyre jellemző az energia- és anyagáramlás és
- képes arra, hogy bizonyos határokon belül állandó értéken tartsa a belső tulajdonságait, homeosztázisa van, önszabályozó rendszer.
- Klímaváltozás
- A tengerek szintjének emelkedése, a vizek savasodása, tengeráramlatok megváltozása.
- Csapadékviszonyok megváltozása, elsivatagosodás, özönvízszerű esőzések.
- A Föld átlaghőmérsékletének emelkedése, globális felmelegedés.
- Jégsapkák, gleccserek olvadása.
- Ökoszisztémák fajösszetételének megváltozása (nem őshonos invazív fajok megjelenése).
- Környezetszennyezés
- Kommunális, ipari, mezőgazdasági eredetű.
- Talajok, levegő, vizek szennyezése.
- Fosszilis energiahordozók túlzott használata, a légköri CO2-koncentráció emelkedése.
- Ózonréteg elvékonyodása, UV sugárzás mértékének fokozódása.
- Hulladékkezelés, tárolás problémái, újrahasznosítás alacsony mértéke.
- Természetes élőhelyek csökkenése, eltűnése, erdőírtás, erdőtüzek.
- Biodiverzitás csökkenése, túlhalászat, fajok kihalása.
- Túlnépesedés, városiasodás.
- Éhezés, ivóvízhiány.
- Világjárványok kialakulása (pl. COVID19).
- Társadalmi konfliktusok (háborúk).
- a mezőgazdasági hozamok növekedése, javuló élelemzés,
- a jobb szociális környezet és egészségügyi ellátás, javuló higiénés körülmények,
- technológiai fejlődés,
- a járványok visszaszorítása,
- csecsemőhalálozás nagymértékű csökkenése,
- mindezek eredményeképpen a halálozási arány csökkent, a várható élettartam nőtt.
- Rossz minőségű ivóvíz, elégtelen higiénés feltételek.
- Hajléktalanság.
- Zaj- és légszennyezés. A városi stressz egyik oka a zajterhelés. A levegőszennyezés miatt egyre gyakoribbak a légzőszervi megbetegedések, például a hörghurut, az asztma és a tüdőrák.
- Túlzsúfoltság.
- Növekszik az agresszió, a bűnözés, a kábítószerfogyasztás,
- a nagyvárosi polgár elidegenedik a környezetétől,
- csökken a közösségi szellem, individualizálódik a társadalom, ami egyik fő oka a családok szétesésének.
- Szegregáció, a társadalmi csoportok területi elkülönülése.
- Csökken a természetes zöldterület.
- Nagyobb és koncentráltabb a szennyezőanyag- és hulladékkibocsátás.
- A közlekedés által okozott légszennyezés (szén-dioxid, szén-monoxid, nitrogén-oxidok, kén-dioxid, ólom, azbeszt), gyakran szmog (füstköd) kialakulásához vezet.
- A kibocsátott üvegházhatású gázok, ill. a jelentős felületű aszfalt erősen növeli a városok levegőjének hőmérsékletét.
- Továbbá fokozott zaj-, fény-, víz- és talajszennyezés.
- Növekszik a szennyvíz mennyisége, amelynek kezelése további problémákat vet fel.
- Sajátos nagyvárosi ökoszisztéma jön létre.
- megépítése nagy területet igényel,
- mikroklimatikus hatásaik és a gépkocsik gázkibocsátása nagyobb területet érint,
- felszíni hőmérsékletük jóval nagyobb, mint a növényzettel borított talajé, fokozódik a talajból a víz párolgása, ezért az út melletti talajrészek kiszáradnak.
- Az autópályák mentén jellegzetes zonáció figyelhető meg, az utak mellett szárazságtűrő, melegkedvelő egyéves fajok jelennek meg,
- a kozmopolita gyomfajok elszaporodnak és velük együtt kártevőik és fogyasztóik is terjednek.
- Az élőhelyek feldarabolódnak, a populációk elszigetelődhetnek (genetikai sodródás),
- az utak gyakran állatok vándorlási útvonalait keresztezik.
- a bioszférát felépítő ökoszisztémák
- és az ember alkotta építmények együttesei értendők.
- az ökológiai rendszerek működőképességének fenntartása,
- a természeti és épített környezeti értékek megőrzése,
- a környezetet szennyező és károsító kibocsátások csökkentése,
- a bekövetkezett károk felszámolása, a helyreállítás,
- valamint a természeti erőforrások fenntartható hasznosítása.
- a széndioxid-adó,
- a talajszennyezési adó,
- energiaadó,
- hulladékadó
- szennyvízadó.
- a gazdasági szereplők energiatakarékosságra történő ösztönzése,
- környezetbarát technológiák fejlesztése, használata,
- a természeti erőforrásokkal való takarékos gazdálkodás ösztönzése,
- a környezetkárosítás mérséklése,
- továbbá az adó okozta áremelkedés általában csökkenti a szóban forgó termék iránti keresletet.
- a társadalmi,
- a gazdasági és
- a környezeti pilléreken (lásd alább).
- A megújuló erőforrások kiaknázása nem lehet gyorsabb azok regenerálódási üteménél,
- a feleslegessé vált anyagok kihelyezésének mértéke nem haladhatja meg a környezet feldolgozó kapacitását,
- a nem megújuló erőforrások használatát a minimumra kell szorítani.
- Az erőforrásokat hatékonyan használjuk, a hulladékképződést minimalizáljuk.
- A szennyezést korlátozzuk azon a szinten, amely nem károsítja sem a természeti rendszereket, sem az emberi egészséget.
- Az emberek egészségét biztonságos, tiszta és kellemes környezet fenntartásával védjük, és a prevencióra helyezzük a hangsúlyt.
- A természet sokféleségét értékként kezeljük és óvjuk.
- A közösségek helyi igényeit helyileg elégítjük ki.
- Az áruk, a szolgáltatások a környezet megterhelése nélkül legyenek megközelíthetők.
- A passzív természetvédelem gondoskodik a védett értékek – fajok, területek - törvényes védelméről, de nem avatkozik bele a természetes folyamatokba.
- Az aktív – cselekvő – természetvédelem során az emberi beavatkozás következménye lesz a természetközeli állapot fennmaradása. Kezelés lehet pl.:
- kaszálás (réteken, legelőkön, ez a cserjésedést akadályozza meg),
- erdőművelés,
- nádaratás,
- vadászat,
- tavak kotrása, vízutánpótlás biztosítása stb.
- Védett területek létrehozása.
- Fajok védelme, biodiverzitás megőrzése.
- Kereskedelem korlátozása.
- Jogalkotás.
- nemzeti parkok (NP),
- tájvédelmi körzetek (TK),
- természetvédelmi területek (TT).
- a Hortobágyi,
- a Kiskunsági,
- és a Körös-Maros Nemzeti Park.
- Aggteleki,
- Bükki,
- Balaton-felvidéki,
- és az Őrségi Nemzeti Park.
- Fertő-Hanság,
- Duna-Dráva,
- Duna-Ipoly Nemzeti Park
- A környezeti problémák hatásos megoldásának hiánya többletköltséggel jár.
- A tiszta technológiákba való beruházások új piacokat és munkahelyeket teremtenek. Társadalmaink ökológiai modernizációja épp ezért a gazdasági modernizáció szerves része. Forrás: Dr. Damjanovich Imre
- Ramsari egyezmény- a nemzetközi jelentőségű vizes területekről, 1971.
- Rioi egyezmény - a biológiai sokféleségről, Rio de Janeiro, 1992.
- Kiotói jegyzőkönyv - ENSZ keretegyezmény a klímaváltozásról, Kyoto 1997.
- struktúrgéneket (lacZ, lacY, lacA), amelyek olyan fehérjék aminosavsorrendjét kódolják, amik lehetővé teszik a laktóz felvételét és lebontását,
- továbbá régió szakaszokat, amelyekről nincs átíródás – nem kódolnak fehérjét -, ezek a struktúrgének átíródását lehetővé tevő – szabályozó - egységek.
- három struktúrgén (lacZ, lacY, lacA),
- promoter (lacP), az RNS-polimeráz kapcsolódási helye,
- operátor régió (lacO), egy olyan DNS-szakasz, ahol egy gátlófehérje (represszor) képes megkötődni, megakadályozva a fenti struktúrgének átírását, abban az esetben, ha nincs laktóz a tápanyagok között.
- a gátlófehérje az operátorhoz kapcsolódik és megakadályozza a struktúrgének átírását.
- a gátlófehérje megköti a laktózt, szerkezete megváltozik, leválik az operátorról és nem képes tovább akadályozni az RNS-polimerázt a struktúrgének átírásában.
- Eredményül átíródnak a struktúrgének, fokozódik a laktóz felvétele, lebontása (a szabályozás negatív visszacsatolás, mivel a laktózkoncentráció növekedését, annak csökkenése követi).
- a homológ kromoszómák véletlenszerű szétválása,
- az átörökítő DNS átrendeződése (intrakromoszómális rekombináció),
- az ivarsejtek véletlenszerű találkozása.
- ha a mutáció nem kódoló, öröklődés szempontjából közömbös szakaszt érint,
- vagy információt hordozó szakaszt érint, azonban a genetikai kód degeneráltsága miatt, egy báziscsere nem változtatja meg az aminosavsorrendet, tekintve, hogy a megváltozott kodon ugyanazt az aminosavat kódolja (csendes mutáció).
- fizikai okok:
- a nagy energiájú sugárzások roncsolják a DNS-t,
- UV, rtg, radioaktív sugárzás.
- Hőmérséklet emelkedés.
- a nagy energiájú sugárzások roncsolják a DNS-t,
- Kémiai hatások.
- Biológiai tényezők, pl. baktérium- és gombatoxinok és más természetes eredetű növényi vagy mikrobiális eredetű anyagok.
- fajonként eltérő,
- diploid szervezetekben gyakoribb,
- egy fajon belül génenként változó,
- emberben átlagosan 10-5 nagyságrendű.
- Pontmutáció (egy vagy néhány nukleotidot érint).
- Kromoszómamutációk (egy vagy több kromoszómát érint).
- Kromoszóma szerkezeti változások.
- Kromoszóma számbeli változások.
- Aneuploid, ill,
- euploid genommutáció.
- kicserélődhet egy bázis és így pl. stopjellé alakul.
- Kieshetnek, ill. beépülhetnek bázisok (frame shift mutációk), ami az egész leolvasási keret eltolódásához vezet.
- kromoszómarészek átrendeződését,
- a kromoszómák számának megváltozását vagy
- az egész kromoszómaszerelvény – genom - számának változását eredményezik (ploidiaszint változás).
- Deléció: egy kromoszómadarab kiesése.
-
- Cri du chat – macskasírás szindróma.
- A macskasírás szindróma estében az 5. kromoszóma rövid karjának vége hiányzik, kisfejűséggel, holdszerű arccal, macskanyávogásszerű hanggal és szellemi elmaradottsággal jár.
- Duplikáció: egy kromoszóma régió megkétszereződése.
- Inverzió: egy kromoszóma szakasz 180 fokos átfordulása az adott kromoszómán belül.
- Transzlokáció: két nem homológ kromoszóma közötti részek (kölcsönös) áthelyeződése.
- az ivarsejtek,
- a spórák, a növények ivaros nemzedékének a sejtjei,
- az algák, mohák testi sejtjei,
- a hím méhek, darazsak, hangyák. Ezek az állatok szűznemzéssel, azaz megtermékenyítetlen petéből jönnek létre. Ivarsejtjeiket mitózissal hozzák létre.
- mongoloid szemrés,
- holdvilág arc,
- nagy nyelv,
- 50 körüli IQ,
- négyujjas tenyérbarázda,
- szívhibák,
- továbbá sokféle belső fejlődési rendellenesség.
- Az 5’ véget a nukleázokkal szemben az ún. sapka (cap) védi.
- Továbbá közvetlenül a transzkripció után, az RNS 3'-végéhez kb. 150-200 adenin nukleotidot tartalmazó, információt nem hordozó poli-A farok kapcsolódik.
- intron (intragén) szakaszokat, melyek az érett messengerből kivágódnak, és tartalmaznak
- exonokat, amelyek a gének kifejeződő szakaszai.
- Egyetlen nukleotid tehát nem lehet jel – kodon -, mert így csupán 4-féle aminosav beépülését lehetne szabályozni.
- Ha két-két nukleotid alkotna egy-egy jelet, akkor is csak 42 = 16-féle kodon alakulhatna ki.
- A 3 nukleotidból álló egységek már 43 = 64-féle kombinációt hoznak létre, ami bőségesen elegendő a 20-féle aminosav kódolásához.
- Az AUG-kodon a transzláció start jele (egyben a metionin kodonja).
- Három kodon jelzi a fehérjeszintézis végét, ezeket a tripleteket stop kodonnak nevezzük, ezek nem kódolnak aminosavakat.
- Az adott aminosavval összekapcsolódik, s mivel antikodon része komplementer a megfelelő kodonnal, megfelelteti egymásnak az aminosavat és az aktuális kodont.
- A polipeptidlánc szintézise energiát igényel, ezért előzőleg az aminosavakat magasabb energiaállapotba kell hozni, azaz aktiválni kell. Az aktiválás során ATP közreműködésével az aminosavak nagy energiájú kötéssel kapcsolódnak a tRNS-molekulák 3’ végéhez.
- Csak homozigóta formában jelentkezik a hatás,
- a jelleg megjelenése egy-egy generációt átugorhat.
- Az ilyen betegségek mindkét nemet azonos valószínűséggel érintik.
- A beteg gyermek születésének a valószínűsége tünetmentes hordozó szülőktől 25%.
- A rokonházasságok nagymértékben megnövelik az autoszomális recesszív betegségek veszélyét.
- Fenilketonúria
- Albinizmus
- Sarlósejtes anémia
- Cisztás fibrózis
- Galaktózémia
- A homozigóta recesszívek sarlósejtes vérszegénységben halnak meg,
- a homozigóta dominánsok pedig maláriában.
- Heterozigótákban a kóros hemoglobin védelmet biztosít a malária kórokozójával szemben, így ők nem betegednek meg sem maláriában, sem sarlósejtes vérszegénységben.
- Az öröklésmenetben minden nemzedékben megjelenik a
- Betegség esetén egyaránt érintett lehet mindkét nem azonos valószínűséggel,
- egészséges szülőknek nem lehet beteg gyermeke, de beteg utódnak legalább egyik szülője mindig beteg.
- A betegséget mutató egyén többnyire heterozigóta, mert a homozigóták sokszor életképtelenek.
- Az érintett szülőnek 50% az esélye, hogy továbbörökítse betegségét.
- Leginkább a csontrendszerrel kapcsolatos rendellenességek ilyenek.
- Achondroplasia
- Marfan-szindróma
- Polydactylia - sokujjúság
- Huntington-szindróma
- Megrövidült végtagok.
- Nagy fej kiemelkedő homlokkal és lapos orrnyereggel.
- Rendezetlen fogsor.
- Görbe lábszár.
- Normál intelligencia.
- Jóval több érintett férfi, mint nő (a férfiaknak csak egy X-kromoszómájuk van).
- Érintett nők homozigóta recesszívek (XX), az érintett férfiak hemizigóták (XY).
- Heterozigóta – hordozó - nőknél enyhébb betegségtünetek jelentkezhetnek, az X-kromoszóma véletlen inaktivációja révén.
- Hordozó nők 50%-os eséllyel adják tovább a recesszív állélt.
- Érintett férfiak anyja és lányai tünetmentes hordozók lehetnek, fiai mind egészségesek.
- A fenotípus tehát egy-egy generációt “átugorhat”.
- Érintett nők (ritka!) apja és valamennyi fia is érintett, lányai tünetmentes hordozók.
- Vörös-zöld színtévesztés
- Hemofília
- Fogzománc-elszíneződés
- Öröklött látóidegsorvadás
- Öröklött farkasvakság
- Izomsorvadás
- hemofília A: VIII. faktor hiánya, ill.
- hemofília B: IX. faktor hiánya, az X-kromoszómához kötött öröklődő betegségek.
- A családfa az autoszomális dominánshoz hasonló lefutású, azonban a két nem érintettsége nem egyforma.
- Kétszer annyi nő érintett, mint férfi (2 X-kromoszóma).
- Jellemző az X-hez kötött hím letalitás. Mivel a férfiak hemizigóták, ezért nincs normál allél, így a hímnemű embriók már a méhen belül elhalnak.
- Az érintett nők tünetei sokszor enyhébbek és változékonyabbak, mint a beteg férfiaké.
- Egy érintett nő utódainak 50%-a – tekintet nélkül nemükre – beteg.
- Egy érintett férfi minden leánya beteg, minden fia egészséges (az apa leányainak mindig az X-kromoszómáját adja, fiainak mindig az Y-t!)
- Ilyen betegség a hipofoszfatémia (régebbi nevén D-vitamin-rezisztens angolkór), amelyet növekedési visszamaradottság, angolkór és alacsony vérfoszfátszint jellemez.
- Érintett férfiak apja is érintett.
- Érintett férfiak valamennyi fia is érintett.
- A fenotípus minden generációban megjelenik.
- az osztódás fázisára,
- az osztódások közötti ún. interfázisra.
- Az interfázisos sejtben
- a maghártya élesen elkülöníti a mag állományát,
- melynek belsejét a kromatinállomány tölti ki.
- Az osztódó sejtben
- a maghártya feldarabolódik, majd eltűnik,
- a kromatinállomány kromoszómákká tömörödik, a kromoszómák jól felismerhetőek.
- G1 szakasz, (G, angol gap = (idő)rés)
- S szakasz,
- G2 szakasz.
- növekednek,
- a sejtszervecskék megsokszorozódnak,
- lemerült energiaraktáraikat feltöltik.
- a replikáció, melynek során 46 DNS -molekulából - melyek egykromatidás kromoszómák - 92 DNS-molekula lesz (46 kétkromatidás kromoszóma),
- tehát létrejönnek a kromoszómák másik kromatidái.
- Rövid nyugalmi szakasz, amelyben a sejt létrehozza azokat az anyagokat, amelyek az osztódáshoz kellenek (pl. húzófonalak fehérjéit). Mind a G1, mind a G2 szakaszra intenzív fehérjeszintézis jellemző.
- A sejtközpont kettéosztódik, a sejt két pólusára vándorol.
- a protoonkogének, melyek olyan fehérjéket kódolnak, amik növekedési jeleket fognak fel a sejtekben. E géneknek mutációi eme fehérjék aktivációját idézik elő, aminek hatására az érintett sejt úgy érzékeli, mintha a növekedési jelzés folyamatosan jelen volna. Az aktivált protoonkogéneket onkogéneknek nevezzük.
- Tumor-szuppresszorok, daganatelnyomó gének fehérjéi működésük során féken tartják a sejtosztódást, a sejtciklust hiba esetén leállítják.
- A DNS-hibajavítók és
- az apoptózis-szabályozók (sejthalálgének), amelyek olyan fehérjéket kódolnak, melyek a programozott sejthalált beindítják.
- egyrészt környezeti tényezők – pl. karcinogén (rákkeltő) hatások - okozzák a rákhoz vezető mutációk felhalmozódását, melyek kialakulását öröklött genetikai tényezők is növelhetik.
- Másrészt tisztán örökletes, genetikai okok játszanak egyértelműen meghatározó szerepet a rák kialakulásában. A rosszindulatú daganatok nagyjából öt-tíz százaléka tartozik ebbe a csoportba.
- nem törnek be a környező szövetekbe, és nem pusztítják el őket,
- egy kötőszövetes hártya borítja a sejthalmazt, amely elszigeteli a környezettől,
- a belőlük származó sejtek nem szóródnak szét a szervezetben, nem képeznek áttétet,
- általában lassan növekednek, rendszerint eltávolíthatók, gyakran nem újulnak ki,
- és ami a legfontosabb, a jóindulatú daganatok általában nem veszélyeztetik az életet.
- Ha a jóindulatú daganat például belső elválasztású mirigyekben fejlődik, akkor az általuk termelt hormon szintje jelentősen megemelkedhet, ami hormonális zavarokhoz is vezethet.
- a daganatos sejtek kontrollálatlanul és megállíthatatlan módon osztódnak.
- A tumort szinte soha nem borítja határoló hártya, és ezért
- a tumorsejtek betörve a környező szövetekbe (orvosi kifejezéssel ezt beszűrődésnek, infiltrációnak nevezik) áttéteket képeznek.
- Amint a daganat növekszik, megkezdi saját vérellátásának kiépítését. A tumorsejtek elveszik a táplálékot az egészséges sejtek elől, a beteg gyengül, és fogyásnak indul.
- Az ún. karcinómák hámszövetben fejlődnek ki. A bőrön kívül ide tartoznak a szervek belső falát burkoló sejtrétegek is, például a bélcső üregének hámborítása.
- Az ún. szarkómák kötőszövetekből indulnak ki.
- Klasszikus genetikai génfogalom: a gén egy olyan anyagi egység, amely egy tulajdonság kialakulását határozza meg.
- Molekuláris genetikai génfogalom: első megközelítésben, a gén a DNS-molekula egy szakasza, amely egy polipeptidlánc aminosavsorrendjét kódolja.
- Ezenkívül vannak t- és rRNS-, ill. miRNS-gének is.
- struktúrgén: olyan DNS-szakasz, amely RNS-be átíródhat:
- vagy egy polipeptidlánc aminosavsorrendjét határozza meg (mRNS),
- vagy r- és tRNS-, ill. miRNS-molekulákat kódol,
- szabályozó régió, amely nem íródik át, a struktúrgének működését, átíródását szabályozza (pl. operátor régió, lásd lac operon).
- Kromoszómaszerelvény (kromoszóma-alapszám): a fajra jellemző kromoszómák egyszeres sorozata, pl. ember esetén 23 darab kromoszóma (a haploid sejtek kromoszómaszáma, genomja).
- Haploid sejtek/szervezetek egyszeres kromoszómaszerelvényűek (pl. ivarsejtek, spórák, mohák testi sejtjei, mohák, harasztok előtelepének sejtjei).
- Genom: a haploid sejtek egyszeres genetikai állománya (n), egy ivarsejt genetikai anyagának az összessége, egyszeres kromoszómaszerelvény.
- Diploid sejtek/szervezetek kétszeres genomot (2n), azaz kettős kromoszómaszerelvényt tartalmaznak (pl. általában a testi sejtek).
- Poliploid sejtek többszörös – 3n, 4n - kromoszómaszerelvényt tartalmaznak (pl. a termesztett növények egy része).
- Az allél (génváltozat) a kromoszóma egy adott szakaszán (lókuszán) elhelyezkedő gén variációja.
- Homozigóta egyedben az adott két allél teljesen azonos, jelölésük: pl. AA vagy aa.
- Heterozigóta egyedben az adott két allél különbözik, jelölése: pl. Aa.
- A domináns – uralkodó - allél hatása elnyomja a recesszív allélt, heterozigótákban is kifejeződik,
- a recesszív – lappangó - allél csak homozigóta formában tudja kifejteni hatását.
- Kodominancia esetén mindkét allél kifejeződik, nem nyomják el egymás hatását. Ilyen pl. az emberi vércsoportoknál figyelhető meg. Az "A" vércsoportért felelős allél és a "B" vércsoportért felelős allél egyszerre is képes kifejeződni, ekkor "AB" vércsoport alakul ki (vagy még ilyen az MN vércsoport öröklődése).
- Genotípus: a jelleg létrehozását biztosító genetikai szerkezet, a tulajdonság meghatározásának az anyagi alapja, a DNS adott szakaszának minősége, adott génre nézve lehet homozigóta domináns vagy recesszív, ill. heterozigóta.
- Fenotípus: a látható megjelenési forma.
- Fenom: a megfigyelhető tulajdonságok, a fenotípusok összessége.
- A genetika tudományának megalapítója.
- Osztrák középiskolai szerzetestanár.
- Gondos keresztezési kísérleteket végzett a kolostora kertjében 1857 és 1868 között.
- Sok adatot gyűjtött, statisztikailag értékelte az eredményeket.
- Sok változata van.
- A növény öntermékenyítő, de könnyen keresztezhető
- Rövid tenyészidejű.
- Sok utódot hoz létre.
- Mendel csupán néhány kiragadott – szám szerint 7-et - tulajdonságot vizsgált.
- Borsószem alakja, ami lehet kerek és szögletes, felülete sima vagy ráncos.
- Borsószem színe lehet sárga és zöld.
- A virág színe lehet bíbor és fehér.
- Hüvely színe lehet zöld és sárga.
- Stb.
- kis testméretű,
- egyszerűen fenntartható,
- rövid generációs idejű,
- sok utódot hoz létre,
- és könnyen keresztezhető.
- vírusok,
- baktériumok (pl. Escherichia coli),
- élesztőgombák,
- kerti borsó,
- ecetmuslica (Drosophila melanogaster),
- egér.
- Különböző tulajdonságokat mutató (pl. bíbor és fehér virág) tiszta – homozigóta, egyforma egyedekből álló - vonalak kiválasztása (P, parental = szülői nemzedék).
- Az eltérő fenotípusú – lila, fehér - vonalak keresztezése mesterséges beporzással.
- Majd az utódok egymás közötti további keresztezése. (F, filial = utódnemzedék)
- A vizsgált tulajdonságot egyetlen gén két allélja határozza meg, melyek domináns-recesszív viszonyban állnak.
- Ezen gén domináns allélja okozza az F1 fenotípust (bíbor színt),
- a recesszív fenotípus (fehér virág) csak homozigóta formában, F2-ben jelentkezik.
- Az öröklődést ún. faktorok határozzák meg (ezeket ma géneknek nevezzük).
- Minden egyed egy faktorpárt - azaz allélpárt - hordoz egy adott tulajdonságra.
- Az ivarsejtek képződése során a faktorpárok – allélpárok - tagjai szétválnak egymástól, és külön ivarsejtekbe jutnak, melyek így az allélpároknak csak egyik tagját hordozzák (meiózis).
- Az ivarsejtek zigótává egyesülése véletlenszerű, nem függ a hordozott faktorok -gének - természetétől.
- A gamétatisztaság szabálya
- Az uniformitás szabálya (Mendel I. tv.)
- homozigóta, úgy az utódok kizárólag domináns fenotípusúak lesznek,
- azonban, ha heterozigóta, akkor fele-fele arányban kapunk domináns, ill. recesszív fenotípusú utódokat.
- A reciprocitás szabálya
- A szegregáció – hasadás - szabálya (Mendel II. tv.)
- Független öröklődés, szabad kombinálódás szabálya (Mendel III. tv.)
- mag színe: sárga (A) és zöld (a),
- mag alakja: sima (B) és ráncos (b).
- Az F1 egyedei a homológ kromoszómapárok tagjainak a véletlenszerű szétválása miatt – interkromoszomális rekombináció – 4-féle ivarsejtet (AB, Ab, aB, ab) képeznek azonos valószínűséggel (25%).
- Tekintve, hogy az ivarsejtek találkozása véletlenszerű, bármelyik hímivarsejt bármelyik petesejtet megtermékenyítheti, F2 nemzedék egyedei 4x4-féle kombinációban jöhetnek létre.
- Minden lehetőséget figyelembe véve F2-ben 9:3:3:1 fenotípusarányt kapunk.
- A magalak génje a magszín génjétől tehát függetlenül öröklődik.
- F2 nemzedékben a két tulajdonság szabadon kombinálódott, hiszen létrejöttek mind a szülőktől, mind az F1 nemzedéktől különböző, új tulajdonságú egyedek, mint pl. sárga-ráncos, ill. zöld-sima.
- Minden mendeli allélpár domináns-recesszív viszonyt mutatott (teljes dominancia).
- Mindegyik allélpár testi kromoszómán volt.
- Mindegyik vizsgált allél életképes volt homozigóta formában.
- Minden vizsgált gén csak egyetlen fenotípust befolyásolt.
- Egyetlen tulajdonságot egyetlen allélpár határozott meg.
- Minden mendeli fenotípus génje különböző kromoszómán volt található.
- Mindegyik vizsgált fenotípusa jól azonosítható volt, mert nem függött a környezeti hatásoktól és a genetikai háttértől.
- A dominanciaviszonyok lehetséges változatai
- A nem teljes dominancia (inkomplett dominancia), intermedier öröklésmenet
- Kodominancia (MN, ABO vércsoport A és B alléljeinek viszonya)
- Letális allélek (életképtelen homozigóta genotípusok)
- Egyetlen gén többféle fenotípusos jellegre hat (pleiotrópia)
- Több gén befolyása ugyanarra a jellegre (génkölcsönhatások)
- Nemhez kötött öröklődés
- Extranukleáris öröklődés (anyai öröklődés) és az anyai hatás
- Kapcsolt öröklődés, amikor a jellegeket meghatározó allélek azonos kromoszómán vannak.
- az MN vércsoport öröklődése, ill.
- az ABO vércsoport esetén az A és a B allél viszonya.
- Ha a heterozigóta egyed fenotípusa megegyezik az egyik allélra homozigóta egyed fenotípusával, akkor az az allél teljes dominanciát mutat a másik felett. Ez esetben az a domináns géntermék egy dózisa elég a dupla géndózisnak megfelelő fenotípus kialakításához. A másik allél ez esetben recesszívként viselkedik.
- Az inkomplett dominanciánál (intermedier öröklésmenet) az allélek dupla, szimpla és nulla dózisa más-más fenotípust okoz.
- Kodominanciánál az allélok egymással egyenértékűek, hatásuk egyszerre megnyilvánul.
- Az IA és IB allél két különböző antigén meghatározásáért felelős,
- az i allél nem kódolja sem az A, sem a B antigént, azaz jelenléte az A és a B antigén hiányát okozza (0-s vércsoport).
- Teljes dominancia IA ill. IB és i között,
- kodominancia IA és IB között áll fenn.
- egy adott gén allélváltozatai, ekkor allélikus kölcsönhatásokról beszélünk, ill.
- különböző géneknek kölcsönhatásai, ekkor génkölcsönhatásokról szólunk.
- Intrakromoszomális – kromoszómán belüli rekombináció. A meiózis első főszakaszában történik, a homológ kromoszómák egy-egy kromatidája között. Intrakromoszomális rekombináció esetén a rekombinánsok aránya kevesebb, mint 50 százalék.
- Az interkromoszomális rekombináció szintén az első főszakaszban zajlik a homológ kromoszómák véletlenszerű szétválásának köszönhetően. Az interkromoszomális (független) rekombináció mindig 50%-os rekombinációs gyakoriságot eredményez (vagy-vagy).
- Két gén között 50% a rekombinációs gyakoriság, ha azok nem kapcsoltak.
- Ha a kapcsolt gének a kromoszóma ellentétes végein helyezkednek el, közöttük a rekombináció gyakorisága az 50%-hoz közeli érték, látszólag úgy viselkednek, mint a nem kapcsolt gének.
- egyrészt megkülönböztetjük a testi kromoszómákat – autoszómák -,
- másrészt az ivarmeghatározásért felelős ún. ivari kromoszómákat - gonoszómák.
- a nőstények két egyforma (XX; homogaméta),
- a hímek pedig két különböző (X,Y heterogaméta) ivari kromoszómával rendelkeznek.
- Nőkben két egyforma XX,
- férfiakban az egyik kisebb, az Y-, a másik olyan, mint a nők X-kromoszómája.
- Minden petesejt X-tartalmú, azonban
- a spermiumok fele X-, másik fele Y-kromoszómát hordoz.
- egy ivarsejtbe jut mind a két ivari kromoszóma, és így
- a másik ivarsejt ivari kromoszóma nélküli lesz.
- a női jellegű XX genotípusú zigótában az egyik X-kromoszóma a petesejtből (anyai kromoszóma), a másik X-kromoszóma a hímivarsejtből (apai kromoszóma) származik,
- a hím jellegű zigótában az X-kromoszóma a petesejtből, az Y-kromoszóma pedig a hímivarsejtből eredeztethető.
- beteg anyának nem lehet egészséges fia,
- beteg lánynak nem lehet egészséges apja.
- Jóval több érintett férfi, mint nő (a férfiaknak csak egy X-kromoszómájuk van).
- Érintett nők homozigóta recesszívek (XX), az érintett férfiak hemizigóták (XY).
- Heterozigóta – hordozó - nőknél enyhébb betegségtünetek jelentkezhetnek, az X-kromoszóma véletlen inaktivációja révén.
- Hordozó nők 50%-os eséllyel adják tovább a recesszív állélt.
- Érintett férfiak anyja és lányai tünetmentes hordozók lehetnek, fiai mind egészségesek.
- A fenotípus tehát egy-egy generációt “átugorhat”.
- Érintett nők (ritka!) apja és valamennyi fia is érintett, lányai tünetmentes hordozók.
- Vörös-zöld színtévesztés
- Hemofília
- Fogzománc-elszíneződés
- Öröklött látóidegsorvadás
- Öröklött farkasvakság
- Izomsorvadás
- A családfa az autoszomális dominánshoz hasonló lefutású, azonban a két nem érintettsége nem egyforma.
- Kétszer annyi nő érintett, mint férfi (2 X-kromoszóma).
- Jellemző az X-hez kötött hím letalitás. Mivel a férfiak hemizigóták, ezért nincs normál allél, így a hímnemű embriók már a méhen belül elhalnak.
- Az érintett nők tünetei sokszor enyhébbek és változékonyabbak, mint a beteg férfiaké.
- Egy érintett nő utódainak 50%-a – tekintet nélkül nemükre – beteg.
- Egy érintett férfi minden leánya beteg, minden fia egészséges (az apa leányainak mindig az X-kromoszómáját adja, fiainak mindig az Y-t!)
- Ilyen betegség a hipofoszfatémia (régebbi nevén D-vitamin-rezisztens angolkór), amelyet növekedési visszamaradottság, angolkór és alacsony vérfoszfátszint jellemez.
- Kizárólag férfiak érintettek.
- Érintett férfiak apja is érintett.
- Érintett férfiak valamennyi fia is érintett.
- A fenotípus minden generációban megjelenik.
- Az IA és IB allél két különböző antigén meghatározásáért felelős,
- az i allél nem kódolja sem az A, sem a B antigént, azaz jelenléte az A és a B antigén hiányát okozza (0-s vércsoport).
- Teljes dominancia IA ill. IB és i között,
- kodominancia IA és IB között áll fenn.
- A domináns allél az RH-faktort (D-antigén) kódolja, így az RH+ vércsoport meghatározásáért felelős,
- a recesszív allél pedig az RH-faktor hiányát eredményezi.
- az RH+ fenotípusú személyek vagy homozigóta domináns, vagy heterozigóta genotípusúak lehetnek,
- az RH- emberek pedig csak homozigóta recesszív genotípusúak lehetnek.
- a fajon – populáción - belül az egyes tulajdonságok gyakoriságának megváltozásában,
- új tulajdonságok megjelenésében nyilvánul meg.
- a szaporodás,
- az öröklődés
- és a változatosság.
- A jelleg átlagától valamilyen irányban eltérő tulajdonságnak kedvez.
- A szélsőségeket részesíti előnyben.
- száraz környezetben a keskenylevelű fűfajok a rátermettebbek.
- Mesterséges szelekciónál (nemesítésnél, állattenyésztésben, növénytermesztésben) gyakran alkalmazott módszer.
- A szélsőértékek kerülnek szelekciós előnybe az átlaggal szemben.
- Az átlagértékek kerülnek szelekciós előnybe.
- Természetben a leggyakoribb.
- Pázsitfűfélék levelének szélessége átlagos környezetben. A széleslevelűek könnyen kiszáradnak, a keskenylevelűek kevésbé hatékonyan fotoszintetizálnak.
- Domináns letális allél esetén a szelekció rögtön megvalósul. Ugyanakkor a hibás allél a mutációs rátájának megfelelően újraképződhet.
- Recesszív letális allél esetén csak a homozigóták (aa) szelektálódnak ki rögtön, a heterozigótákban tovább lappanghat az allél. A nemzedékek során az allél gyakorisága csökken, előfordulási valószínűsége közelít a 0-hoz.
- A homozigóta recesszívek sarlósejtes vérszegénységben halnak meg,
- a homozigóta dominánsok pedig maláriában.
- Heterozigótákban a kóros hemoglobin védelmet biztosít a malária kórokozójával szemben, így ők nem betegednek meg sem maláriában, sem sarlósejtes vérszegénységben.
- A virágok és az őket beporzó rovarok kapcsolata.
- A paraziták és a gazdaszervezetek kölcsönhatásai.
- A ragadozók és a prédaállatok felépítése és viselkedése.
- Növények és növényevők együttes fejlődése.
- Szimbióta szervezetek alkalmazkodása.
- A lombfakadás és a lombfogyasztó rovarok megjelenésének összehangoltsága.
- Vulkanikus szigetek, melyekre néhány egyed véletlenül vetődött.
- Először azokat az élőhelyeket foglalhatták el, mint a kontinensen,
- majd mikor azok telítődtek, új, üres élőhelyeket népesítettek be versengés nélkül.
- Ma 14 pintyfaj él a szigeteken az eredeti magevőkön kívül - virágevő, rovarevő -, melyek alkalmazkodtak az új környezethez, amit az eltérő csőrszerkezetük mutat.
- a rovarok és a gerincesek szárnya,
- a vakond és a lótücsök ásólába,
- a halak és a cetek úszói.
- a fajon – populáción - belül az egyes tulajdonságok gyakoriságának megváltozásában,
- új tulajdonságok megjelenésében nyilvánul meg.
- fajok közötti diverzitás, ami a különböző fajok eltérő génállományának, ill. fenotípusos megjelenésének a sokfélesége,
- fajon belüli, populációk közötti diverzitás, ami a faj egyes populációinak eltérő genetikai összetételét fejezi ki, hiszen egy adott allél előfordulási gyakorisága eltérő lehet a különféle populációkban, mint pl. az AB0 vércsoportot meghatározó allélgyakoriság-eltérés az egyes emberi populációk között,
- egy populáció egyedei közötti genetikai sokféleség, ami a populáción belül fellelhető allélváltozatok egyedekben való előfordulásától függ, pl. az AB0 vércsoportrendszerben 4-féle vércsoportot különböztetünk meg annak alapján, hogy a populáción belül a vércsoportokat meghatározó 3-féle allél – IA, IB, i – milyen párt alkotnak az egyedekben,
- egy egyed DNS-ében fellelhető genetikai diverzitás, ami a genomon belül az egyes génpárokat alkotó allélok változatosságát jellemzi, azaz az egyes génekre nézve mekkora arányban jelentkezik a heterozigóta genotípus (ha minden allélpár heterozigóta genotípusú, akkor szélsőségesen nagy, amennyiben minden allélpár homozigóta genotípusú, akkor szélsőségesen alacsony a diverzitás mértéke).
- A jelleg átlagától valamilyen irányban eltérő tulajdonságnak kedvez.
- A szélsőségeket részesíti előnyben.
- száraz környezetben a keskenylevelű fűfajok a rátermettebbek.
- Mesterséges szelekciónál (nemesítésnél, állattenyésztésben, növénytermesztésben) gyakran alkalmazott módszer.
- A szélsőértékek kerülnek szelekciós előnybe az átlaggal szemben.
- Az átlagértékek kerülnek szelekciós előnybe.
- Természetben a leggyakoribb.
- Pázsitfűfélék levelének szélessége átlagos környezetben. A széleslevelűek könnyen kiszáradnak, a keskenylevelűek kevésbé hatékonyan fotoszintetizálnak.
- Domináns letális allél esetén a szelekció rögtön megvalósul. Ugyanakkor a hibás allél a mutációs rátájának megfelelően újraképződhet.
- Recesszív letális allél esetén csak a homozigóták (aa) szelektálódnak ki rögtön, a heterozigótákban tovább lappanghat az allél. A nemzedékek során az allél gyakorisága csökken, előfordulási valószínűsége közelít a 0-hoz.
- A homozigóta recesszívek sarlósejtes vérszegénységben halnak meg,
- a homozigóta dominánsok pedig maláriában.
- Heterozigótákban a kóros hemoglobin védelmet biztosít a malária kórokozójával szemben, így ők nem betegednek meg sem maláriában, sem sarlósejtes vérszegénységben.
- nem adaptív jellegűek, melyek sokszor hirtelen, véletlenszerűen, előre nem látható módon következnek be, mint pl. a mutáció, genetikai sodródás, migráció,
- a változás hátterében adaptív folyamatok is állhatnak, a populációk a szelekciós folyamatok következtében lassan, fokozatosan alkalmazkodnak a környezetükhöz, melynek során az arra alkalmas genotípusok kiválogatódnak.
- egy faj átalakul (anagenezis, nem jön létre új elágazás),
- egy fajból szétválással több képződik (kladogenezis, új elágazás jön létre, ahol az egyik faj megtarja eredeti vonásait).
- Ebben az esetben a véletlenszerű szétválás miatt elsősorban a genetikai sodródás hatására képződnek új fajok.
- Ilyen izoláció például a kontinensek vándorlása, a folyók kialakulása, ill. az egymással kapcsolatban nem álló édesvízi tavak vagy az óceáni szigetek élővilágának kialakulása során ment végbe.
- a földrajzi izoláció következtében alakulhat ki.
- Eltérővé válhat a szaporodási időszak.
- A szaporodási szokások megváltozhatnak.
- A fajok populációi ugyanazon földrajzi területen különböző élőhelyeket foglalnak el, és így általában nem találkoznak egymással. Az eltérő ökológiai környezethez való alkalmazkodás eredményeképpen az elkülönülő populációrészek egyedei sajátos úton fejlődhetnek.
- Vulkanikus szigetek, melyekre néhány egyed véletlenül vetődött.
- Először azokat az élőhelyeket foglalhatták el, mint a kontinensen,
- majd mikor azok telítődtek, új, üres élőhelyeket népesítettek be versengés nélkül.
- Ma 14 pintyfaj él a szigeteken az eredeti magevőkön kívül - virágevő, rovarevő -, melyek alkalmazkodtak az új környezethez, amit az eltérő csőrszerkezetük mutat.
- a rovarok és a gerincesek szárnya,
- a vakond és a lótücsök ásólába,
- a halak és a cetek úszói
- nem válnak szét a meiózis során, így ugyanabba az ivarsejtbe kerül mindkét homológ kromoszóma, vagy
- megkezdik a sejtek a mitózisukat, azonban a szétváló kromatidák ugyanabba a sejtbe kerülnek.
- a populációk genetikai összetételével,
- illetve a genetikai összetételt változtató mechanizmusokkal foglalkozik.
- allélek (A és a), relatív gyakoriságuk p és q és
- genotípusok (AA, Aa, aa) gyakoriságaival D, H, R írhatjuk le.
- az AA genotípusú egyedek relatív gyakorisága D,
- az Aa genotípusúaké H,
- az aa genotípusúaké pedig R
- (D+H+R = 1).
- Intermedier vagy kodomináns öröklésmenet esetén ez nem jelent problémát.
- Domináns-recesszív öröklődésnél viszont a heterozigótákat nem lehet megkülönböztetni a domináns homozigótáktól, ezért a genotípus- és allélgyakoriságokat csak közvetve lehet meghatározni (lásd később).
- az AA homozigóták minden allélja A, (p2)
- de a heterozigótákban csak az allélek fele A. (2pq/2)
- az A allél relatív gyakorisága p = D+H/2; (= p2 + 2pq/2)
- az a allél relatív gyakorisága q = R+H/2. (= q2 + 2pq/2)
- A két allél relatív gyakoriságának összege természetesen 1, vagyis p+q = 1.
- 25 homozigóta domináns AA, D = 25/100 = 0,25
- 50 heterozigóta Aa, H = 50/100 = 0,5
- 25 homozigóta recesszív aa. R = 25/100 = 0,25
- nincs szelekció, azaz az egyedek azonos eséllyel érik meg a felnőttkort,
- nincs mutáció,
- a szaporodás csak ivaros úton történhet,
- a párosodás véletlenszerű, minden egyednek azonosak a szaporodási esélyeik,
- nincs migráció (ki- és bevándorlás) és
- a populáció mérete nagy, így nincs mintavételi hiba, ún. genetikai sodródás.
- a p2 a homozigóta dominánsok,
- a 2pq a heterozigóták,
- a q2 a homozigóta recesszívek relatív gyakoriságát fejezi ki.
- A hímivarsejtek p hányada A, q hányada a genotípusú. Összesen p+q = 1
- A petesejteké szintén: p hányada A, q hányada a genotípusú. Összesen p+q = 1
- A párosodás véletlenszerű. Bármelyik hímivarsejt bármelyik petesejtet megtermékenyíthet.
- (p + q) x (p + q) = 1 azaz (A + a) x (A + a) = 1
- p2 + 2pq + q2 = 1 amiből p = p2 + 2pq/2 és q = q2 + 2pq/2
- p (A) = 0,6
- q (a) = 0,4
- A allélt tartalmazó hímivarsejtek gyakorisága p = 0,6
- a allélt tartalmazó hímivarsejtek gyakorisága q = 0,4
- Összesen: 0,6A + 0,4a = 1
- A petesejteké szintén: 0,6A + 0,4a = 1
- A megtermékenyítés véletlenszerű: (0,6A+0,4a). (0,6A+0,4a) = 1
- A következő nemzedék tehát: 0,36AA + 0,48Aa + 0,16aa = 1
- Amiből A allél gyakorisága: (D+H/2) = 0,36 + 0,48/2 = 0,6 p = p2 + 2pq/2
- csak az AA és Aa genotípus együttes gyakoriságát (D+H),
- illetve az aa recesszív homozigóták R gyakoriságát lehet megadni.
- p2 + 2pq + q2 = 1
- Az Rh-vércsoport a népesség 15%-a, így dd relatív gyakorisága 0,15.
- q2 = 0,15 ebből q = 0,3872
- p = 1- q = 0,6128, ebből p2 = 0,3755 azaz 37,5%
- a heterozigóták 2p.q = 0,475 azaz 47,5%
- Van mutáció.
- Az egyedek ki- és bevándorolhatnak.
- Van szelekció; az egyedek rátermettsége, alkalmazkodó készsége eltérő.
- Nem azonos az egyes genotípusok szaporodási esélye (nem véletlenszerű párválasztás, szexuális szelekció).
- A populáció mérete lehet kicsi.
- A hallópálya 5 neuronos, tehát 4 szinapszis van benne: 4 · 0,5 = 2 ms késés 1 pont
- A 10 cm-t (0,1 m)-t 1 ms alatt teszi meg, a teljes időigény tehát 1+2 = 3 ms 1 pont
- 1,8 / 100 = 0,018 s = 18 ms alatt teszi meg az utat az akciós potenciálhullám. 1 pont
- Eközben 2 szinapszis található, tehát 2,6 ms-ot késik az ingerületátadás, azaz összesen 20,6 ms után jelenik meg a vázizomrost membránján az akciós potenciál. 1 pont
- p+q= 1→p=0,993 1 pont
- Ideális populációban a heterozigóták gyakorisága: 2pq = 2ꞏ0,007ꞏ0,993 = 0,0139 (~ 0,014) 1 pont
- 1700000 ꞏ 0,014= 23800 heterozigóta van Budapesten 1 pont
- Fiú: 0,5 / 50% 1 pont
- Leány: 0 / 0% 1 pont
- q = 1/3500 = 0,00029 1 pont
- A HbS allél gyakorisága: q = (0,1466 + 0,1818) / 2 = 0,1642
- A heterozigóták gyakorisága: 2 pq = 0,2744767 (kerekítve: 0,2745)
- l = 4,25cm → λ = 0,085 m f = c/ λ → f = 4000 Hz (1/s) 1 pont
- Az ábra alapján ez 8 mm-re kelt ingerületet. 1 pont
- Két heterozigótának 0,25 valószínűséggel születik beteg gyereke. 1 pont
- Egészséges ember heterozigótaságának valószínűsége: 2pq/(2pq+p2) 1 pont
- Két egészséges házasságából születendő beteg gyerek valószínűsége ebben a populációban: (2pq/2pq+p2)2 * 0,25 = 0,0359 = 3,59%, kerekítve 3,6%. 1 pont
- F: aa 1 pont
- E: Aa 1 pont
- E-től 0,5 eséllyel kap a gyermeke A allélt, tehát annak a valószínűsége, hogy mindkét gyerek beteg: (0,5)2 = 0,25 1 pont
- 1/2500 · 0,8 · 9800000 = 3136, tehát elvileg 3136 ilyen személy él az országban. 1 pont
- Mindkét lábát azonos mértékben terheli.
- Mind egykarú, mind kétkarú emelők esetében, egyensúlyban a forgatónyomatékok egyenlők.
- A forgatónyomaték az erő és az annak hatásvonalára vonatkozó erőkar szorzata.
- Sziszüphosz súlyát az m∙g képlet alapján számíthatja ki.
- A nehézségi gyorsulás értéke: 9,81 m/s2
- A növekedési (anagén) fázis férfiaknál 2–4, a nőknél 4–6 évig tart. A hajhagyma sejtjei ebben az időszakban alulról folyamatosan kapják a táplálékot, és gyorsan osztódnak, a hajszál naponta kb. 0,35 millimétert nő. A hajszálaknak mintegy 85%-a van ebben a fázisban.
- Ezt követően az átmeneti (katagén) fázis következik, mely 1-2 hétig tart, mialatt a hajszál leválik a hajhagymáról, megszűnik a tápanyag-ellátása. A levált hajszál lassan kifelé tolódik. Ebben a szakaszban szőrtüszőink 3%-a van.
- A szőrtüsző ezután nagyjából három hónapos nyugalmi (telogén) szakaszba ér, ezalatt szünetelteti működését. A hajszál ezt az időszakot lassan kitolódva a szőrtüszőben tölti, közben hajhagymája elszarusodik. A nyugalmi fázis végére e tok elvékonyodik, s már nem akadályozza a távozást. A régi hajszál akkor hullik ki, ha az új kitolja a felszínre, vagy valamilyen mechanikai inger – pl. fésülés – húzza ki helyéről. A szőrtüszőknek egyszerre 10–15%-a van ebben a szakaszban.
- Végül a hajhagyma újra aktivizálódik, az újabb anagén szakaszban vadonatúj hajszál kezd képződni. Egy hajhagyma átlagosan 20 hajszál termelését követően merül ki.
- Egy szőrtüsző teljes ciklusa nőknél: 5 év ("4-6 év") anagén fázis, 10,5 nap ("1-2 hét")
- katagén fázis, és 3 hónap telogén fázis,
- ez összesen 5*12*30+10,5+90=1900,5 nap
- Átlagosan ennyi idő alatt hullik ki a 100000 szál haj,
- vagyis 1 nap alatt 53 szál hullik ki.
- Júlia: Xa XA Péter: XA Y 1 pont
- Tehát a fiúgyermekük 0,5 / 50% eséllyel lesz színtévesztő. 1 pont
- q2=0,005 q= 0,0707, p= 0,9293 1 pont
- Júlia 2pq= 0,1314 valószínűséggel heterozigóta. 1 pont
- Tehát 0,1314 · 0,5 = 0,0657 vagyis 6,6% az esélye, hogy a fiúgyermek színtévesztő lesz. 1 pont
- q2=0,005 q= 0,0707, p= 0,9293 1 pont
- Ha Júlia fenotípusát ismertnek tételezzük fel (színlátó), akkor annak valószínűsége, hogy heterozigóta: 2pq/(p2+2pq), 1 pont
- ezért színtévesztő gyermekük (0,1321) ∙ 0,5 = 0,6603, azaz 6,6% eséllyel születhet. 1 pont
- A forgatónyomaték az erő és az erőkar szorzata.
- Az egy irányba ható forgatónyomatékok összegzendők.
- Az ellentétes irányba ható forgatónyomatékok egyensúly esetén kiegyenlítik egymást.
- A nehézségi gyorsulás értéke: 9,81 m/s2
- A súlyzó és az alkar tömegéből fakadó forgatónyomaték: 14×3×9,81 + 30×15×9,81 = 4826,52 N×cm. 1 pont
- Ez megegyezik a hajlítóizom által kifejtendő forgatónyomatékkal, azaz az alkar által kifejtett erő: 4826,52/4 = 1206,63 N. 1 pont
- 2pq/(p2+2pq) = 0,0750/(0,923+0,075) = 0,075 1 pont
- 0,075·0,075·0,25 = 0,0014 (0,14%) 1 pont
- Számítás és következtetés: p (bükk) = 13/25 = 0,52; q (cifra kankalin)= 10/25 = 0,4 pq = 0,208.
- Lehetséges kölcsönhatás: a zárt bükkös árnyékolása miatt a kankalin túl kevés fényhez jutna. 1 pont
- Eltérő környezeti igények: a kankalin vízigénye jelentősen, de zavarástűrése és nitrogénigénye is kisebb a bükknél, és inkább a meszes talajt (alapkőzetet) kedveli.
- A kövi fodorka gyakorisága: 6/25 = 0,24, pq = 0,096,
- Lehetséges magyarázat: hasonló környezeti igények (W,R,N). 1 pont
- Mindkét szülőnek mindkét jellegre nézve heterozigótának kellett lennie, 1 pont
- ezért az együttes valószínűség: 0,75 • 0,75 = 0,5625 (A 0,56 és az 56,25% és a 9/16 is elfogadható) 1 pont
- Recesszív homozigóta nagyszülő, és heterozigóta szülők esetén. 1 pont
- (5∙106 db/mm3) ∙ (5∙106 mm3) = 2,5 ∙ 1013 (2∙1013 és 3,5∙1013 között elfogadható) 2 pont
- A1 és A2; A1 és A3 / A1, A2, A3 1 pont
- Önmegporzás nem történhet, mert a pollenszemek mindenképpen tartalmazzák az egyik, a szülői egyedben is megtalálható allélt. Másként is megfogalmazható. (Csak indoklással.) 1 pont
- A1A3 : A2A3 : A1A2 = 1 : 2 : 1 1 pont
- A homozigóták aránya 1%, azaz gyakoriságuk 0,01 q2= 0,01, q= 0,1 1 pont
- Ezért p = 0,9 A hordozók gyakorisága pedig 2pq = 2 (0,1 · 0,9) = 0,18 azaz a népesség 18%-a hordozza a kérdéses allélt. 1 pont
- 850 nukleotidpár: 1700 db nukleotid -> 1700 db bázis. 1 pont
- 1700*0,22 = 374 db citozin, s ugyanennyi guanin 1 pont
- 1700-2*374 = 952 , ennyi timin és adenin együtt 1 pont
- ennek fele-fele az adenin s a timin: 476-476 db. 1 pont
- q2 = 1/15 000 q = 1/15000 1 pont
- p=1-q p=1-0,0081 1 pont
- 2pq= 0,016 1 pont
- 1,6% hordozza a betegség allélját 1 pont
- A vizelettel ürülő PAH mennyisége: 1500 · 672 mg = 1 008 000 mg/nap 1 pont
- Ez a vesén áthaladó PAH 90%-a, ezért a teljes áthaladó mennyiség 1 008 000 : 0,9 = 1 120 000 mg/nap 1 pont
- Azaz 1 120 000 : 24 : 60 = 777,8 mg/perc 1 pont
- Ez a mennyiség 777,8 : 1,2 = 648,2 cm3 vérplazmában található meg 1 pont
- Ennyi vérplazmát 648,2 : 0,55 = 1178,5 cm3 vér tartalmaz, azaz A vese vérátáramlása 1178,5 cm3/perc 1 pont
- 1 perc alatt 1500 / 24⋅60 = 1,04 ≈ 1 cm3 vizelet képződik, 1 pont
- A percenként képződő 1 cm3 vizelet (az adat szerint) 672 mg PAH-at tartalmaz 1 pont
- A vérplazmából 90% PAH kerül a vizeletbe, tehát a percenként átáramló vér PAH-tartalma = 672 / 0,9 = 746,7 mg 1 pont
- A PAH-koncentrációból számolható a percenként átáramló vérplazma térfogata: 746,7 / 1,2 ≈ 622,2 cm3 vérplazma/perc 1 pont
- Ennyi vérplazmát 622,2 cm3 / 0,55 =1131,3 cm3 vér tartalmaz, ennyi a vese percenkénti véráramlása 1 pont
- 5,0–6,0 liter közötti teljes vértérfogatot feltételezve 1 pont
- a vérplazma teljes térfogata ennek 55%-a / 2750 cm3 – 3300 cm3. 1 pont
- Ekkor a beadott PAH összes mennyisége 2750 cm3 – 3300 cm3 * 1,2 mg/cm3 = 3300 mg – 3960 mg. 1 pont
- A vizeletben, a feladat szövege szerint 1 nap alatt megjelenő PAH mennyisége 1500 cm3 * 672 mg/cm3 = 1 008 000 mg 1 pont
- E számítás szerint a vizeletben megjelenő PAH mennyiség nagyobb, mint a beadott PAH mennyiség 1 pont
- Egy összehúzódási ciklus 60/75 = 0,8 s-ot vesz igénybe. Ebből 0,22 + 0,08 = 0,3 s az összehúzódás fázisa. 1 pont
- A szív tehát (kb.) 0,8-0,3 = 0,5 s-ot pihen egy ciklus alatt, azaz 1 perc alatt 37,5 másodpercet. 1 pont
 |
Csontváz
Madarak lábtípusai az életmódnak megfelelően alakultak ki:
Izomzat
A madarak szárnyát hatalmas méretű mellizmok mozgatják, amelyek a szegycsont tarajáról erednek.
Kültakaró
A külső hámréteg gyengén fejlett, csak igen vékony szaruréteg borítja, mivel a tollazat átvállalja a szaruréteg fő feladatait;
A kültakaró kizárólag a madarakra jellemző képletei a pikkelyeredetű tollak, amelyek tehát
A tollak lehetnek:
Kontúrtollak melyek lehetnek
Pehelytollak
Dísztollak
A kontúrtollak részei:
Mivel a toll élettelen szaruképződmény, regenerálódni nem tud. Az elhasznált tollak cseréje a vedlés. Erre általában évente egyszer kerül sor. A vedlés hormonális szabályozás alatt áll.
 |
Emésztőrendszer
A fej tömegének jelentős csökkenését eredményezte, hogy a törzsfejlődés során a madaraknál a fogazat helyett csőr alakult ki, melynek üreges csontos alapját kívülről szarutok fedi. A csőr alakja változatos, jól tükrözi az egyes fajok táplálkozásmódját.
Az előbél speciális része a nyelőcső alsó tágulata a begy, amely
A gyomor 2 üregű:
A zúzógyomorból a felaprított gyomortartalom újra visszakerül a mirigyes gyomorba, ahol tovább emésztődik a táplálék.
A középbél az emésztés és a felszívás fő helyszíne.
Az utóbél a húgyvezetőkkel, az ondóvezetőkkel. ill. a petevezetékkel együtt a kloákába torkollik.
Légzés
Az állatvilág legfejlettebb légzőapparátusával rendelkeznek. A repülés jelentős energiaigényének kielégítésére gyors és hatékony anyagcsere kell, ehhez pedig nagy mennyiségű oxigén szükséges.
A madarak tüdeje léghajszálcsöves szerkezetű, aminek köszönhetően - mind a belégzés, mind a kilégzés során - a levegő benne folyamatos áramlásban van.
A tüdőkhöz 5 pár légzsák csatlakozik, melyek térfogatváltozásuk révén biztosítják a tüdőben a levegő folyamatos haladását. A légzsákok kitöltik a madarak testüregét és beterjednek a csontokba is (levegő tartalmú csontok). A légzsákoknak köszönhetően a madarak jóval nagyobb mennyiségű levegőt tudnak a tüdőn keresztül áramoltatni, mint amilyen a tüdőknek megfelelő légtérfogat. Gázcserében nem vesznek részt, az kizárólag a tüdő légzőfelszínén keresztül zajlik (külső gázcsere).
A levegő útja:
orrnyílás → garat → felső gége → légcső → alsó gégefő → főhörgők, melyek belépnek a tüdőbe → faágszerű hörgőcsoportok → tüdősípok, amik hatszögű csőszerű képződmények, ezek falában, a léghajszálcsövekben történik meg a gázcsere → légzsákok → vissza.
Kettős légzésük van, melynek során mind a belégzéskor, mind a kilégzéskor történik gázcsere, mivel a belégzés végén a tüdőből a levegő a légzsákokba kerül, kilégzésnél innen visszaáramlik a tüdőbe, annak tüdősípjait ismét átjárja, így a levegő kétszer halad át a légzőfelületen. Ezen kívül egyes légzsákokba a belégzés során közvetlenül a légcsőből, a tüdő kikerülésével jut az oxigénben gazdag levegő, ami majd innen kilégzéskor kerül a tüdőbe.
A madaraknál a hangadás szerve az ún. alsó gégefő, amely a légcső két főhörgőre ágazásánál alakul ki. Több típusa van, a legfejlettebb az énekesmadarakra jellemző.
 |
Keringési rendszer
A kisebb testű madarak szíve percenként még nyugalmi helyzetben is több százat ver, de egy házi veréb szíve stressz állapotban akár 850-szer is összehúzódhat percenként!!!
Szaporodás
A tojás a madarak
A sárgája szikanyag, azaz jelentős táplálék- és energiaforrás az embrió számára, nagyrészt zsírok és fehérje építi fel. Vitaminokban gazdag, a C vitamin kivételével a legtöbb vitamint tartalmazza, pl. A-, D-, B1-, B2-, E-vitamint.
A tojássárgáját körülvevő fehérje három fontos feladatot lát el:
A fehérjét körülvevő többrétegű burokrendszer – magzatburkok - mechanikai, kémiai, immunológiai védelmet nyújt az embrió számára, ugyanakkor porózussága révén biztosítja a gázcserét.
A petefészekből kilökődött érett petesejt – a szikanyagokban gazdag sárgája - a petevezetőbe kerül, ahol fokozatosan haladva rárakódnak a tojás további alkotórészei: a tojásfehérje, a héjhártyák és a tojáshéj.
Érzékszervek
A repülő életmódnak köszönhetően rendkívül fejlettek az érzékszerveik és az idegrendszerük.
A szemek többnyire a fej két oldalán találhatók, aminek köszönhetően a látótér 300 foknál is nagyobb lehet (az erdei szalonkánál 360 fok), ugyanakkor kisebb a két szem látóterének átfedése, ezért a térlátásuk csekélyebb mértékű (nem úgy a baglyoknál).
 |
21. tétel: Gyűrűsférgek, csigák, rovarok |
 |
22. tétel: Emlősök és hüllők |
 |
23. tétel: Az élőlények viselkedése |
 |
24. tétel: Az állati szövetek |
 |
26. tétel: Ízületek, csontok |
A csontrendszer sokrétű biológiai szerepe
A csontrendszer szerepe a szervezet kalciumanyagcsere-szabályozásában lásd még 4.8.4.
A kalcium-anyagcsere szabályozása – mások mellett, pl. ösztrogének, - nagyrészt három hormon, a parathormon, a D-vitamin (kalcitriol), és a kalcitonin hatásán alapul.
A csontszövet lásd még 3.4.4. fejezet
Az állati szervezet legkeményebb szövete, a csontos halaktól kezdve minden gerinces állat belső vázának az alapanyaga. A csontszövetet
összetétel szerint
felépítés szerint
A szerves állományt
A szervetlen állományt
A test Ca-tartalmának 90%-a a csontokban van, így a csontrendszer egyfajta kalciumraktárként is szolgál.
A szerves állomány a csontok rugalmasságát, a szervetlen állomány a csontok szilárdságát biztosítja.
A csontszövet lemezes szerkezetű, a nyúlványos csontsejtek egymással összekapcsolódva koncentrikus körökbe rendeződnek. A sejtek között az alapállomány szintén körkörösen rendeződött lemezei futnak, amelyet szervetlen sók, kollagénrostok és szerves kötőanyag épít fel. Ily módon a csontszövetben mikroszkopikus hengerek alakulnak ki (ún. oszteonok), melyek közepén idegek és egy vérér fut. A központi vérereket, amelyek a csont hossztengelyével párhuzamosan futnak, sugárirányban haladó erek kötik össze. A csontszövet ilyen módon vérerekkel átszőtt, élénk anyagcseréjű szövet.
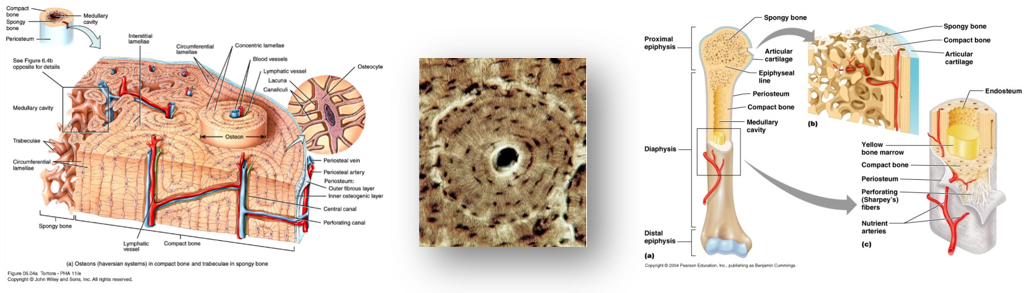 |
A csöves csontok szerkezete
Kívülről minden csontot csonthártya borít, mely
A csonthártya belső rétegében csontképző sejtek helyezkednek el, innen indul a csont vastagodása, ill. a sérült csont regenerációja. A csontszövet az egész élet folyamán mindig az aktuális terhelésnek megfelelően épül fel, azaz, ha pl. öregedéskor a test súlypontja eltolódik, azt a csont belső szerkezetének megváltozása követi. A folyamat során ún. csontfaló sejtek lebontják a csontszövetet, a csontképző sejtek pedig az új terhelés irányában építik át.
A csonthártya alatt tömör csontállomány található, amely lényegében lemezes csontszövet, körülhatárolja a szivacsos csontállományt, melynek üregeit a vörös csontvelő tölti ki, ami jellemzően
A velőüregben, a csöves csontok középső, üreges szakaszában, sárga csontvelő található, amely inaktív, elzsírosodott vörös csontvelő. A velőüreget a csontbelhártya határolja, mely lebontószerepével biztosítja a velőüreg növekedését a csontok vastagodása során.
A csontok összetételének életkori változása
Az életkor előrehaladtával a csontok
Mivel a csontok rugalmasságát biztosító összetevők tartalma idősebb korban csökken, ezért a csontok törékenysége az öregedéssel fokozódik.
A csontok fejlődése
Magzatban a vázelemeket többnyire még porcszövet építi fel, mely fokozatosan alakul át csontszövetté. A csontfejlődés az ivarérettség elérése után ér véget, kb. 16-21 éves korig tart. Gyermekkorban a csöves csontok végein még növekedési zóna, növekedési porcsáv található, ahol a csont hosszirányú növekedése zajlik. A növekedési porckorong a csont vége felé porcot hoz létre, majd a porcbontó sejtek lebontják a porcot és a helyét csontsejtek foglalják el. Először a csontok középső része csontosodik, majd a csontvégek következnek. Az agykoponyacsontok képződése ugyanakkor embrionális kötőszövet (mesenchyma) átcsontosodásával megy végbe.
Mint már volt róla szó, csontképző sejtek nemcsak a csontban, hanem a csonthártya ún. mély rétegében is vannak, melyeknek köszönhető a csontok rárakódásos vastagodása.
Csontok alakja
A csöves csontok
A lapos csontok
A köbös csontok
A szabálytalan alakú, üreges csontok az arckoponyacsontok.
 |
Csontösszeköttetések
A folytonos összeköttetéseknél a kapcsolódó csontok között anyagfolytonosság van.
 |
A megszakított összeköttetéseket ízületeknek nevezzük. Az ízületekben a kapcsolódó csontok között kisebb távolság van, ami a csontok elmozdulását teszi lehetővé. Az ízületeket szerveknek tekintjük, mivel önmagukban lezárt szerkezeti és működési egységek.
Ízületek felépítése
Az ízületi csontvég
Az ízületi tok termeli az ízületi nedvet, mely súrlódáscsökkentő, a porcfelszínt csúszóssá teszi.
Az ízületi szalagok erős kötőszöveti szalagok. Részben az ízületi tokon kívüli önálló képződmények, részben a tokhoz tartozó képletek. Az ízületek összetartásában - az ízületi tokon és szalagokon kívül – a külső légnyomásnak is jelentős szerepe van.
Ízületek típusai
Az ízületek típusait az elmozdulás mértéke, iránya és a felszínek alakja szerint csoportosítjuk.
A feszes ízületek tényleges mozgásra alkalmatlanok, ill. csak minimális elmozdulást tesznek lehetővé, mert ízületi felszíneik szabálytalanok, szalagkészülékük pedig igen feszes, ilyen pl.
A mozgékony ízületek a kapcsolódó csontok nagyobb mértékű elmozdulását teszik lehetővé. A legtöbb ízületünk ide tartozik, további csoportosításuk az ízületi felszín alakja és az elmozdulás iránya alapján történik.
 |
 |
Koponyacsontok
A koponya két részből áll,
Az agykoponya csontjai az agyat veszik körül, jellemző rájuk, hogy varratos összeköttetésekkel kapcsolódnak össze (idősebb korban összecsontosodnak).
A magzati koponya csontosodása magzati korban nem fejeződik be, születéskor a csontok még nem érnek össze, közöttük kötőszövetes lemezek (kutacs) találhatók, melyeknek köszönhetően egyrészt a magzat könnyebben halad át a szülőcsatornán, másrészt lehetővé teszik csecsemőkorban az agy jelentős térfogatnövekedését.
Az agykoponyán – nyakszirtcsonton - egy nagyobb nyílás, az öreglyuk található, ahol a nyúltvelő folytatása, a gerincvelő kilép a koponyából. Az agykoponya alapján még sok kisebb nyílás is látható, amelyek erek és agyidegek ki- és belépési helyei.
Az agykoponyát 7 csont, 2 páros és 3 páratlan alkotja:
 |
A törzsváz csontjai
Két része van: a gerinc és a mellkas.
Gerinc
A csigolyák elhelyezkedése és alakja alapján a gerincet szakaszokra osztjuk.
Szakaszai:
A csigolyák mérete lefelé, a növekedő terhelésnek megfelelően egyre nő. A keresztcsonti csigolyák kb. 2-3 éves korra egységes keresztcsonttá nőnek össze, ami segíti a medence stabilitását, ezáltal a felegyenesedést.
A csigolyák felépítése
Az első 24 csigolya valódi csigolya, a keresztcsont és a farki csigolyák nagymértékben módosulnak, ezért ezeket álcsigolyáknak nevezzük.
A valódi csigolyákon megkülönböztetjük
A csigolyák között párosan kétoldalt lépnek ki a gerincvelőidegek.
A mellkas
A mellkast 12 pár borda és a szegycsont alkotja. A bordákat a szegycsonthoz való kapcsolódásuk alapján 3 csoportba soroljuk.
Hátul a bordák a hátcsigolyák testéhez és harántnyúlványaihoz ízesülnek.
A szegycsont a kulcscsonttal és 10 pár bordával ízesül, elöl ez zárja a mellkast, lapos csont, vörös csontvelőt tartalmaz.
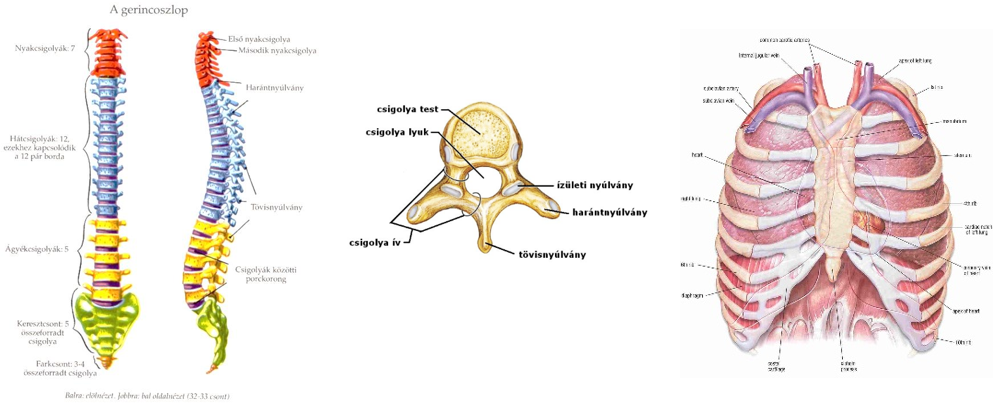 |
Végtagok csontjai
A végtagokon megkülönböztetünk függesztőövet és szabad végtagot.
Felső végtagok
Függesztőöv a vállöv, részei
Szabad felső végtag részei a kar, az alkar és a kéz, csontjai:
Az alsó végtagok
Függesztőöv a medenceöv, mely 3 nagy csontból áll,
A medencecsont a
A medence alakja másodlagos nemi jelleg, nőknél szélesebb és alacsonyabb, a szeméremcsontok által bezárt szög pedig tompább, mint a férfiaknál, aminek a szülésben van jelentősége.
A szabad alsó végtag részei a comb, a lábszár és a láb, csontjai
A test középvonalához közelebb eső sípcsont alsó, kiszélesedő része a belső bokát, a szárkapocscsont hasonló része a külső bokát képezi.
 |
A mozgásszervrendszer egészségtana
Csípőficam
A csípőficam a combcsont fejének a csípőbe való ízesülésének zavara. A csípőficam újszülöttkortól diagnosztizálható fejlődési rendellenesség. Jellemzően a lánycsecsemők gyakrabban érintettek, mint a fiúk. Kialakulásának hátterében részben genetikai tényezők állnak, részben a magzat méhen belüli elhelyezkedése játszik szerepet.
Kezelésénél elsődlegesen ún. terpeszpelenkázást alkalmaznak, melynek eredményeképpen a combcsont feje megfelelően illeszkedik a medencecsonthoz. Amennyiben ez elégtelen, Pavlik-kengyel használata javasolt, súlyos esetben pedig 1 éves kor felett műtéttel lehet korrigálni a rendellenességet.
Ficam
A ficam során az ízületi fej kimozdul az ízületi vápából és kóros helyzetben rögzül, súlyosabb esetben az ízületi tok elrepedhet, az ízületi szalagok elszakadhatnak. Jellemző tünetei a fájdalom, a duzzanat, a mozgáskorlátozottság, az ízület erőteljes torzulása.
Rándulás
A ránduláskor az ízületi fej kimozdul az ízületi vápából, majd visszatér az eredeti helyzetbe. Tünetei hasonlóak a ficamhoz, azonban annál enyhébbek.
Sérüléskor elsősegélynyújtáshoz a sérült ízületet helyezzük nyugalomba, polcoljuk fel, tegyük sínbe, alkalmazzunk hideg borogatást, ficam esetén a további kezeléseket – a kificamodott ízület helyretétele, szükség esetén műtét –- kizárólag orvos végezheti.
Gerincferdülés (scoliosis)
A gerinc oldalirányú elhajlása, amihez egyidejűleg a csigolyák hossztengely körüli elfordulása is társul. Mivel a mellkas deformálódik, szövődményként jelentkezhet fájdalom, a szív és a tüdő működésében zavarok léphetnek fel. A gerincferdülés kezelése enyhébb esetben gyógytornával történik, súlyosabb esetben ajánlott a megfelelő fűző viselése, ill. esetenként szükséges lehet műtéti beavatkozás.
Porckorongsérv
Az egészséges porckorong egy korong alakú szerv, mely a külső részén körkörös, rostos gyűrűből áll, míg belül magas víztartalmú kocsonyás mag található. Porckorongsérv esetén a rostos gyűrű átszakad és a kocsonyás anyag a gerinccsatorna vagy gerincideg felé kiboltosodik. Az életkor előrehaladtával a porckorong elkezd kiszáradni, vizet veszít és így elveszíti rugalmasságát. Az ellaposodó porckorongban egy hirtelen terhelésre a gyűrű átszakad, a belső, kocsonyás anyag kinyomódik és létrejön az előbb említett kitüremkedés. Ez megtörténhet egy rossz mozdulat, nehéz súlyok nem megfelelő emelése során. A porckorongsérv kialakulásának a kockázatát növeli a helytelen testtartás, a túlsúly, az edzetlenség, illetve a bemelegítés nélküli hirtelen erőkifejtés.
Lúdtalp
A láb egészségesen meglévő boltozatának megsüllyedése. Kialakulhat fejlődési zavar következtében, de leggyakrabban túlterhelés, túlsúly, izomgyengeség az oka.
Csonttörések
A csonttörés esetén a csontállomány folytonossága megszakad. Ilyenkor a csontban törési rés keletkezik. A kiváltó ok lehet egy közvetlen erőhatás, de előfordulhat másik betegség következményeként is, mint pl. daganatos betegségeknél, vitaminhiánynál, csontritkulásnál. A csonttörés lehet
A csonttörések tünetei: fájdalom, duzzanat, vérömleny, mozgáskorlátozottság, nyílt törésnél külső-belső vérzés. A csonttörés gyanúját röntgenfelvétel, MRI vagy CT vizsgálat erősítheti meg.
Enyhébb esetben a csonttörés konzervatív módon kezelendő, pl. külső rögzítéssel (gipsz vagy kötés), súlyosabb esetben műtéti beavatkozás szükséges, melynek során velőűrsínnel, csavarral, lemezzel stb. rögzítik a törést.
Implantátumok, csontprotézisek
A csontok sérüléseinek (csonttörés), kopásainak kezelésére műtéti úton beépített különféle eszközök használhatók. A beépített eszközöket implantátumnak, illetve ízületi és csontkopások esetén protézisnek is nevezzük.
Az implantátumok nagy részének az anyaga rozsdamentes acélötvözet vagy titánötvözet, lehetnek lemezek, szegek, csavarok.
A protézisek a kopások esetében a pótlandó csontterületek formáját utánozzák, alkatrészei egyes esetekben készülhetnek műanyagból (polietilén) vagy kerámiából is.
27. tétel: Az emberi izmok |
Mozgásunk megvalósítása kétféle szervrendszerrel történik:
Az emberi testben több, mint 600 elkülönült vázizmot különböztetünk meg, amely a testtömeg átlagosan kb. 45-50%-át adja.
Az izmokat az inak kapcsolják a csontokhoz,
Az inakat rostos kötőszöveti pólya, az ínhüvely veszi körül, amely súrlódást csökkentő folyadékot tartalmaz.
Az izmok középső, izomrostokat tartalmazó, összehúzódó része az izomhas. Az izomfejek a két, vagy több résszel eredő izmok különálló részei, melyek száma szerint beszélünk biceps, triceps, quadriceps izmokról.
Az izmok alak szerint lehetnek
 |
Fontosabb izmaink:
A végtagokon ellentétesen - antagonista módon - működő izmokat különböztetünk meg, mint pl. hajlító és feszítő izmokat. A hajlító izmok összehúzódásakor a kapcsolódó csontok által bezárt szög csökken (hajlítás), a feszítő izmok összehúzódásakor a csontok által bezárt szög nő (feszítés).
Távolításkor az adott testrészek – végtagok – a test középvonalától, ill. egymástól távolodnak – ujjak –, közelítéskor pedig az előbbivel ellentétes irányú mozgást végeznek.
Forgatásnak nevezzük a csont hossztengelye mentén történő elfordulást.
A vázizmokat vázizomszövet építi fel, amely harántcsíkolt izomszövet, megtalálható
A vázizmok működésére jellemző, hogy
 |
A vázizmok felépítése lásd még 3.4.4. fejezet
Az izomroston belül továbbá megkülönböztetünk
A miofibrillumok felépítése és az izom-összehúzódás
A vázizmot fény- vagy elektronmikroszkópban vizsgálva az izomrostok harántirányban – a hossztengelyre merőlegesen – csíkozottságot mutatnak, azaz világosabb és sötétebb sávok váltják egymást. A sávozottságot az okozza, hogy az izomfonalkötegek a hossziránynak megfelelően szakaszosan váltakozva más-más felépítésűek, de az egyforma szakaszok pontosan egymás mellett helyezkednek el. Az izomfonalak már csak elektronmikroszkópban láthatók. Kétféle izomfonal váltakozik hosszirányban:
A vékony fonalakban a globuláris aktin monomerek – G-aktin – összekapcsolódva két egymás köré tekeredő, gyöngysorszerű fonalat képeznek, az F-aktint. A vékony fonalakat a Z-lemeznek nevezett képlet tartja egybe. A két Z-lemez közé eső szakaszt szarkomernek nevezzük. A szarkomera az izomfonalköteg periodikusan ismétlődő egysége.
A vastag fonal a miozin fehérjéből áll, melynek ATP-bontó enzimhatása van. A vastag fonalat 24 miozinmolekula építi fel (mint a kötegbe fogott golfütők).
A miozinmolekula két részből áll:
Az izmok előbbiekben vázolt felépítése – elemi fehérjék (aktin, miozin) + segédfehérjék → izomfonalak → izomfonálköteg → izomrost → izomnyaláb → izom – jól mutatja a hierarchikusan szerveződő élőrendszerek felépítését és működését.
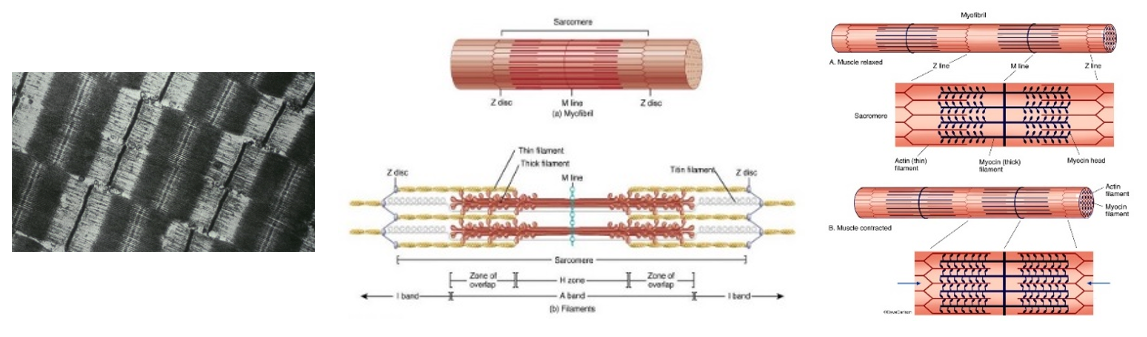 |
Az izom-összehúzódás csúszómechanizmusa
Egy elernyedt, nyugalomban lévő izomban a vékony fonalak csak kis mértékben nyúlnak be a vastag fonalak közé, a vékony és a vastag fonalak közötti kapcsolódások száma csekély. Az izom-összehúzódás során a vékony fonalak becsúsznak a vastag fonalak közé, aminek köszönhetően a Z-vonalak egymáshoz közelítenek, s így a szarkomerek megrövidülnek. Az egyes szarkomerek megrövidülése kismértékű, de mivel az izom-összehúzódás mindig az izom hossztengelyével párhuzamosan zajlik, méretcsökkenésük összeadódik, ami az izom nagymértékű megrövidülését eredményezi.
Lépések röviden
A fenti lépések sokszori ismétlődése miatt a szarkomerek megrövidülnek. Az összehúzódás addig tart, ameddig a Ca-ion jelen van. A miozinfejek mozgása, alakváltozása energiaigényes, melynek fedezetét az ATP-hidrolízis energiája jelenti. Ez az izom-összehúzódás csúszómechanizmusa. A csúszás a Z-vonalakat egymás felé közelíti, a vékony fonalaknak megfelelő világos sáv szélessége csökken, ugyanakkor a sötétebb átfedési sáv szélessége nő, az összehúzódás alkalmával tehát a szarkomerek rövidülnek meg és nem az izomfonalak.
 |
Animáció
https://drive.google.com/file/d/18P_WffEvP1uhB9-6OLkjNYfh6measIi4/view?usp=sharing
A vázizomrostokat az idegrendszer idegrostok útján működteti. Az idegi összeköttetéseitől megfosztott vázizom elsorvad. Az idegrostok ingerülete szinapszisok közvetítésével átterjed az izomrostokra és akciós potenciált vált ki. A Ca2+ hiánya fokozza az ideg-izom ingerlékenységet, csökkenti az izomrostok ingerküszöbét, ami izomgörcsökhöz vezet. A Mg2+ hiánya hasonló következményekkel jár, hiszen az ATP nem önmagában, hanem Mg2+-ionnal együtt kötődik a miozin feji részéhez. A Mg-ATP-komplex kötődés hatására az aktin-miozin kapcsolat bomlik. Ha nincs Mg-ATP, akkor a miozinfej hozzákapcsolódva marad az aktinhoz (rigor állapot).
Az idegrostok szétágazhatnak és több izomrostot láthatnak el. Egy idegrosthoz kapcsolódó izomrostok motoros egységet képeznek. A finom működésű izmokban (kéz izmaiban) egy idegrosthoz átlagosan 3-6 izomrost kapcsolódik, míg a durvább mozgások kivitelezésében közreműködő izmokban – pl. comb feszítő izma – akár több száz.
Az izomerő attól függ, hogy hány motoros egység dolgozik egyszerre. Mindig csak annyi izomrost húzódik össze, amennyi az adott erő kifejtéséhez szükséges, a többi pihen. A dolgozó rostok kimerülésekor a pihenők átveszik a feladatot.
Ha megfelelően rövid időközönként több idegi impulzus éri az izmot, tartós összehúzódás váltható ki. Ilyenkor az izom nem tud elernyedni, az összehúzódások ereje összeadódik.
Az izom működése lehet:
Az izomműködés energiaforrásai
A tápanyagokat – zsírokat, szénhidrátokat (glükózt), aminosavakat – az izomrostok a vérből veszik fel. A pihenő izom a szénhidrátokat jelentős mennyiségben képes tárolni glikogén formájában. Az izom-összehúzódás közvetlen energiaforrása az ATP-hidrolízis energiája. Az ATP egyrészt a biológiai oxidáció, másrészt relatív oxigénhiány esetén a tejsavas erjedés során jön létre. A vázizomszövetben az ATP-n kívül az energiatárolásban egy másik vegyület, a kreatin-foszfát is közreműködik. A kreatin-foszfát feladata, hogy a kimerülő ATP-készleteket regenerálja.
kreatin-P + ADP = kreatin + ATP
Az ábrán az egyes energiaforrások százalékos megoszlásának változása látható egy 5 perces maximális terhelés során.
Ismert a hemoglobinon kívül egy másik oxigénkötő fehérje is az emberi szervezetben, a mioglobin, amely a vázizmokban található. A mioglobin csupán egy polipeptidláncból és egy vastartalmú hemből álló összetett fehérje. A két fehérjében az oxigénkötődés mechanizmusa hasonló, ugyanakkor a mioglobin alacsonyabb pH-n és alacsonyabb oxigénkoncentráció esetén is képes kötni az oxigént, hiszen feladata az oxigén raktározása. Tartós összehúzódás vagy hosszantartó izomműködés esetén, amikor a vázizmok oxigénellátásának hatékonysága csökken, a mioglobinhoz kötött oxigén leválva átmenetileg képes kompenzálni a kialakult oxigénhiányt.
Hosszantartó izomműködés esetén szervezetünk energiakészletei kimerülhetnek, ami a fáradságérzet kialakulásának egyik oka. Az izmok kifáradásának hátterében számos különböző tényező áll, mint pl.
Tartós fizikai terhelés során a szervezet fáradással szembeni ellenálló képességét az állóképességgel jellemezzük. Az edzettség a fizikai terhelés hatására kialakuló egyfajta erőnléti állapot, teljesítőképesség.
A rendszeres, folyamatos sportolás, a fokozódó terhelés növeli az edzettséget, mellyel párhuzamosan javul az állóképesség, a szervezet azon képessége, hogy a nagy fizikai megterhelés során biztosítja a biológiai egyensúly fenntartásához szükséges energiát.
Az összehúzódás sebessége alapján megkülönböztetünk:
 |
Biomechanika
A biomechanika a mechanika törvényszerűségeit alkalmazza az élő szervezetekre. Az emberi test mechanikus működése egyes esetekben hasonlatos az egyszerű emelők működéséhez.
Az emelő egy rögzített (forgás)tengely (forgáspont) körül elfordítható merev rúd, amely használható teher emelésére vagy mozgatására. A tengely vagy forgáspont az emelő elforgathatóan megtámasztott pontja.
Kétoldalú (kétkarú) emelőről beszélünk, ha a teher(erő) (súly) és az aktív erő a tengelyhez képest az emelő különböző oldalán található.
Egyoldalú emelőről beszélünk, ha a teher és az aktív (emelő) erő az emelő ugyanazon oldalán van a tengelyhez képest.
A forgáspont és a teher közötti távolság a teherkar (k1),
a forgáspont és az aktív erő közötti távolság pedig az erőkar (k2).
Az erő forgató hatását megadó fizikai mennyiséget forgatónyomatéknak hívjuk. Egy erő forgatónyomatékát megkapjuk, ha az erő nagyságát megszorozzuk az erőkarral.
Forgatónyomaték = Erő x Erőkar Erő = Tömeg x Gyorsulás F = m x g
Az emelő törvénye kimondja, hogy egyensúly esetén az aktív erőnek a forgáspontra vonatkozó forgatónyomatéka egyenlő a teher forgatónyomatékával:
Emelőelv: Teherkar × Tehererő = Erőkar × Erő azaz G×(k1) = F×(k2)
A szívizomszövet
A szívizomszövet, mely a harántcsíkolt izomszövetnek egy speciális félesége, a szív falának középső rétegében található meg.
Jellemzői:
Alaktanilag két sajátságban különbözik a vázizomszövettől:
 |
A simaizomszövet
A simaizomszövet elnyúlt orsó alakú sejtekből épül fel. A sejtmag a sejt közepén helyezkedik el. A sejtekben megtalálható az aktin és a miozin, de a harántcsíkolt izomszövetre jellemző nagyfokú rendezettség nem jellemző.
Előfordulása:
Tulajdonságai:
28. tétel: A fehérjék emésztése |
Az emésztés során a tápanyagok makromolekulái a tápcsatorna üregében monomerekre hasadnak, hidrolizálnak, melyek aztán felszívódással többnyire a vérbe kerülnek.
A bontást
A fehérjék emésztése a gyomorban kezdődik meg. A gyomor nyálkahártyájának kötőszöveti rétege csöves mirigyeket tartalmaz. A mirigyek
Ezen váladékok összessége a gyomornedv.
A magas H+-koncentráció következtében a gyomornedv pH-ja O,9 és 1,5 között van, amely optimális feltételeket biztosít a pepszin számára.
A pepszin fehérjeemésztő enzim, amely inaktív formában, mint pepszinogén kerül elválasztásra. A pepszinogén részben az alacsony pH hatására, részben autokatalízissel aktiválódik. A pepszin a fehérjéket adott aminosavaknál hasítja, így darabolva oligopeptidekre bontja őket.
A hasnyálmirigy külső elválasztású része termeli a hasnyálat, amely emberben napi 1,2-1,5 l, pH-ja 8 körül van. A hasnyál tartalmaz tripszint, amely inaktív tripszinogén formájában választódik el, a polipeptideket oligopeptidekre bontja (peptidkötéseket hidrolizál, endopeptidáz).
A vékonybélben a tápanyagok makromolekuláinak monomerekre való végső bontását a bélhámsejtek felszínéhez kötött enzimek végzik. A membránpeptidázok - az aminopeptidáz és a dipeptidáz - oligopeptidek hidrolízisét végzik, így befejezik a fehérjék aminosavakká történő hidrolízisét.
Az aminosavak többsége aktív transzporttal szívódik fel a vékonybélből a vérbe.
29. tétel: A szénhidrátok és lipidek emésztése |
Az emésztés során a tápanyagok makromolekulái a tápcsatorna üregében monomerekre hasadnak, hidrolizálnak, melyek aztán felszívódással többnyire a vérbe kerülnek.
A szénhidrátok emésztése a szájüregben megkezdődik. A nyálat a szájüregbe nyíló 3 pár nagy nyálmirigy termeli:
Naponta kb. 5OO-6OO ml nyál termelődik, kémhatása semleges, ill. enyhén lúgos. Fő összetevői:
A hasnyálmirigy külső elválasztású része termeli a hasnyálat, amely emberben napi 1,2-1,5 l, pH-ja 8 körül van. A hasnyál tartalmaz amilázt, amely a nyálamilázhoz hasonló, ezért szintén a keményítőt és a glikogént hasítja maltózegységekre.
A vékonybélben a tápanyagok makromolekuláinak monomerekre való végső bontását a bélhámsejtek felszínéhez kötött enzimek végzik. A maltáz, a szacharáz, a laktáz diszacharidok monoszacharidokra bontásáért felelősek.
A monoszacharidok aktív transzporttal szívódnak fel a vékonybélből a vérbe.
A ragadozók és a mindenevők vastagbele a tápanyagok felszívásában nem játszik fontos szerepet. Itt főleg víz, ionok, és esetleg kis tömegű szerves anyagok, mint pl. B- és K-vitaminok szívódnak fel.
A növényevők vastagbelében levő jelentős mikrobatömeg egyszerű szénhidrátokat, aminosavakat, zsírsavakat és B-vitaminokat képez, melyek itt még felszívódnak.
A lipidek – neutrális zsírok - emésztése a vékonybélben zajlik a hasnyálmirigyben termelődő lipáz segítségével.
A hasnyálmirigy külső elválasztású része termeli a hasnyálat, amely emberben napi 1,2-1,5 l, pH-ja 8 körül van. A hasnyál tartalmaz tehát lipázt – kezdetben inaktív, epesavak hatására válik aktívvá –, amely a lipidek észterkötéseit hidrolizálja, így a neutrális zsírokat monogliceridekre és zsírsavakra bontja.
A lipidek emésztését az epe segíti. A lúgos kémhatású epe a májban termelődik, a vékonybél egyik igen fontos emésztőnedve, azonban emésztőenzimeket nem tartalmaz. A képződött epe az epehólyagban raktározódik, s közben jelentősen bekoncentrálódik. Az epevezeték a hasnyálmirigy vezetékével egyesülve közösen nyílik a patkóbélbe.
Az epében mások mellett epesavak (epesavas sók, kialakítják a lúgos kémhatást) találhatók.
Az amfipatikus epesavak feladata kettős:
A nagyobb hidrofób molekulák – pl. a zsírok, mivel vízben oldhatatlanok, lipoproteinek formájában – a nyirokkeringésbe jutnak.
30. tétel: Az emésztés szabályozása |
A táplálkozás során a táplálékkal vesszük fel a szervezetünk felépítéséhez és az életfolyamatok fenntartásához szükséges tápanyagokat. Szűkebb értelemben táplálkozás alatt csak a táplálék bevitelét értjük a szervezetbe, de tágabb értelmezésben ide soroljuk a táplálék anyagainak megemésztését, felszívódását és az anyagcserében történő hasznosulását, illetve az emészthetetlen anyagok kiürítését is.
A táplálék tartalmazza a létfontosságú tápanyagokat, ill. a szervezet számára fel nem használható, felesleges anyagokat (salakanyagok).
Élelmiszer minden olyan anyag, amely változatlanul vagy feldolgozva fogyasztásra alkalmas.
A tápanyagok funkció szerint csoportokba sorolhatók:
Továbbá fontos tápanyag a víz, mely szerkezetmeghatározó, oldószer, reakciópartner és reakcióközeg, szállítóközeg.
Minőségi éhezés kialakulása során elegendő mennyiségű táplálék kerül a szervezetbe, de abban nem megfelelő az egyes tápanyagok mennyisége, ill. aránya, leggyakrabban ásványi anyagok, vitaminok, esszenciális aminosavak hiánya okoz problémát.
Mennyiségi éhezés során az elfogyasztott tápláléknak a kelleténél kevesebb a mennyisége és az energiatartalma, ezért a bevitt táplálék nem fedezi a napi energiaszükségletet.
Az éhség egy motivációs állapot, a táplálék felkeresésére, a táplálék elfogyasztására irányuló belső késztetés. Kiváltásában többféle élettani tényező – pl. alacsony vércukorszint, gyomor üressége stb. – játszik szerepet.
A táplálkozással kapcsolatos idegi szabályozó mechanizmusok központjai a hipotalamuszban vannak. Itt található
Az éhség érzetét
Ez a szöveti hormon továbbá
Szintje a vérben étkezés előtt a legmagasabb és közvetlenül utána a legalacsonyabb. Testsúlycsökkenéskor nő a vérben a ghrelinszint, hízáskor pedig csökken.
A jóllakottság jelzései
A leptin a ghrelinhez hasonlóan a hipotalamikus neuronokon – jóllakottsági központ – keresztül fejti ki a hatását. A tartósan alacsony leptinszint folyamatos éhségérzetet okozhat, ezért a leptinhiány már kora gyermekkortól jelentkező súlyos elhízást eredményez. Ennek ellenére sok esetben az elhízás magas leptinszint mellett is kialakul. Ezt a jelenséget leptinrezisztenciának nevezzük.
Egy adott élelmiszer vércukorszintre gyakorolt hatását a glikémiás indexszel lehet jellemezni, mely egy 0-100-ig terjedő számérték, nagyságát
A magas indexű tápanyagok gyorsan és nagymértékben emelik meg a vércukorszintet, aminek következtében az inzulinelválasztás jelentősen fokozódik, s mivel az inzulin csökkenti a vércukorszintet, a táplálkozást követően 1-2 órával hipoglikémia alakul ki, ami erős éhségérzetet eredményez.
Az agytörzsben találhatók a táplálkozással kapcsolatos nyálkahártyareflexek központjai, mint pl. a nyelés, rágás, hányás, nyálelválasztás.
31. tétel: Az ember légzése |
Bevezetés, alapfogalmak
A légzőszervrendszer azon túl, hogy
számos feladatot lát el, mint pl.
A lebontó anyagcserében keletkező szén-dioxid felhalmozódása veszélyeztetné a szervezetünk homeosztázisát, tekintve, hogy vízben oldódva szénsavvá alakul, ami a testfolyadékok kémhatását savas irányba tolja el. Ebben az értelemben a légzőszervrendszer kiválasztó működést is végez.
A terminális oxidációhoz szükséges oxigént, ill. az elsősorban a citrát ciklusban keletkező felesleges szén-dioxidot légzési gázoknak nevezzük.
A légzést fizikai és kémiai folyamatként is értelmezzük. Fizikai értelemben beszélhetünk
A diffúzión alapuló gázcsere lehet
A légcsere a tüdő és a külső légtér között zajló, nyomáskülönbségen alapuló levegőáramlás.
Kémiai értelemben légzésnek a biológiai oxidáció folyamatát értjük, ekkor a folyamatot sejtlégzésnek is nevezzük.
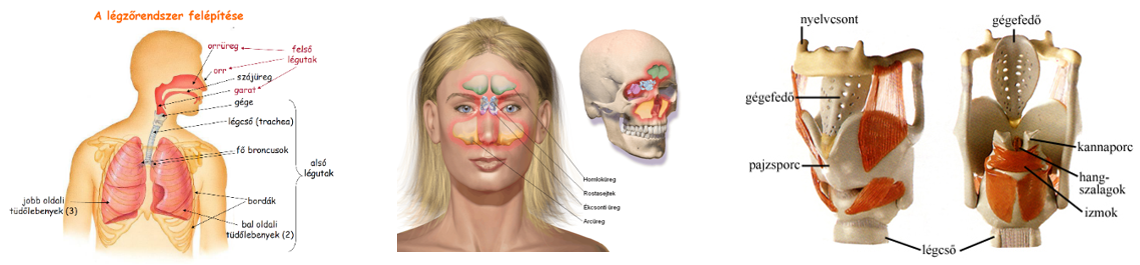 |
A légzőrendszer felépítése
A légzőszervrendszerhez tartoznak
továbbá tágabb értelemben
A levegő a tüdő légzőfelszínéig a légutakon keresztül jut el. A felső légutakhoz tartozik
az alsó légutakat
Az alsó légutak a főhörgőktől kezdve a tüdőben futnak.
Nyugodt légzés esetén a levegő az orron, ill. az orrüregen keresztül jut a légutakba. Megkülönböztetjük az orr külső részét és belső üregrendszerét.
Az orr külső részét
Az orrüreg igen bonyolult járatrendszer, részben a külső orrban, részben a koponya üregrendszerében található. Az orr üregrendszere
A nagyobb üregek – melyeket középen a csontos, ill. porcos orrsövény választ el – az külső orrnyílásokkal kezdődnek és a belső orrnyílásokkal nyílnak a garatba. Az orrüreg hátsó részének külső, oldalsó falain helyezkedik el a 3 pár csontos orrkagyló – alsó, középső, felső – amelyek növelik az orrüreg belső felületét és kialakítják a megfelelő légáramlatokat.
A melléküregek az orrüreg kiöblösödéseként fejlődnek ki. Ilyen
Az orr üregrendszerét – a melléküregeket is – nyálkahártya béleli, felületét csillós hengerhám fedi, amelyben igen sok nyálkatermelő mirigysejt van. A csillók csapkodási iránya olyan, hogy a nyálkába beletapadt port a garat felé hajtja, ahonnan a szennyeződések a nyeléssel a gyomorba kerülnek. A hám alatti kötőszövet igen gazdag mirigyekben, erekben. Az orrüreg felső részén található az idegi eredetű, kemoreceptorokban gazdag szaglóhám.
Nyugodt légzés esetén a levegő az orrüregen keresztül jut az alsó légutakba, így feladata
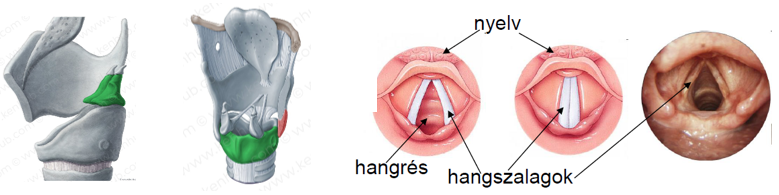 |
Hangképzés
Az alsó légutak a gégével nyílnak a garatból. A gége
A gége porcai
A porcok közül a legnagyobb a pajzsporc, mely elölről határolja a gégét, két egymással elöl összetalálkozó lemezből áll. A két lemez által bezárt szög az életkorral változik, ill. másodlagos nemi jelleg, férfiaknál a pajzsporc lemezei hegyesszöget zárnak be, nőknél és gyerekeknél a bezárt szög tompább. Minél kisebb a lemezek által bezárt szög, annál hosszabbak a hangszalagok és ennek megfelelően annál mélyebb a hang fekvése. A mély hangú férfiak pajzsporcának csúcsi része erősen előre ugró, ez az ádámcsutka.
A pecsétgyűrűhöz hasonlító gyűrűporc a pajzsporc alatt található. A hátrafelé néző “pecsét” felső széléhez ízesülnek a páros, háromoldalú, piramis alakú kannaporcok.
A kannaporcok alapjánál erednek a hangszalagok.
A gégefedő porca felülről határolja a gége nyílását, levélszerű, rugalmas porcból álló lemez. Helyzeténél fogva megakadályozza, hogy nyeléskor a falat a légcsőbe jusson.
A gége porcait erős kötőszövetes lemezek tartják egybe.
A gége porcai közül a kannaporcok mozgékony ízületekkel kapcsolódnak a gyűrűporchoz, aminek köszönhetően a porcok között kifeszülő nyálkahártyalemezek és ezek széli részei, a hangszalagok a gége űrterének alakját változtatni képesek. A porcokat több harántcsíkolt izomköteg mozgatja.
A hangszalagok - melyek hámszövetből, kötőszövetből és vázizomszövetből épülnek fel – a kannaporcok és a pajzsporc között feszülnek ki, a köztük levő háromszögletű nyílás a hangrés, amely normális légzés alatt nyitott, háromszög alakú. A két hangszalag összezáródásával a gége egyrészt légmentesen lezárható, másrészt a kiáramló levegő általi rezgésük hangok képzésére teszi őket alkalmassá.
Hangképzéskor a hangszalagok közelednek egymáshoz, a hangrés szűkül, a kiáramló levegő megrezegteti a hangszalagokat, aminek következtében a felettük levő légoszlopban rezgéshullámok – hanghullámok – keletkeznek.
A hang magassága a hangszalagok vastagságától, hosszától, feszességétől és a hangrés alakjától függ. A szűkebb hangrés és a kifeszült, ill. rövidebb hangszalagok magas hangot eredményeznek. A hangmagasság érzetét a rezgésszám/frekvencia határozza meg.
A hangerő a kiáramló levegő mennyiségétől és sebességétől függ. Lásd még 4.8.1.10. fejezet.
Az egyes magán- és mássalhangzók képzésében a gégén kívül a garat, a nyelv, az ajkak, a fogak, a száj-, és az orrüreg is részt vesznek.
Az egyes emberek hangszíne eltérő, aminek az az oka, hogy a személyekre jellemző hangképzéskor a gégében keletkezett alaphangok az egyénenként különböző felületekről – fogakról, nyelvről, szájpadlásról –, és a rezonátorüregekből visszaverődve különböző erősségű felhangokkal dúsulnak.
A felhangok (más szóval részhangok) egy zenei hanggal egy időben megszólaló, de külön nem hallható hangok sora. A hangzó test anyaga, alakja és mérete mellett a részhangok mennyisége és ereje határozza meg a hangszínt: azonos magasságú hangok ezért szólnak másképpen a különböző hangszereken és énekhangokon.
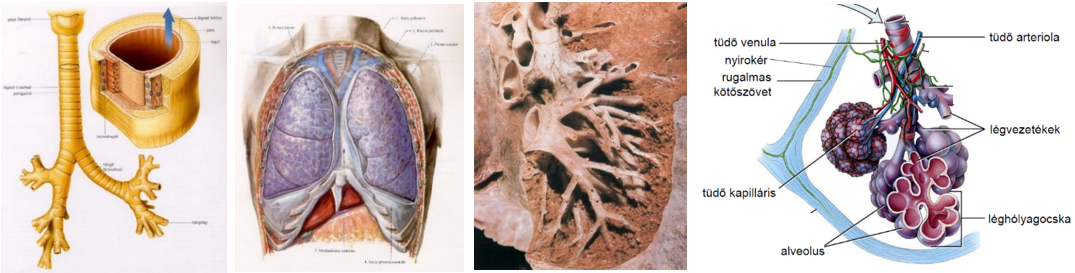 |
Az alsó légutak további szakaszai
A légcső C alakú porcokból (20) felépülő, nyálkahártyával bélelt cső. Mögötte, hozzá szorosan kapcsolódva helyezkedik el a nyelőcső. A légcső nyálkahártyáját csillós hengerhám borítja, a hámban sok nyálkatermelő kehelysejttel. A csillók csapkodása a garat felé áramoltatja a nyálkarétegbe tapadt port.
A légcső két főhörgőre oszlik, melyeket porcgyűrűk merevítenek, de egyébként a légcsővel azonos felépítésűek.
A két főhörgővel a légutak a tüdőben folytatódnak. A főhörgők egyre kisebb átmérőjű hörgőkre és hörgőcskékre ágaznak, amik végül a léghólyagokban végződnek. Az elágazódások bonyolult rendszere az ún. tüdőfát hozza létre.
A főhörgőktől a hörgőcskék felé haladva folyamatosan változik a légutak falának szerkezete. A fal felépítésében egyre kevesebb a porcszövet – a hörgőcskék falából már teljesen hiányoznak a porcok –, egyre több a kötőszövet és a simaizomszövet, a hámszövet is egyre laposodik, a léghólyagok falában már laphám található.
A tüdő a mellüregben található páros szerv, oldalról a mellkasfal, alulról a rekeszizom határolja, felső csúcsa a kulcscsontig nyúlik. Alakja kúphoz, állománya szivacshoz hasonlítható. A két tüdő egymás felé eső felszínein találhatók a tüdőkapuk, ahol a tüdők erei és a főhörgők lépnek be, ill. ki. A tüdőt mély hasadékok lebenyekre tagolják, a jobb 3, a bal 2 lebenyből áll.
A tüdőben a legutolsó hörgőcskeelágazások a szőlőfürtszerű léghólyagokba nyílnak, amelyeket léghólyagocskák bonyolult rendszere épít fel. A léghólyagocskák felülete a légzőfelület, emberben elérheti a 150 m2-t, itt zajlik a külső gázcsere.
Ennek megfelelően faluk igen vékony. A léghólyagocskák felszínén többféle sejttípust különböztetünk meg, találhatók itt
A léghólyagocskák közötti igen vékony válaszfalban vannak
Az egyes légzési gázok diffúziójának az irányát a koncentrációkülönbségek, az ezzel arányos parciális nyomáskülönbségek határozzák meg.
A parciális nyomásérték az a részleges nyomásmennyiség, ami a gázkeverék összes nyomásából az illető komponensre jut. Pl. a levegő össznyomása 101 kPa, ebből 21 kPa jut az oxigénre, és kb. 0,04 kPa jut a szén-dioxidra.
A tüdőben a levegő összetétele különbözik a külső levegő összetételétől, a gázcsere miatt kevesebb oxigént és több szén-dioxidot tartalmaz.
A tüdő a mellkasban saját térfogatánál nagyobb teret tölt ki, a mellhártyák által a tüdő mintegy ki van feszítve a mellkas belső felszínére. A mellhártya a nagy savós hártyák – hashártya, szívburok, mellhártya – egyike. A két tüdőnek külön mellhártyarendszere van, melyek zsákszerűen veszik körül a tüdőfeleket. A mellhártyának két lemeze van, a külső lemez a mellkas belső felszínére, a belső lemez a tüdő külső felszínére tapad. A két lemez között súrlódást csökkentő savós folyadék található, aminek a nyomása mindig kisebb, mint a tüdőben a levegő nyomása, így a tüdő kifeszül a mellkas falára.
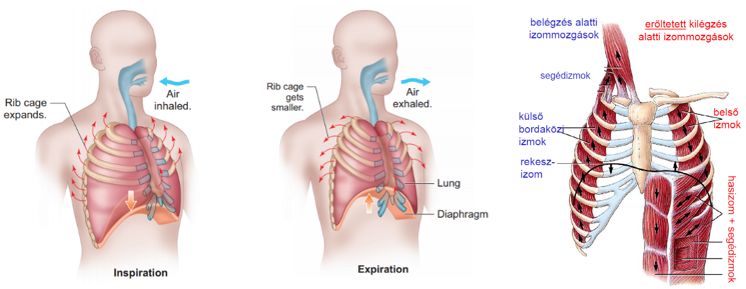 |
Légzőmozgások
A külső légtér és a tüdő légtere közötti légcserét a mellkas térfogatát változtató légzőmozgások teszik lehetővé. A tüdőnek saját izomzata nincs – leszámítva az egyes légutakat szűkítő simaizmokat –, így önálló mozgásra képtelen, a mellhártyák közvetítésével belülről rátapad a mellkasfalra, passzívan követi annak mozgását, térfogatváltozásait. Ez a térfogatváltozás a vázizmok közé sorolható, légzőizmoknak nevezett
Nyugodt belégzéskor
Az aktív izomösszehúzódásoknak köszönhetően
Attól függően, hogy a belégzésben mely izmok működése dominál, megkülönböztetünk hasi légzést és mellkasi légzést.
A hasi légzésben a rekeszizom vesz főleg részt, ez inkább a tüdő alsóbb részeit tölti meg levegővel, a férfiak, a gyerekek, ill. a sportolók körében gyakoribb.
A mellkasi légzésnél inkább a bordaközi izmok működése hangsúlyosabb, így elsősorban a tüdő felső része telik meg levegővel, ez inkább a nőkre jellemző.
Erőltetett belégzéskor a vállizmok is szerepet játszanak a váll felemelése révén a mellkas térfogat növekedésében.
Nyugodt kilégzés során
A nyugodt kilégzés passzív folyamat, a belégzőizmok elernyedése miatt történik, a mellkas súlyánál fogva nehezedik rá a tüdőre, s összenyomja. Ezt segíti a tüdő szövetének nagyfokú rugalmassága, ami a tüdőt összehúzni igyekszik.
Erőltetett kilégzéskor már ún. járulékos kilégzőizmok is közreműködnek, mint pl.
A légzőműködések szabályozásáért felelős idegi központok az agytörzsben vannak. A légzőmozgások szabályozásában a legfontosabb inger a vér szén-dioxid-tartalma, koncentrációjának növekedése a belégzés kiváltásáért felelős neuroncsoport aktivitását fokozza.
A légcsere térfogati jellemzői
A légcsere térfogati változásait spirométerrel lehet meghatározni.
Fizikai terhelés során – az intenzív izomműködés energiaszükségletének fedezéséhez – fokozódik a biológiai oxidáció, mely eredményeképp nő a vér CO2- koncentrációja, ami a légzőműködések intenzitásának – a légzési perctérfogatnak – növekedését eredményezi.
Ugyanakkor minél intenzívebb fizikai terhelésnek van kitéve egy szervezet, annál több oxigénre van szüksége a működő izmok energiaellátása érdekében. Az oxigénfelvevő képesség azonban korlátozott és egyénileg változó. A test által maximálisan felvehető és szállítható oxigén mennyiségét a V(O2)max értékkel adhatjuk meg. A V(O2)max megadja pl., hogy a test 1 kilogrammja hány milliliter oxigént szállít és vesz fel percenként (ml/kg/min).
A V(O2)max értéke kor-, nem- és edzettségfüggő. Az életkor előrehaladtával a légzőrendszer rugalmasságának elvesztése, a mellkasfal merevségének növekedése, a légzőizmok erejének csökkenése, a légzőfelület és a tüdőt ellátó kapillárisok csökkenése korlátozza a V(O2)max értékét.
A megnövekedett oxigénigényt
Tehát pl. futás közben egy sportoló percenkénti légzésszáma kevesebb, mint egy nem sportolóé, miközben légzési perctérfogatuk közel azonos. A fizikai igénybevétel hosszú távon a tüdő levegőbefogadó képességének növekedésével jár, aminek egyenes következménye a tüdő vitálkapacitásának növekedése. Ennek értéke átlagos felnőtt szervezet esetében 4,5 dm3 körüli, sportolóknál és fúvós hangszeren játszó zenészeknél elérheti akár a 6-7 dm3-t is.
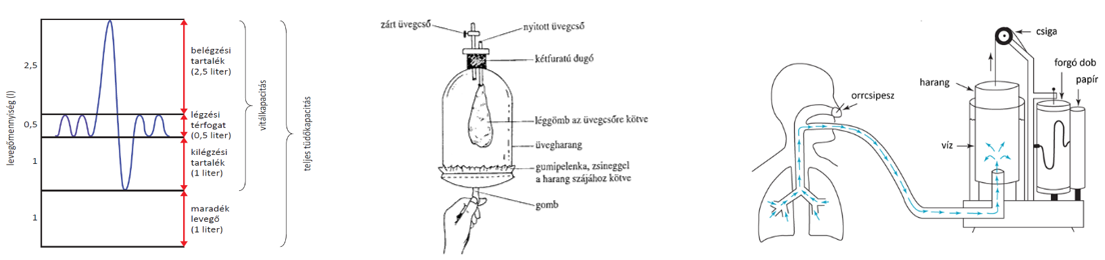 |
A légzőmozgásokkal kapcsolatos nyomás- és térfogatváltozások
Belégzés során a tüdőben eleinte csökken a nyomás, majd a folyamat második felétől ismét növekedik, amíg el nem éri újra a légköri nyomás – 760 Hgmm, 101 kPa – értékét.
Kilégzéskor eleinte növekedik a nyomás a tüdő légterében, majd a folyamat második felétől elkezd csökkenni addig, ameddig egyenlő nem lesz ismét a légköri nyomással.
A mellhártya két lemeze között a nyomás végig kisebb, mint a tüdőben, ami biztosítja, hogy a tüdő rátapadjon a mellkas belső felszínére. A különbség a tüdőben mérhető nyomásváltozáshoz képest, hogy a belégzés során mindvégig csökken, a kilégzés során pedig mindvégig nő a nyomás a két lemez között. Amennyiben a mellhártya két lemeze közé levegő kerül, a tüdőben található rugalmas rostoknak köszönhetően a tüdő kis gombóccá ugrik össze, s így képtelen követni a mellkas mozgását. Ez a jelenség a légmell.
A légzőrendszer nyomás- és térfogatváltozásainak összefüggéseit a Donders-féle tüdőmodellel demonstrálhatjuk.
A kísérletet
A rekeszt modellező gumimembránt lehúzva a harang és a tüdő közötti térben a légköri nyomásnál alacsonyabb nyomást hozunk létre, a változást a léggömb rugalmas fala passzívan követi, kitágul és levegő áramlik a belsejébe (belégzés).
A gumilapot elengedve, az visszatérve eredeti helyzetébe, fokozza a nyomást a harang belsejében, a léggömb összenyomódik és a levegő kiáramlik (kilégzés).
A légzés és a légzőrendszer egészségtana
A légzőszervrendszeri megbetegedéseket okozhatják
Az egyik leggyakoribb, viszonylag enyhe lefolyású rendellenesség az orrnyálkahártya-gyulladás, mely különböző eredetű gyulladások összefoglaló neve. Típusai
A szénanátha egyike a legelterjedtebb allergiáknak. A szénanáthát leggyakrabban
A közönséges nátha az orr nyálkahártyájának
A garatgyulladás vagy torokgyulladás rendszerint
A középfülgyulladás rendszerint a felső légutak gyulladásos folyamataival hozható kapcsolatba, tekintve, hogy a garat a fülkürtön keresztül kapcsolatban áll a középfüllel, s így az ide bejutó kórokozók idézik elő a gyulladást.
Rekedtség esetén
A hörghurut
Az asztma
A tuberkolózis (tbc)
A tüdőgyulladás elsősorban
A dohányzás hazánkban az alkoholfogyasztás után a második legveszélyesebb legális droghasználat, Magyarországon az áldozatok száma évente közel 28 ezer. A cigarettában 50 vegyületről ismert, hogy rákkeltő hatású. A dohányban lévő nikotin nagyobb mennyiségben halálos méreg, kis mennyiségben pedig élénkítő hatású,
A nikotin azért nem okozhat mérgezést, mert lebomlása nagyon gyors, halálos dózisban nem képes felhalmozódni a szervezetben. A nikotin a központi idegrendszerben az ún. jutalmazó központokban dopamin felszabadulását váltja ki. A dopamin felelős a nikotin-függőség kialakulásáért.
A dohányzás
A tüdőrák a hörgőkben vagy a tüdő szövetében kialakuló rosszindulatú daganat. Kialakulását eredményezhetik a dohányzáshoz köthető kátrányszármazékok, ill. okozhatja légszennyeződés, azbeszt, egyéb vegyi- és radioaktív anyagok.
A légutakba került idegen test – pl. étel, folyadék – légúti elzáródást okozhat, ami az oxigén-utánpótlás megszűnése miatt életveszélyes állapothoz vezethet. A mentők kiérkezéséig minél hamarabb kezdjük meg az elsősegélynyújtást. Próbáljuk eltávolítani az idegen testet, köhögtessük a sérültet, ütögessük a lapockák közötti területet, ill. gyakoroljunk hirtelen nyomást a bordák alatti hasi részre. Ha a sérült eszméletlen, fektessük stabil oldalfekvésbe.
Gázmérgezésről
32. tétel: A vérgázok szállítása |
A vörösvértestek
A vörösvértestek emlősökben sejtmag nélküli, önálló mozgásra képtelen, középen homorú, korong (bikonkáv) alakú sejtek.
Számuk férfiakban 5 millió/mm3, nőkben 4,5 millió/mm3.
A vörösvértestek szárazanyagának 95%-át a hemoglobin tölti ki. A hemoglobin 4 alegységből felépülő, vörös színű, összetett fehérje.
Minden alegység
A vörösvértestekben sejtmag, belső membránrendszer, mitokondrium nincs, s így szinte teljes tömegét a hemoglobin adja. Anyagcseréje minimális, mivel mitokondriumokat nem tartalmaz, a sejt csupán glikolízist, ill. tejsavas erjedést folytat.
A vörösvértestek feladata az oxigén és részben a szén-dioxid szállítása. Az oxigén a hem vas-ionjához kötődik, miközben a fehérje harmadlagos és negyedleges szerkezete egyaránt megváltozik.
A hemoglobin oxigénnel való telítettségének a mértékét elsősorban a közeg oxigén koncentrációja - s ezzel arányosan a parciális nyomása - határozza meg.
A hemoglobin-molekula oxigénkötő-képességét az ún. oxigéntelítési görbével jellemezhetjük (szaturáció).
A haemoglobin (folyamatos vonal) és a mioglobin (szaggatott vonal) szaturációja (oxigén telítettsége) a pO2 függvényében. A felső vastag vonal a tengerszinten a tüdőben mérhető pO2 mellett észlelt szaturációt jelzi.
A hemoglobin fontos tulajdonsága, hogy oxigéntelítettsége pH-függő. Minél savasabb a vér, úgy csökken az oxigénkötődés erőssége, azaz alacsonyabb pH-n a hemoglobin molekulák kevésbé kötik az oxigént. Ez a jelenség biztosítja, hogy a működő szövetekben - ahol a keletkezett szén-dioxid savas kémhatást okoz - a hemoglobin könnyebben adja le az oxigént.
A tüdőben fordított a helyzet, ahol a magas oxigén koncentráció - és magasabb pH miatt - a hemoglobin újra oxigénnel telítődik.
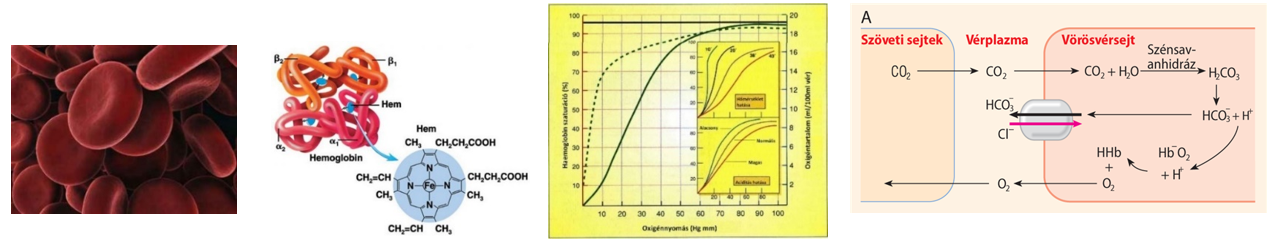 |
A szövetekben:
A tüdőben
A CO2 molekulák továbbá
A magzat hemoglobin molekulái abban különböznek a felnőttétől, hogy bennük a béta alegység helyett gamma alegység van. A magzati hemoglobin erősebben köti az oxigént, mint a felnőtt hemoglobin, aminek köszönhetően a magzat vére át tudja venni az oxigént az anyai vérből.
Ismert a hemoglobinon kívül egy másik oxigénkötő fehérje is az emberi szervezetben, a mioglobin, amely a vázizmokban található. A mioglobin csupán egy polipeptidláncból és egy hemből álló fehérje.
A grafikon a hemoglobin és a mioglobin oxigéntelítettségét mutatja, a vér oxigénje (parciális) nyomásának függvényében. Az A vér a vénás vér, a B vér az artériás vér. A két fehérjében az oxigén kötődés mechanizmusa hasonló, a grafikonról ugyanakkor megállapítható a mioglobin alacsonyabb pH-n és alacsonyabb oxigénkoncentráció esetén is köti az oxigént, hiszen feladata az oxigén raktározása.
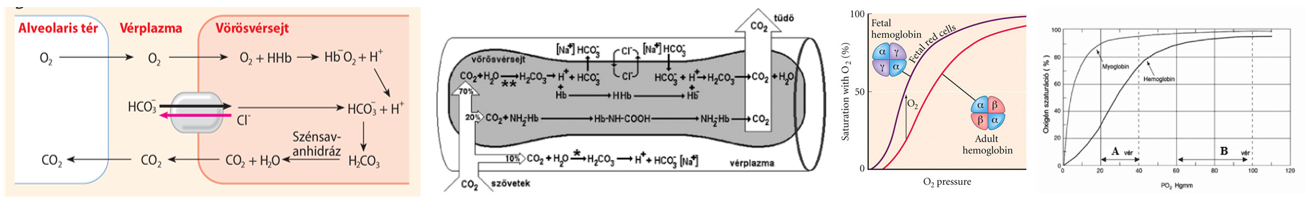 |
33. tétel: A szénhidrátok útja a szervezetben |
A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek.
A szénhidrátok biológiai jelentősége az emberi szervezetben:
A táplálékkal felvett szénhidrátok emésztése a szájüregben megkezdődik amiláz enzim hatására. A nyálamiláz feladata a keményítő, glikogén maltózzá bontása.
A hasnyálmirigy külső elválasztású része termeli a hasnyálat, amely szintén tartalmaz amilázt, amely a nyálamilázhoz hasonló, ezért szintén a keményítőt és a glikogént hasítja maltózegységekre.
A vékonybélben a poliszacharidok monomerekre való végső bontását a bélhámsejtek felszínéhez kötött enzimek végzik. A maltáz, a szacharáz, a laktáz diszacharidok monoszacharidokra bontásáért felelősek.
A monoszacharidok Glükóz-Na+ kotranszporttal szívódnak fel a vékonybélből a vérbe. A glükóz felvétele aktív transzport, az alacsonyabb koncentrációjú hely felől a magasabb koncentrációjú hely felé szállítódik, amihez az energiát a Na+ koncentrációgradiense által meghatározott diffúziója biztosítja. Tehát a glükóz energetikailag kedvezőtlen irányba megy, a Na+ pedig kedvezőbe, de végső soron a kettő eredője kedvező. A mechanizmus másodlagos aktív transzport, hiszen a folyamat feltétele a Na+ koncentrációkülönbsége a sejthártya két oldalán, melyet a Na+-K+ pumpa hoz létre ATP felhasználásával.
A monoszacharidok a vérkeringés útján eljutnak a sejtekhez, melyek facilitált diffúzióval, szállítómolekulák segítségével felveszik őket. Többféle – 14 - glükóztranszporter ismert (GLUT).
Inzulin hatására a GLUT-1 transzporterek mellé, az addig a sejt belsejében, vezikulákban tárolt GLUT-4 transzporterek beleolvadnak a külső membránba, s megnövelik a zsírsejtek és az izomrostok glükózfelvételét.
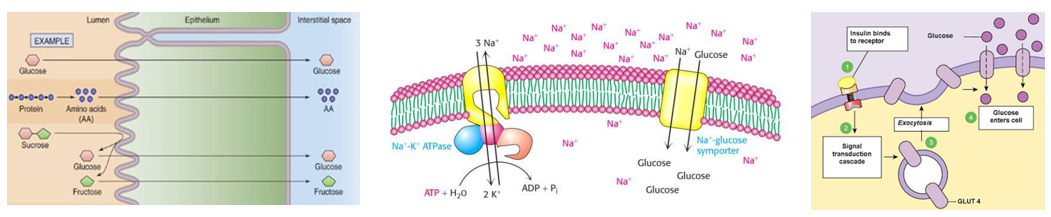 |
A sejtekbe jutott monoszacharidok - többnyire glükóz -
A máj a szervezet szénhidrát forgalmának legfontosabb szabályozója:
A máj szénhidrátforgalommal összefüggő, egyik igen fontos feladata a vércukorszint szabályozása.
A máj szénhidrátforgalmát egyrészt idegi, másrészt hormonális tényezők befolyásolják.
A szervezetszintű szénhidrátforgalmat szabályozó fontosabb hormonok
Az inzulin
Az inzulin egy 51 aminosavból álló polipeptid. Az inzulin az anyagcserét anabolikus irányba tolja el, a tápanyagok, testépítő vegyületek beépülését, raktározását serkenti.
Összefoglalva:
Az inzulin elősegíti a glükóz felvételét, lebontását, zsírokká (máj, zsírszövetek) vagy glikogénné (máj, vázizom) alakulását.
Egészséges emberekben a vizelet gyakorlatilag cukormentes, azonban inzulin hiányában, mivel magas a vércukorszint a glükóz megjelenik a vizeletben. Ennek következtében
Bármely, a csatornában nem felszívódott ozmotikusan aktív anyag – pl. cukor – csökkenti a víz visszaszívódását, s emiatt a következő szakaszba érkező folyadék mennyisége megnő. A későbbi szakaszok nem képesek a megnövekedett vízmennyiséget visszaszívni, ezért a vizelet mennyisége megnövekszik. Ezt a jelenséget ozmotikus diuresisnek nevezzük, ezzel magyarázzuk a cukorbetegek nagyobb mértékű vizeletürítését.
Az adrenalin
Az adrenalin kémiailag aminosav származék, hatását tekintve ún. stresszhormon, nyugalomban a vérben szintje minimális, azonban megterhelés esetén a szervezet izgalmi állapotának a kialakításáért felelős.
Hatása:
A kéregállomány középső rétegében elsősorban a szénhidrát anyagcserét befolyásoló hormonok termelődnek (glükokortikoidok), melyek közül a legfontosabb a kortizol.
A kortizol az éhezéshez történő alkalmazkodásért felel:
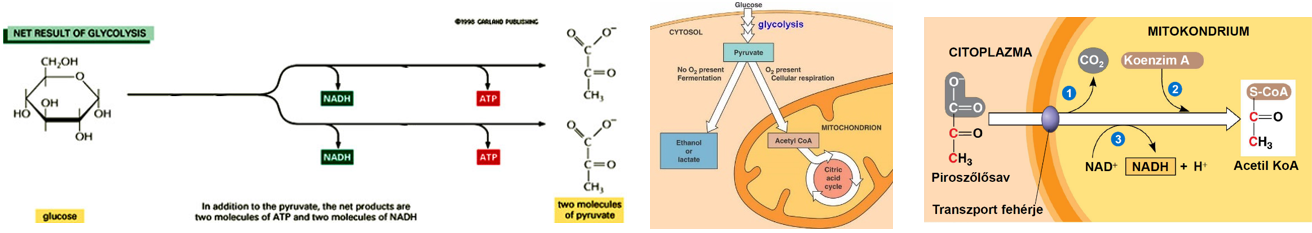 |
A szénhidrátok sejteken belüli lebomlása
Központi jelentőségű anyagcsere-folyamat:
A glükózlebomlás 2 útja:
A biológiai oxidáció
Ha a glükóz lebomlása oxigén jelenlétében folyik, aerob körülmények között, biológiai oxidációról beszélünk. A glükóz lebomlásának leghatékonyabb módja, melynek terméke a szén-dioxid és a víz.
C6H12O6 + 6 O2 = 6 CO2 + 6 H2O
Három fő szakaszra osztható:
A glikolízis
Glikolízisnek nevezzük a glükóztól a piroszőlősavig vezető reakciósorozatot.
A glikolízis eredményeképpen a glükózból molekulánként keletkezik:
A glikolízis nettó energianyeresége 2 ATP glükózmolekulánként (szubsztrát szintű foszforiláció).
A piroszőlősavnál (piruvát) a reakcióutak szétágaznak.
Aerob körülmények között a piroszőlősav CO2 és H vesztése mellet acetilcsoporttá alakul.
Az acetilcsoport szállítómolekulához, KoA-hoz kapcsolódik, amely elszállítja a citrátkörbe.
Piroszőlősav + KoA = acetil-KoA + CO2 + NADH + H+ (oxidatív dekarboxileződés)
A piroszőlősav aerob bomlása és a további reakciók eukariótákban már a mitokondriumban, prokariótákban a citoplazmában játszódnak le.
A citrátkör, citromsavciklus, Szent-Györgyi-Krebs-ciklus
A citrátkör lényege, hogy az acetilcsoport C-atomjai szén-dioxiddá alakulnak. A keletkezett szén-dioxid a légzés útján távozik szervezetből.
Az oxidáció során felszabaduló hidrogének H-szállító koenzimekhez (NAD+, FAD) kapcsolódnak.
A ciklus során több lépésben távoznak a szén-dioxid- molekulák, ill. a H-atomok.
A folyamat energianyeresége minimális, mindössze egyetlen GTP keletkezik acetilcsoportonként, melynek bomlása ATP képződését teszi lehetővé.
A citrátkör az anyagcsere egyik legfontosabb anyagelosztója. A folyamat köztestermékei különböző bioszintézisek kiindulási vegyületei.
A glükóz hidrogénjei H-szállító koenzimekhez kapcsolódtak, NADH-kat képezve, amelyek nagy energiatartalmú vegyületek, mivel erősen redukáltak. Energiatartalmuk felszabadítása végső oxidációjukkal történik, amely a terminális oxidáció folyamatában valósul meg.
A terminális oxidáció
A terminális oxidáció lényege, hogy a NADH-ról származó hidrogének elektronjai - a fotoszintézis fényszakaszához hasonlóan - egy elektrontranszportláncba kerülnek, ahol az elektronok áramlása energiafelszabadulással jár, amely ATP szintézisére fordítódik (oxidatív foszforiláció). Az elektronok szállításában vastartalmú összetett fehérjék, ún. citokrómok vesznek részt.
Az elektrontranszportlánc utolsó tagja oxigént köt meg, így az elektronok végső soron az oxigénre kerülnek. Az így keletkezett oxidion az oldatban található protonokkal vízzé egyesül.
Tehát a NADH szubsztrátoktól átvett protonjai és elektronjai – hidrogénjei - egy bonyolult elektron-transzportláncon keresztül jutnak el a végső elektronfelfogó molekulához, az oxigénhez.
A teljes folyamatsort, a glükóztól a szén-dioxid és a víz keletkezéséig, sejtlégzésnek, biológiai oxidációnak nevezzük.
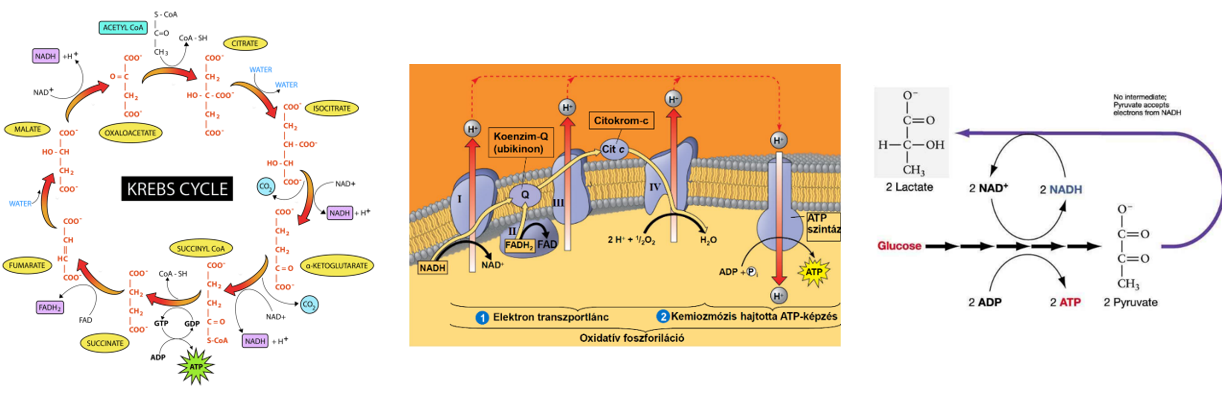 |
A tejsavas erjedés
Anaerob körülmények között a glikolízisben keletkezett NADH-molekulák oxigén hiányában nem tudják a H-jüket a terminális oxidációban leadni, emiatt mennyiségük megnő a citoplazmában.
A felhalmozódó NADH-val ugyanakkor a glikolízisben keletkezett piroszőlősav pedig tejsavvá alakul. A folyamatnak köszönhetően a NAD+-molekulák újra képződnek, aminek köszönhetően a glikolízis zavartalanul végbemehet.
Tejsavas erjedés zajlik az állati szervezetekben és az emberben a vörösvértestekben, a vázizomszövetben romló oxigénellátottság esetén. A tartós izomösszehúzódás csökkenti az izmok oxigénellátottságát, az oxigénhiány miatt az izmok működésükhöz az energiát tejsavas erjedéssel biztosítják.
Idővel a vérkeringés a májba szállítja a tejsavat, amelyből újra glükóz képződik kortizol hormon hatására (glükoneogenezis), ill. oxidálódva lebomlik.
34. tétel: Véralvadás |
A vérlemezkék
A vér legkisebb alakos elemei, számuk 300 000/mm3. Nevük arra utal, hogy emlősökben szabálytalan alakú, sejtmag nélküli, apró sejttöredékek, így élettartalmuk 5-10 nap, igen rövid.
A vörös csontvelőben keletkeznek úgy, hogy az őssejtekből differenciálódó óriássejtek sejtplazmájából hasadnak le. A bennük található enzimek révén véralvadásban játszanak szerepet.
 |
A vérzéscsillapítás, a véralvadás
A keringési rendszer alapfeladata a szállítás, a különböző anyagok eljuttatása a test egyik pontjáról a másikra. Az elmozdulás igénye szükségessé teszi, hogy ennek funkcionális közege folyékony legyen. A vér és a nyirok zárt rendszerben áramlik, azonban előfordulhat, hogy a csőrendszer fala kisebb-nagyobb sérüléseket szenved, így a benne lévő folyadék eltávozhat. Ennek megakadályozására a rendszerben keringő folyadék kis részlete képes megszilárdulni, ami tömítést képezve elzárja a nyílást. E meglehetősen komplikált folyamatot vérzéscsillapításnak nevezzük.
A vérzéscsillapítás szakaszai:
A sérült ér artériás részén szerotonin hatására az érfal reflexesen összehúzódik, csökkentve az átfolyó vér mennyiségét és nyomását.
Az érfalat bélelő hámréteg sérülésekor a vérrel érintkező kötőszöveti kollagén hatására
Amennyiben a fenti folyamatok nem elegendők a vér elfolyásának a megakadályozására, megindul a véralvadás bonyolult, többlépcsős biokémiai láncreakciója, mely lényegében egy kaszkádrendszer, melyben a résztvevő anyagok láncreakciószerűen egymást aktiválják.
Összesen 13-féle összetevője van a véralvadásnak, amelyeket I-XIII római számokkal jelzett véralvadási faktoroknak nevezzünk. Ilyenek pl.
A véralvadási faktorok nagy részét a máj termeli.
A véralvadási folyamat két – belső, külső - párhuzamos, egymást erősítő úton indul el, ahol a kezdő lépések különböznek, a további történések viszont azonosak.
A külső út (extrinsic)
A kezdő lépést az váltja ki, hogy a sérülés helyén a vér érintkezésbe kerül az érfal külső sejtjeinek a membránjában elhelyezkedő szöveti faktornak (régen tromboplasztin) nevezett integráns membránfehérjével. A szöveti faktor nem található meg a vérrel közvetlenül érintkező sejtek (érbelhártya sejtek, fehérvérsejtek, vörösvérsejtek) membránjában. Ezért a vérben található faktor csak az érsérülés helyén találkozik a szöveti faktorral, ez a találkozás indítja meg véralvadás láncreakcióját.
A VII. faktor aktiválódásának eredményeképpen, közvetetten, más faktrorok – Xa. és az V. - aktiválódásának köszönhetően, Ca2+ jelenlétében a vérplazmában megtalálható inaktív protrombin aktív trombinná alakul.
A protrombin a májban termelődik, előállításához K-vitamin szükséges, ezért a K-vitamin elengedhetetlen a normális véralvadáshoz, hiánya vérzékenységhez vezet.
A vérzés csillapodásával a vérlepényen át, a nyomásviszonyok miatt, egy sárgás áttetsző folyadék szűrődik ki. Ez a vérszérum (vérsavó), amely fibrinogén mentes vérplazma.
A véralvadást a sebgyógyulás követi. Olyan sejtek hálózzák be az alvadékot, melyek kötőszövetet építenek a seb helyén. A sérült sejtek elhalnak, alkotóelemeik lebomlanak. A vérrög lebontásáért főként a plazmin nevű enzim a felelős. A fibrin megjelenése aktiválja a plazmint, amely több darabra hasítja a fibrint.
Természetes körülmények között a véralvadás az extrinsic út faktoraival indul meg. Az extrinsic út elegendő trombint biztosít a fibrinogén → fibrin átalakuláshoz.
A belső utat az érfal sérülésének következtében megjelenő kollagénrostok indítják meg. Az intrinsic út folyamatsora felerősíti az alvadási folyamatot, stabilizálja az alvadékot és korlátozza az idő előtti fibrinolízist.
Továbbá az intrinsic út teszi lehetővé a levett vér megalvadását kémcsőben, ahol a folyamat kezdő lépése a vérplazma érintkezése az üvegfelülettel.
Laboratóriumi körülmények között, a levett vérben, a Ca2+ eltávolításával, megkötésével megakadályozható a véralvadás (kalciumkötő anyagok: EDTA, citrát, oxalát).
Műtétek során a nemkívánatos alvadás megakadályozására, az egyes faktorok aktiválódását gátolják meg pl. a máj által termelt heparinnal, ugyanis fiziológiás körülmények között a kalcium-ion megkötése nem járható út, mivel a vérplazma Ca2+ koncentrációjának a csökkenése súlyos izomgörcsöket eredményez, ami fulladásos halált okozhat.
Az alvadási rendszer állapotáról alvadási (trombin) idő mérése tájékoztat. Ez azt az időtartamot jelenti, amely alatt az üvegedénybe frissen levett vérben megjelenik az első fibrinszál. Az alvadási idő átlagos értéke 5-6 perc.
A véralvadási folyamat egy adott részéről nyújt információt a protrombin idő. Másodpercben megadja, hogy a szöveti faktor hozzáadását követően mennyi idő telik el a vizsgált vérmintában az alvadék képződéséig. A vizsgálatra például akkor van szükség, ha a trombózisra hajlamos páciens valamilyen szájon át szedhető véralvadásgátló kezelést kap, segítségével vizsgálható, hogy a véralvadásgátló gyógyszerek megfelelőek-e a vérrög képződésének megelőzésére.
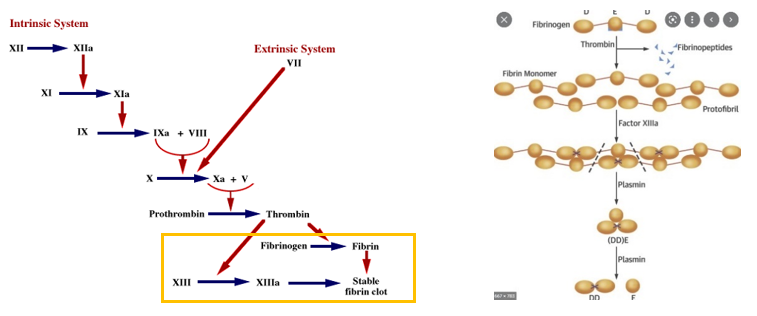 |
A vérzékenység
A vérzékenység fokozott vérzést jelent, leggyakoribb oka a véralvadás zavara. Tág fogalom, a kissé elhúzódó egyszerű vérzésektől az életveszélyes állapotokig terjedhet.
Kóros vérzékenység – szerzett, ill. veleszületett - létrejöhet:
Ér-eredetű rendellenességek
A kiserekből kiinduló spontán vérzések jellemzik. Az ilyen jellegű vérzések a bőrön és nyálkahártyán keletkeznek, aprón, pontszerűen.
Leggyakoribb kiváltó tényezők:
Öregedés: az érfalak rugalmatlanok, sérülékenyek, melynek eredményeként gyakran látjuk az idősek bőrén a szétfutó, élénkvörös bevérzéseket.
Hiányállapotok: legismertebb a C-vitamin hiánya miatt fellépő kollagénszintézis zavara, mely az érfalakat rendkívül sérülékennyé teszi (skorbut).
A vérlemezkék problémái
Számbeli csökkenés
Funkciózavar
Véralvadási faktorokkal kapcsolatos zavarok
Szerzett véralvadási zavarok
Ezek gyakoribbak, mint az öröklöttek. Ilyen pl. a K-vitamin-hiány, amely a májban zajló protrombin szintézisében okoz zavarokat.
Örökletes véralvadási zavarok, a hemofília
A hemofília ritka, veleszületett, egész életen át tartó vérzékenység, mely az egyes véralvadási faktorok rendellenes képződésével vagy hiányával kapcsolatos. A betegség lényege a vér alvadékonyságának csökkenése, melynek következtében a hemofíliás betegek sérülések alkalmával hosszabb ideig véreznek, mint az egészségesek.
A leggyakoribbak és legsúlyosabb problémát az ízületi- és izomvérzések jelentik. A belső vérzések által okozott feszítő nyomás miatt a hemofíliát az egyik legfájdalmasabb betegségnek tekinti az orvostudomány. A gyakori bevérzések következtében a betegek ízületei fokozatosan tönkremennek és izmaik sorvadnak.
Az alvadási faktorok mindegyikének leírták örökletes hiányállapotát. Leggyakoribb a
 |
Trombózis
Kórosan fokozott véralvadási zavar, a rendellenesség következtében létrejövő vérrög kitapad az érfalra, aminek következtében kisebb-nagyobb ereket akár el is zárhat. A vérrög (vércsomó, trombus) a véralvadási rendszer aktiválódása révén, a vérlemezkék összecsapódása miatt jön létre. A trombusok leggyakrabban az alsó lábszár mélyvénáiban alakulnak ki.
Mélyvénás trombózisban a lábak mélyben futó, nagy visszerei, vénái zárodnak el. A mélyvénás trombózis az érintett végtag fájdalmas duzzanatát okozza.
A mélyvénás trombózis tünetei
A trombózis okai:
Kialakulására hajlamosítanak:
Embólia
Trombózisról akkor beszélünk, ha valahol az érrendszerben vérrög (trombus) képződik. A trombus növekszik, és előfordul, hogy egy kisebb-nagyobb darabka leszakad belőle. Ezt a leszakadt vérrögöt – embolust - azután a véráram tovább sodorja egészen addig, amíg átfér az ereken. Amikor egy nála kisebb érszakaszhoz érkezik, megakad, és elzárja a vér útját. Ekkor beszélünk embóliáról.
Leggyakoribb a tüdőembólia, mivel a láb mélyvénáiból leszakadt embólus, a nagyvérkör vénás rendszerében haladva egyre nagyobb átmérőjű erekbe kerül, majd a szív üregein – jobb pitvar, jobb kamra – áthaladva a kisvérkör fokozatosan szűkülő ereiben valahol megakad.
Tünetei a hirtelen jelentkező éles, heves mellkasi fájdalom, légszomj, véres köpet. Az, hogy a tüdőembólia milyen súlyos, az embolus, illetve az elzárt ér nagyságától függ. Lehet apró, szinte észrevétlen, de nagyobb embolus esetén életveszélyes állapot is kialakulhat.
35. tétel: A szívműködés |
A szív a vérkeringés központi szerve. Az emberi szív ököl nagyságú szerv. A mellüreg középső részén, kissé bal oldalon található, a két tüdő között, a mellhártyák által határolt üregben.
A szív falának rétegei
A szív falát három réteg alkotja (a vérerekét szintén).
Külső réteg: a szívburok
Kétrétegű savós hártya. A külső réteg zsákszerűen veszi körül a szívet, a belső vékony réteg a szív felszínére tapad. A szívburok két lemeze között van a szívburoküreg, melyet súrlódáscsökkentő savós folyadék tölt ki.
A szív felszínén futnak zsírszövettel körülvéve, az aorta kezdetén eredő, a szívet tápláló ún. koszorúerek (madarakban, emlősökben). A zsírszövet túltápláltság esetén felszaporodhat, amely nehezíti a szív mozgását. A koszorúerek az aorta kezdetéről erednek és a jobb pitvar előtt csatlakoznak vissza a szívbe, teljes mértékben behálózzák a szívet. A koszorúerek megfelelő állapota alapvető feltétele a szív optimális működésének. Túl zsíros táplálkozás esetén az erek belső falára koleszterin tartalmú meszes anyag rakódik le. Ez csökkenti az erek átmérőjét, rontja a szív vérellátását, a szívizomzat elhalhat, melynek következménye az infarktus. A nikotin érszűkítő hatása miatt a dohányzás ezt tovább súlyosbítja (lásd még egészségtan).
A szív vastag középső rétege: a szívizomzat
A szív legbelső rétege: a szívbelhártya
 |
A szív felépítése
A szívben négy üreg található:
A szívet egy függőleges válaszfal jobb-, ill. baloldali részre osztja. Embrionális korban a két pitvar között nyílás található, amely azonban a születés után bezáródik, eltűnik.
A magzat vérének gázcseréjét, táplálékfelvételét, az anyagcseretermékek kiürülését a méhlepény közvetítésével az anyai szervezet végzi. A fejlődő tüdő légtelen, benne nincsen gázcsere, a tüdő vérkeringése, a kisvérkör, tehát nem működik, keringése minimális mértékű.
A pitvarok fala igen vékony a kamrákéhoz képest. A bal kamra fala jóval vastagabb a jobb kamráénál, mivel jóval nagyobb munkát végez (belőle indul ki a nagyvérkör).
A belépő (vénák) és a kilépő (artériák) erek a szív felső részén találhatók.
A vér áramlása egyirányú, a pitvaroktól a kamrák felé történik. A vér mozgásának irányítását a szelepszerűen működő szívbillentyűk biztosítják.
A szívben kétféle billentyű található, amelyek a szívbelhártya származékai:
A vitorlás billentyűk
A vitorlás billentyűk vitorláihoz a kamra felől ínhúrok kapcsolódnak, amelyek a kamra falából kiemelkedő szemölcsizmokról erednek.
A vitorlás billentyűk akkor nyílnak meg, ha a pitvarban nagyobb a nyomás, mint a kamrában, ekkor a vér a pitvarokból a kamrákba áramlik. Ellenkező esetben a billentyűk bezáródnak, a pitvarok felé a túlhajlást az ínhúrok megfeszülése és a szemölcsizmok összehúzódása akadályozza meg.
A zsebes billentyűk
A szív zsebes billentyűi a kamrák és az artériák határán találhatók (zsebes billentyűk még a végtagok és a nyaki vénák falában is előfordulnak). Ha a kamrákban nagyobb a nyomás, mint az artériákban, akkor összelapulva megnyílnak. Ellenkező esetben a három hártyás zseb megtelik vérrel, kidomborodik és összehajolva megakadályozzák a vér visszaáramlását a kamrákba.
Szívhangok
A billentyűk záródásakor jönnek létre.
„Ha a szív bal kamrájába nyíló, úgynevezett kéthegyű vitorlás billentyű a normálisnál lazább szerkezetű, akkor a billentyű kamrai összehúzódáskor túlmozdul, és kis záródási elégtelenség alakulhat ki. Ennek következménye az, hogy kis mennyiségű vér visszaszivárog a pitvarba és kialakul az úgynevezett mitralis billentyű elégtelenség. Ez légszomjjal, fáradtsággal jár, mivel a pitvarba visszaáramló vér akadályozza a tüdővénák ürülését, ill. a tüdőben a vénás vérnyomás növekedése és a szövetközötti folyadék felszaporodása nehezíti az oxigénellátást. A rendellenesség súlyosabb esetben műtéti beavatkozást tesz szükségessé.” Emelt szintű érettségi feladat
 |
A szívműködés folyamata
A szívműködés folyamatában alapvetően két fázist különböztetünk meg:
A vér a pitvarok felől a kamrák felé áramlik, mozgásának irányát a nyomáskülönbség határozza meg, mindig a nagyobb nyomású hely felől a kisebb nyomású tér felé halad.
Pitvar diastole során a nagyvénák felől vér áramlik a pitvarokba.
1. Pitvar szisztole
2. Izovolumetriás/változatlan térfogatú kontrakció/összehúzódás
3. Kamrai vér kiáramlás (gyors majd csökkent ejekció)
4. Izovolumetriás/változatlan térfogatú elernyedés
5. Passzív kamrai telítődés
 |
Tehát a pitvarok összehúzódásakor a kamrák elernyedt állapotban vannak, illetve fordítva.
Egy szisztoléból és egy diasztoléból álló esemény sorozatát szívciklusnak nevezünk. A szív ritmusos összehúzódásaival pumpálja a verőerekbe a vért. A bal kamra a nagy-, a jobb kamra pedig a kisvérkörbe.
A szív ritmusos percenkénti összehúzódásának számát, a szívfrekvenciát a pulzusszámmal jellemezzük. A pulzushullám az artériák falán végigterjedő nyomás- és térfogatváltozás, mely leginkább a nagyvérkör nagy, közép, és kis artériáin érzékelhető.
A szívfrekvencia nyugalmi állapotban átlagosan
Az egy összehúzódás alatt kipumpált vér mennyisége a pulzustérfogat (verőtérfogat), felnőtteknél nyugalomban átlag kb. 60-80 ml kamránként.
A pulzustérfogat és a szívritmus szorzata a perctérfogat.
Perctérfogat = pulzustérfogat x frekvencia
Nyugalomban a bal kamra perctérfogata: 70 ml x 72 = 5040 ml. A két kamra által továbbított vér mennyisége természetesen azonos.
A szív ingerképző és ingerületvezető rendszere
A szív automatikus működésű. Idegi összeköttetéseitől megfosztva is működőképes, mivel önálló ingerületgeneráló és -vezető rendszere van. Ez a rendszer speciálisan módosult szívizomsejtekből áll.
A rendszer részei:
A szinuszcsomó az elsődleges ingerképző központ, a jobb pitvar falában található. Percenként 72 impulzust – akciós potenciált - generál nyugalomban. A pitvar izomzata összehúzódik, és a pitvar-kamrai csomóhoz vezeti az impulzusokat.
A pitvar-kamrai csomó másodlagos ingerképző központ. A szinuszcsomó sérülése esetén percenként 45 impulzust generál.
A pitvar-kamrai csomóból az impulzusokat a kamrák felé a
A szívizomsejtek között elektromos szinapszisok vannak, aminek köszönhetően a szívizomtömeg egyszerre ingerelhető, ugyanolyan mértékű összehúzódást kiváltva. Mindezek eredményeképp a szív a pitvarok felől a kamrák felé húzódik össze.
A szívizomban nem váltható ki tartós összehúzódás, az összehúzódás alatt a szívizom ingerelhetetlen. (Oka: az elhúzódó Ca2+ beáramlás miatt lassan áll vissza a nyugalmi potenciál.)
A szív működésekor keletkező elektromos impulzusokat a bőr felszínén mérni lehet. Ennek grafikus megjelenítése az EKG. EKG-vizsgálat során a szív elektromos aktivitásán keresztül következtetünk a szív állapotára.
A szívműködésre az idegrendszer és a hormonális rendszer gátló, ill. serkentő hatást gyakorolhat. A vérkeringés – szívműködés, vérnyomás – működésének szabályozó központjai a nyúltvelőben találhatók.
A vérkeringés – a vérnyomás és a szív működésének - szabályozása
A vérkeringés szabályozásában hormonális és idegi szabályozó rendszerek vesznek részt.
A szívműködés szabályozása lényegében a szinuszcsomó működésére gyakorolt serkentő, ill. gátló hatások révén történik.
Az idegi szabályozás
Az idegi szabályozóközpontok az idegrendszerben számos helyen megtalálhatók, működésük hierarchikus szerveződésű.
A) Az elsődleges központok az agytörzsi hálózatos állomány nyúltvelői szakaszában helyezkednek el. Itt kétféle, ellentétes hatású központ van.
B) A magasabb agyi központokból (limbikus rendszer) származó ingerületek pl. izgalom, düh, félelmek, szorongások az elsődleges központokon keresztül fokozó hatást – vérnyomás emelkedést, szívfrekvencia növekedést - gyakorolnak a keringésre.
Aortaív reflex
A legfontosabb vérnyomás szabályozó a mindennapi életben!
Szívinfarktus
A szívinfarktus oxigénhiány és tápanyaghiány következtében létrejött szívizompusztulást, ill. az ezeknek következtében kialakuló szívműködési zavarokat jelenti. A koszorúerekben lerakódott meszes plakkok szűkítik a szív ereit, a szűkületben vérrög alakulhat ki, mely teljes elzáródást okozhat.
A súlyos szűkület vagy a teljes elzáródás miatt a szívizomsejtek vérellátása kórosan lecsökken. Ha ez a folyamat hosszú ideig tart, a károsodás visszafordíthatatlanná válik, s kialakul a szívinfarktus.
A fő kockázati tényezők a koszorúér-betegség kialakulása szempontjából:
Tünetei
Egy érgörcs is vezethet szívinfarktushoz, hisz a szív vérellátása ez által is csökken. Különösen elmeszesedett erekben jön létre, a simaizomsejtek összehúzódásával.
Az angina a szív oxigénhiánya miatt jelentkező erős mellkasi fájdalomérzet. A fájdalom típusosan a mellkas középső részén érződik, tehát a szegycsont mögött, kisugározhat a bal vállba, bal karba, de az állkapocsba, illetve a bal lapockába is.
Ritmuszavar
A szívritmuszavar, másképpen aritmia, a szívverés ütemének megváltozását jelenti.
A normálisnál
Pitvarfibrilláció (pitvarremegés)
A pitvarfibrilláció a szív pitvarainak szabálytalan, gyors összehúzódásából eredő ritmuszavar, melynek eredményeképpen a kamra és a pitvar egymástól függetlenül húzódik össze. A pitvarok több pontján, rendszertelenül keletkeznek ingerületek, melyek hatására a kamrák szabálytalan ritmusban és legtöbbször túl gyorsan húzódnak össze. Így a kamrákból kevesebb vér kerül a vérpályákba, mint normális ritmus esetén, ami a szervek vérellátási zavarát okozza. Jellegzetes tünetei lehetnek, légszomj, fáradtságérzés, szédülés, szabálytalan, gyors szívdobogás érzése.
Kamrafibrilláció
A kamrai ritmuszavar esetén a kamrák falában található izomsejtek összehúzódása gyors, nem összehangolt, melynek következtében nincs hatékony összehúzódás. A hirtelen szívhalál hátterében, az esetek többségében (70%) a kamrai ritmuszavar, ún. kamrafibrilláció áll.
Eszméletvesztéssel és életveszéllyel járó állapot, ami azonnali orvosi beavatkozást kíván. Ezt a ritmuszavart defibrillátor kezeléssel kell rendezni. A defibrillátor elektromos impulzust ad le a szívnek, mellyel a szív kaotikus elektromos aktivitását megszünteti és ezáltal lehetőséget biztosít a normális szívritmus visszatérésére. Amennyiben a szívműködés nem indul újra, meg kell kezdeni az újraélesztést. A tévhittel ellentétben a készülék a leállt szívet nem indítja újra.
Ritmusszabályozó (pacemaker)
A szív természetes ingerképzését pótló kis, teleppel működő elektromos eszköz, amit a bőr alá ültetnek be. Rendszerint akkor szükséges, ha a szív természetes ritmusa túl lassú, vagy a szív ingerületvezető rendszerében hiba van, így a kamrák és pitvarok aktivitása nincs megfelelően összehangolva. A beültetett ritmusszabályozó hosszú éveken át biztonságosan vezérli a szív összehúzódásait.
36. tétel: Vér- és nyirokkeringés |
A vérerek felépítése és működése
A gerincesek - így az ember - keringési rendszere zárt. A vérkeringés központja, motorja a szív.
A szívből két vérkör indul ki.
Az aorta rövid felszálló ág után ívben visszahajlik, s érintve a nyelőcsövet és a légcsövet a szív mögött kissé balra a leszálló aortában folytatódik, amely oldalágakat ad le a különféle belső szervek felé, mint pl. máj, bélcső, vese stb. majd a 4. ágyékcsigolya magasságában kettéágazik az alsó végtagokat ellátó közös csípőartériákra. Az aortaív kezdetén erednek a szívet ellátó koszorúerek.
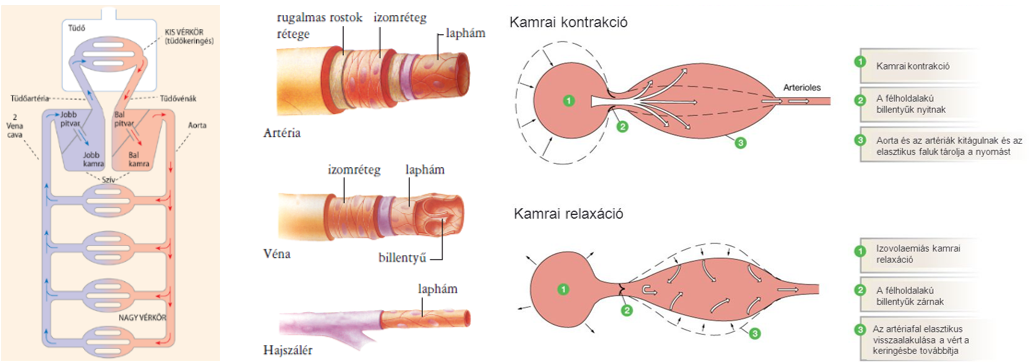 |
Az erek falának szerkezete
A vérerek a valódi kapillárisok kivételével 3 rétegből állnak.
Az egyes rétegek vastagsága, szerkezete az erek nagyságától és feladatától függően változik.
Az artériák szerkezete
Az artériák a szívtől szállítják el a vért. A nagyvérkörben oxigénben gazdag, a kisvérkörben oxigénben szegény vért tartalmaznak.
Az egyes artériák átmérője a szívtől távolodva egyre csökken, ami alapján megkülönböztetünk nagy, közép, kis artériákat és a legkisebb arteriolákat.
Szövettani metszeteken az artériák keresztmetszete kör alakú, mivel faluk vastag, igen rugalmas, így nem nyomódnak össze.
Az artériák falában megtalálható rugalmas rostoknak és a simaizomszövetnek köszönhetően a bennük folyó vér áramlását folyamatossá teszik, a szív felől jövő lökéshullámokat (pulzus) ha nem is kiegyenlítik, de mérsékelik. A bal kamra pulzustérfogatának nagy része a systole időtartama alatt nem hagyja el az aortát, hanem kitágítja azt, és a diastole alatt áramlik tovább a többlet vérmennyiség (szélkazán funkció).
A vénák felépítése
A vénák a vért a szív felé szállítják a kapillárisok felől.
A vénák fala a hasonló keresztmetszetű artériákhoz képest vékonyabb, mivel a középső réteg sokkal kevesebb simaizmot tartalmaz. Másik eltérés, hogy sokkal kevesebb a rugalmas rost mennyisége, így a vénák kevésbé rugalmasak, mint az artériák, faluk tágulékony.
Szövettani készítményeken a környező szövetek nyomásának hatására a fenti okok miatt a vénák keresztmetszete általában lapított, ovális.
A végtagok és a nyak vénáiban a vér áramlását zsebes billentyűk segítik, amelyek megakadályozzák a vér visszaáramlását. A vénás vér áramlását a vázizmok ritmikus összehúzódása az izompumpa is segíti, úgy, hogy az izmok összehúzódásakor a bennük átmenő ereket összenyomják, azonban a billentyűknek köszönhetően a vér csak a szív irányába képes továbbhaladni.
A legkisebb vénák a venulák, majd tovább a szív felé haladva az erek átmérője fokozatosan nő.
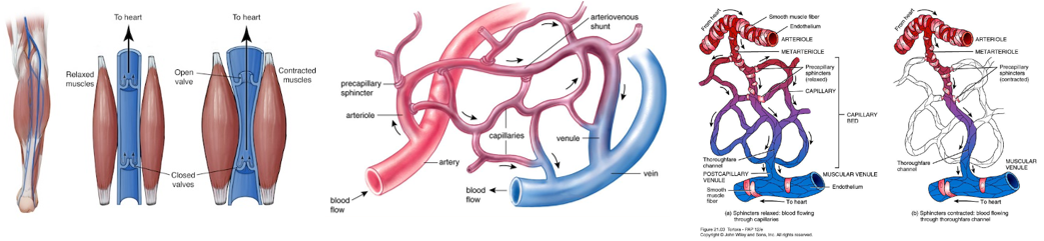 |
A kapillárisok szerkezete
A legkisebb artériákat az arteriolákat és a legkisebb vénákat a venulákat a kapillárisok hálózata kapcsolja össze.
A hajszálereknek két típusát különböztetjük meg.
A kapillárisok fala igen vékony, már csak egyrétegű laphámsejtek és alaphártya alkotja.
A valódi kapillárisok ágainak eredésénél kis simaizomgyűrűk találhatók, melyek segítségével szabályozható a véreloszlás a hajszálérhálózaton belül.
A vérnyomás
A vérkeringési rendszert az erekben keringő vér érfalra gyakorolt nyomása alapján - amit vérnyomásnak nevezünk - két szakaszra osztjuk:
A vérnyomás értékét a légköri nyomás értékéhez viszonyítjuk, úgy, hogy annak értékét - 760 Hgmm, 101 kPa – 0-nak vesszük.
A magasnyomású rendszer a bal kamrával kezdődik, a nagyvérkör artériás rendszerével folytatódik és az arteriolák rendszeréig tart. Mint, ahogy korábban tárgyaltuk a szív bal oldalán:
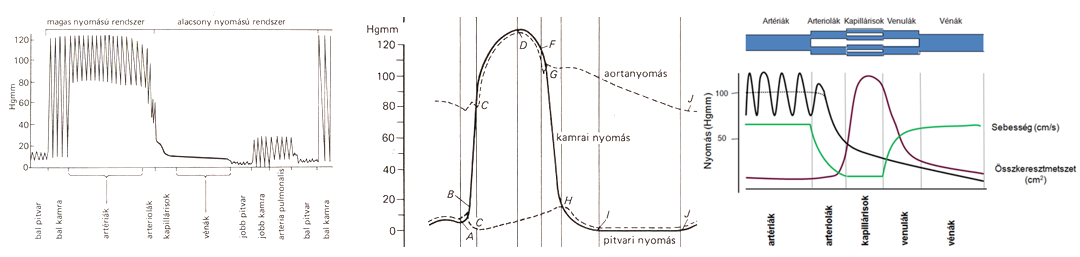 |
Az aortában mért
A kamra összehúzódásakor az artériás rendszerbe került vér nyomáshullámot – pulzust - indít el, amely egyre csökkenő mértékben az arteriolákig jellemző.
Alacsony nyomású rendszer
A véráramlás fizikai jellemzői, vérnyomás, véráramlási sebesség
A véráramlás egyirányúságát, ill. a folyamatosságát az érpálya két végpontja között mérhető nyomáskülönbség tartja fenn. A vérnyomás tehát a nagyvérkörben a bal kamrától a jobb pitvarig, ha nem is egyenletesen, de folyamatosan csökken.
Az erek átmérője folytonosan és jellegzetesen változik a keringési rendszerben, az artériás rendszerben egyre csökken a kapillárisok felé haladva, a vénás rendszerben egyre nő a szívfelé haladva. Az egyedi érátmérő változásával fordítottan változik az erek összkeresztmetszete.
Az erek összkeresztmetszete tehát
A keringési rendszerben az áramlási ellenállás mértékét több tényező befolyásolja, mint pl. az erek átmérőjének változása, vagy a vér viszkozitása (belső súrlódása). Az erek összkeresztmetszetének növekedésével az áramlási ellenállás egyre nő.
Ugyanakkor az ellenállás fokozódásával párhuzamosan csökken a vér áramlási sebessége. Tehát a vér áramlási sebessége fordítottan arányos az erek összkeresztmetszetével, egyenesen arányos az erek átmérőjével.
A véráramlás sebességének a csökkenése kedvez a kapillárisok területén bekövetkező anyagforgalomnak.
Összefoglalva a véráramlás egyirányúságát biztosítja:
A szív által továbbított véráram számára sem az aorta, sem a nagyobb artériák nem jelentenek jelentős áramlási ellenállást. Ezért az ezekben bekövetkező vérnyomásesés is elhanyagolható. A legnagyobb perifériás áramlási ellenállást a kapillárisok előtti arteriolák jelentik. A vér így mintegy a szív és az arteriolák közé "szorul", ami nagy vérnyomást eredményez.
Véreloszlás
A szervezetünkben nincsen minden kapilláris egyszerre nyitva, a szervek működésétől függően a vér különböző mértékben oszlik meg az egyes szervek között.
Az egyes szervek vérellátottságának a mértékét a perctérfogattal fejezzük ki, amely az adott szerven az egy perc alatt átfolyó vér térfogatát jelenti.
A véreloszlást az egyes szervek között a hormon- és az idegrendszer szabályozza, melyek hatásukat az erek simaizomzatán keresztül fejtik ki (értágítás - érszűkítés).
A működő szervek fokozott vérellátása úgy valósul meg, hogy
A keringési szervrendszer alkalmazkodása a fizikai munkavégzéshez
Fokozott terhelés esetén mind a szívfrekvencia, mind a verőtérfogat megnő, így egy átlagember estén a perctérfogat elérheti akár a 21 dm3/perc értéket is (120 cm3 x 180/perc).
Fizikai aktivitás közben nő a verőtérfogat, amíg a szív el nem éri a 120/perces szívfrekvenciát. Ezt követően a szívfrekvencia tovább nőhet a terhelés folytatása esetén, azonban a verőtérfogat a frekvencia növekedésével már nem nő párhuzamosan (annyira lecsökken a kamratelítődés ideje). 1 ml = 1 cm3
Edzés hatására nő a szívizom összehúzódó képessége, a szív bal kamrája több vért képes befogadni, mindennek következményeként megnő a verőtérfogat (az edzetlen ember szívének akár 1.5 x-ére, 170 cm3-re).
A sportolás közbeni magas pulzusszám és a megnőtt verőtérfogat eredményezi, hogy ilyenkor a perctérfogat is jelentősen megnő, a nyugalmi 5 l-ről akár 30 l/perc fölé is.
Edzett egyén a fizikai munkához tehát elsősorban a pulzustérfogat növekedésével alkalmazkodik, kisebb mértékben nő a szívfrekvencia, mint edzetlen egyén esetében.
Rendszeres edzés hatása abban is megmutatkozik, hogy fokozatosan megnő a szívizom tömege, emiatt lassabb a sportolók nyugalmi pulzusa.
Fizikai munkavégzés során ugyanakkor a terheléssel arányosan nő az artériás középnyomás.
A vázizomzat és a bőr ereiben nagyfokú értágulat, így áramlásfokozódás jön létre, ezek a területek intenzív fizikai munka során a perctérfogat 80%-át is kaphatják, ugyanakkor a bélrendszer keringése jelentősen csökken, emiatt nem következik be általános vérnyomásesés.
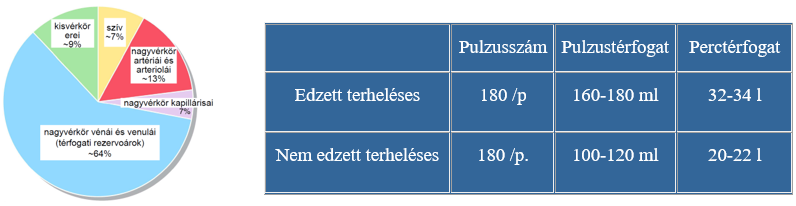 |
A szöveti keringés, nyirokkeringés
Mint már korábban volt róla szó, a zárt keringési rendszerrel rendelkező gerinces állatok és az emberi szervezet 3 folyadékteret foglal magába.
E tereket különféle határoló hártyák - érfal, sejthártya - választják el egymástól, melyeken keresztül élénk anyagforgalom zajlik a folyadékterek között.
A nyirok a szövetnedv nyirokerekbe kerülő része. A szövetnedv és a nyirok a szervezet összes víztartalmának kb. 20-30%-át teszi ki.
A szövetnedv
A szövetnedv képződése
A vérerek közül kizárólag a kapillárisok fala átjárható a víz és a benne oldott kismolekulájú anyagok számára.
A kapillárisok fala vékony, szűrőként működik, féligáteresztő, mivel
A szűrés során lényegében fehérjementes vérplazma jön létre. A nyirokban található fehérjéket – pl. immunfehérjék – az immunrendszer sejtjei termelik.
A folyadék mozgásának az irányát két ellentétes hatás eredője szabja meg.
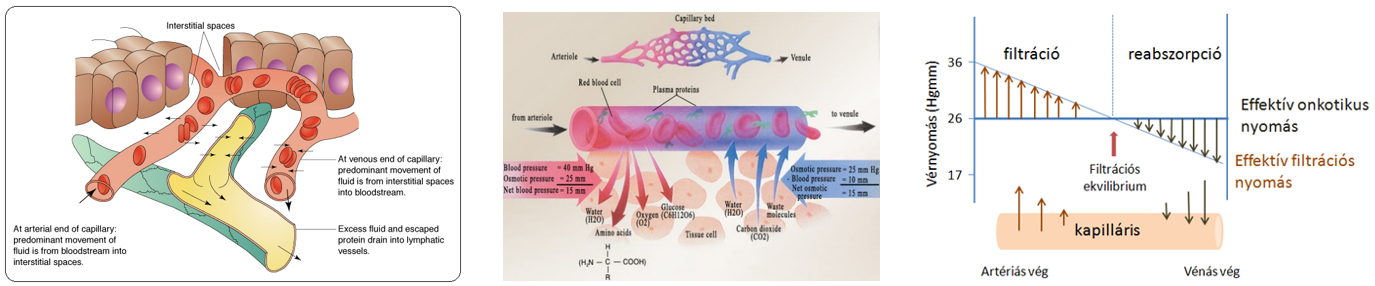 |
1. Az erekben uralkodó vérnyomás, amelynek értéke a kapillárisok kezdetén 35 Hgmm, a folyadékot kifelé préseli.
2. Ennek ellene hat a plazmafehérjék – főleg albuminok – kolloidozmotikus nyomása – ozmotikus szívóereje -, amely 25 Hgmm.
Az ún. effektív (nettó) filtrációs nyomás (Peff= Ph-Pkoll) a két nyomásérték különbsége.
A kapilláris artériás végén a filtráció miatt a plazmafehérjék koncentrációja és így az onkotikus nyomás (plazmafehérjék ozmotikus nyomása) kismértékben nő (1-2 Hgmm), de a változás csekély, ezért az élettani leírásokban gyakran állandónak tekintik.
Tehát abban az esetben, ha Peff pozitív, a folyadék kiszűrődik a szövetek közé, azonban, ha Peff negatív, a szövetnedv egy része visszalép a vérbe. Így működnek az abszorptív kapillárisok, mint pl. a nagyvérköri szöveti kapillárisok.
Abban az esetben azonban, ha a vérnyomás a kapillárisban végig meghaladja a vér kolloid ozmotikus nyomását, nincs folyadék visszaszívódás. Így működnek az ún. nem abszorptív kapillárisok, mint pl. a vesetestecskékben található érgomolyagok.
A szövetnedv oldott vérfehérjéket nem tartalmaz, ezért kolloid ozmotikus nyomása alacsony.
A szűréssel képződött szövetnedv nem képes teljesen visszaszívódni az érpályába, egy része kint marad a szövetek között, aminek napi mennyisége néhány liter, melyet a nyirokkeringés juttat vissza az vérbe.
A nyirok összetétele hasonló a vérplazmáéhoz a plazmafehérjék kivételével. Tartalmaz vizet, sókat, glükózt, aminosavakat, zsírokat, hormonokat, immunfehérjéket, bomlástermékeket, ezeken kívül nyiroksejteket.
A bélben felszívódott zsírok elsősorban a bélbolyhok nyirokereibe kerülnek, amelyekből a vénás rendszerbe jutnak.
Tehát a nyirokrendszer feladata a
Abban az esetben, ha a nyirok képződése zavart szenved ödéma alakul ki, amely a pangó szövetnedv felhalmozódásának köszönhető. Az ödéma oka igen változatos.
A nyirok igen lassú áramlásának egyirányúságát
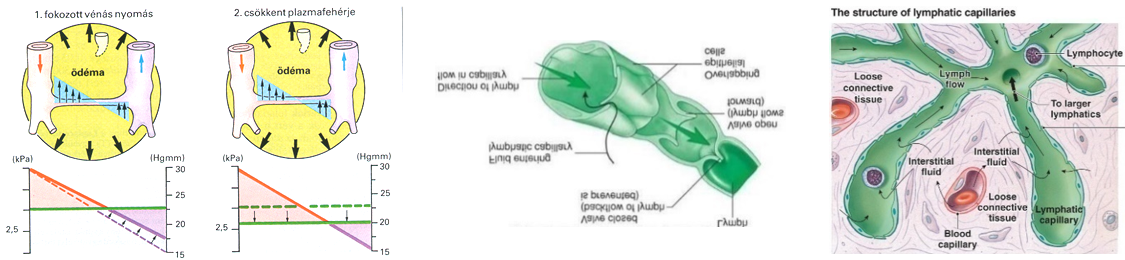 |
A nyirokrendszer részei
A nyirokérrendszer vékony falú, vakon kezdődő nyirok kapillárisokkal indul a szövetekben, majd egyre vastagabb nyirokerekké egyesülve vezetik a nyirkot a vérpályába. A legfőbb nyirokér a – a mellvezeték - jobb pitvar előtt torkollik a vénás rendszerbe. Ugyanakkor nincsenek nyirokerek az idegszövetben.
A nyirokcsomók
A vénákhoz hasonló falszerkezetű nyirokerek összefutási pontjaiban babalakú, borsónyi nyirokcsomók találhatók. Nagyjából egyenletesen helyezkednek el, azonban nagyobb mennyiségben fordulnak elő az ágyéki tájékon, a gerinc mellett, a hónaljban, a nyakon, az állkapocs alatt, a fül tövében.
A nyirokcsomók feladata többrétű:
A nyirokcsomó összegyűjti a kisebb nyirokereket, majd a befolyt nyirok nagyobb nyirokéren távozik. Kívülről egy kötőszövetes rostos tok határolja. Belül nyirokszövet építi fel, amely külső kéregállományra és belső velőállományra különül. Az egész belső állomány szivacsszerű, tele fagocitáló sejtekkel, melyek az áthaladó nyirkot tisztítják.
A kéregállományban apró gömbszerű képződmények, ún. nyiroktüszők találhatók. A nyiroktüszők érett nyiroksejtek halmazai. A nyiroktüszőben az érett nyiroksejtek találkoznak a kórokozókkal, aminek hatására aktiválódnak. Az aktiválódott sejtek sokszoros osztódásba kezdenek, olyan sejteket létrehozva, amelyek már felismerik az antigént, ill. memória sejteket termelnek, amelyek évekig képesek tárolni az antigén jellemzőit. A nyiroktüszők nemcsak a nyirokcsomókban fordulnak elő, hanem számos helyen elszórva vagy csoportosan a szervezetben pl. a lépben, a mandulákban, a bélcső falában (főleg a féregnyúlvány területén).
 |
A mandulák
A mandulák a tápcsatorna elején – orrgarat, nyelv - a nyálkahártyában megtalálható nyiroktüszők csoportosulásai. A nyiroktüszők tömege olyan nagy, hogy a nyálkahártyát kidomborítják, mivel több rétegben, ugyanakkor szabálytalan halmazban fordulnak elő.
Saját kötőszövetes tokjuk nincs, a nyálkahártya mély járatokkal választja el a nyiroktüsző csoportokat. A termelődő hatalmas mennyiségű nyiroksejt átlép a hámszöveten és részt vesz a behatoló antigénekkel szembeni védekezésben. Ennek köszönhető, hogy a szájban fellépő sérülések viszonylag gyorsan gyógyulnak. Ugyanakkor a nyálkahártya hézagaiban speciális baktériumok telepedhetnek meg, jellegzetes mandulagyulladást okozva.
A csecsemőmirigy
A csecsemőmirigy vagy thymus közvetlen a szegycsont mögött, a szív felső részén elhelyezkedő lapos szerv. Újszülötteknél kb. 10 g, majd serdülőkorban éri el a legnagyobb fejlettségét, végül felnőtt korban szinte elcsökevényesedik.
Még az embrionális fejlődés során a thymusba vándorolt T-sejtek itt fejezik be érésüket, melynek során megtanulják megkülönböztetni a saját anyagokat az idegentől. Ezek után innen jutnak a másodlagos nyirokszervekbe, ahol találkoznak az antigénekkel és felismerve azokat aktiválódnak.
A lép
A szervezet legnagyobb nyirokszerve, azonban szemben a nyirokcsomókkal a vérpályához tartozik, a nyirokerek elkerülik.
Feladata többrétű.
A hasüreg baloldalán található, az alsó bordák magasságában. Dinnyeszelethez hasonló alakú szerv. Kívülről kötőszövetes tok határolja, amelyben több-kevesebb simaizomszövet is megtalálható. Ennek elsősorban olyan ragadozóknál van jelentősége, amelyek hirtelen nagy erőkifejtésre képesek, így szükség esetén a lép jelentős mennyiségű vért juttathat a vérpályába.
 |
37. tétel: A veseműködés és szabályozása |
Bevezetés
A kiválasztás minden állati szervezet alapvető önfenntartó működései közé tartozik,
A kiválasztó működés
Tágabb értelemben kiválasztó működést lát el
Szűkebb értelemben azonban csak a kiválasztószervek, a vesék végeznek kiválasztást. A vesék működésükkel biztosítják a szervezet belső egyensúlyát, mivel eltávolítják a felesleges vizet, ionokat, mérgező bomlástermékeket, ezáltal kialakítják a belső környezet – a testfolyadékok – optimális összetételét. Továbbá a vese a benne termelődő szöveti hormonok révén részt vesz a vérnyomás szabályozásában, ill. a D-vitamin aktív formájának előállításában.
A szervezet szabályozott belső egyensúlya a homeosztázis lásd még 4.1.1. fejezet
A többsejtű állatokban a sejtek többsége már nem érintkezik a külvilággal, számukra a környezetet a testfolyadékok jelentik (a szövet közti folyadék és a vér). Ezek a folyadékterek belső környezetet teremtenek a sejtek számára. A belső környezet legfontosabb feladata, hogy biztosítsa a sejtek működéséhez a megfelelő feltételeket az anyagcsere-folyamataihoz szükséges anyagok odaszállításával, ill. a bomlástermékek eltávolításával.
A belső környezet fontos tulajdonsága összetételének viszonylagos állandósága, ami igen bonyolult szabályozás révén valósul meg. A belső környezet szabályozott dinamikus állandóságát homeosztázisnak nevezzük. A homeosztázisnak köszönhetően a sejtek számukra optimális környezetben működhetnek. A belső környezet legfontosabb szabályozott paraméterei a következők:
E paraméterek viszonylagos állandósága a vese működésével biztosíthatók. Az egyes paraméterek értékei fajonként eltérőek lehetnek. Fejlettebb élőlényeknél a testfolyadékok tápanyag-koncentrációja és a testhőmérséklet is a szabályozott tényezők közé tartoznak.
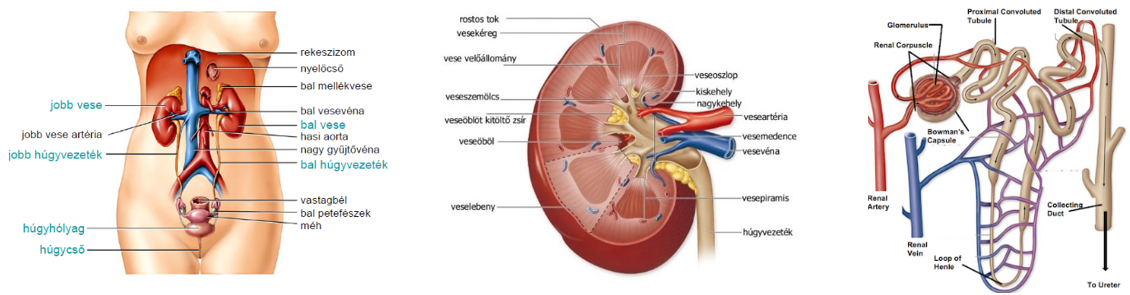 |
Az ember kiválasztó-szervrendszerének a részei:
A vese feladatai
A vese felépítése
A vesék a hasüreg hátsó falán, a gerinc két oldalán, az alsó bordák magasságában helyezkednek el. A vesék alakja babhoz hasonlít. A ‘bab’ homorulatában található a vesekapu, ahol a vesét ellátó erek, idegek lépnek be, ill. ki és itt hagyja el a vesét a húgyvezeték. A vesék fej felé eső csúcsán sapkaszerűen helyezkednek el a mellékvesék, kiválasztást nem végeznek, belső elválasztású mirigyek.
A vese hosszmetszeti képén jól látszik, hogy nem tömör, hanem üreges szerv,
A karéjszerűen elhelyezkedő veseszövetben
A kéregállomány oszlopként benyomul a velőállományba, és azt vesepiramisokra tagolja. A vese központi ürege a vesemedence, elkeskenyedve, éles határ nélkül megy át a húgyvezetékbe.
A vese mikroszkópos szerkezete, a nefronok felépítése és működése
A vese anatómiai és funkcionális egysége a nefron, számuk egy vesében kb. 1,5 millió. A nefron
A vesetestecske (Malpighi-féle vesetestecske) a kéregállományban helyezkedik el,
Az érgomolyag egy speciális hajszálérhálózat, mivel az odavezető arteriola kapillárisokra történő oszlása után újra arteriolává szedődik össze. A kilépő arteriola átmérője kisebb, mint az odavezető éré, aminek a jelentősége abban áll, hogy a szűrőműködéshez megfelelően magas marad a vérnyomás a kapillárisban, ill. a jelentős mennyiségű folyadékkilépés – szűrőműködés – ellenére sem esik a vérnyomás jelentősen ezen az érszakaszon. Az érgomolyag kapillárisait pórusos laphámsejtek bélelik, rajtuk 50-100 nm széles nyílások találhatók.
Az érgomolyagból kilépő arteriola rövid lefutás után ismét hajszálerekre bomlik, melyek harisnyaszerűen veszik körül ugyanannak a nefronnak az elvezető csatornáját. Ezek a kapillárisok összeszedődve már vénákká egyesülnek, amik végül a vesekapun keresztül hagyják el a vesét.
A kehelyszerű Bowman-tok érgomolyag felé eső részét speciális laphámsejtek, ún. lábassejtek bélelik egy rétegben, melyek nyúlványos sejtek, nyúlványaik nem érnek össze, közöttük kisebb-nagyobb rések vannak, ezeken keresztül az érgomolyag alaphártyája kapcsolatban van a Bowman-tok üregével. Így az ér és a Bowman-tok ürege között csupán az alaphártya képezi a válaszfalat, ami egyben szűrőberendezésként is működik.
Az elvezető csatorna felépítése és működése alapján 3 további szakaszra tagolható,
A Bowman-tokból eredő közeli csatorna zömmel a kéregállományban található. A csatorna falát működésének megfelelően – felszívás – mikrobolyhos egyrétegű hám alkotja.
A proximális tubulus belépve a velőállományba egy hosszú, sugárirányú, majd hajtűszerűen visszakanyarodó csőben, a Henle-kacsban folytatódik. A távoli csatorna a Henle-kacs felső részének a folytatása, szintén a kéregállományban található.
A nefronok disztális tubulusai a velőállományban sugarasan futó gyűjtőcsövekbe nyílnak. Átlagosan 6 nefron csatlakozik a gyűjtőcsatornába. Több gyűjtőcsatorna közös kivezetőcsőbe vezet, melyek többedmagukkal a veseszemölcsök csúcsán nyílnak a vesemedencébe.
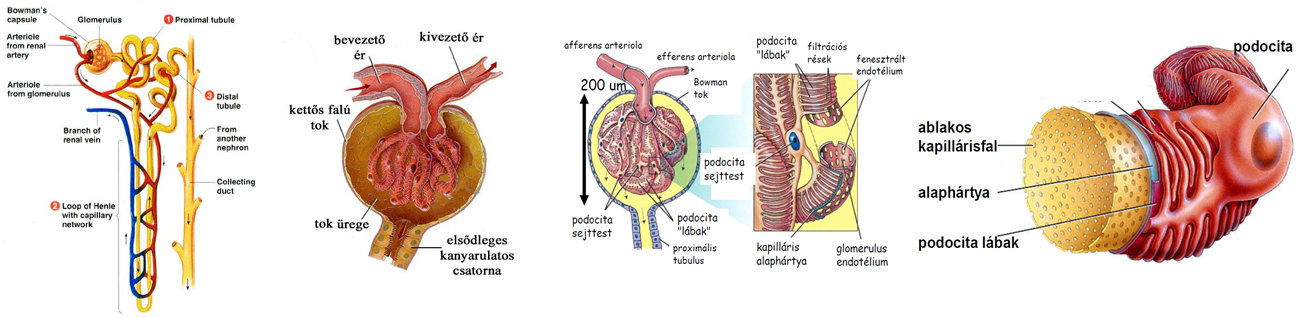 |
A vese működése
A vesék működésének a terméke a vizelet, amely
A szűrőműködés során csaknem fehérjementes vérplazma, a szűrlet jön létre. A szűrletben a fehérjéken kívül a vérplazma minden alkotója változatlan koncentrációban van jelen. Így tartalmaz
A szűrőberendezés ugyanakkor visszatartja a vér alakos elemeit és a plazmafehérjék nagy részét, pl. az albuminokat, globulinokat, fibrinogént, így azok sem a szűrletben, sem a vizeletben nem jelennek meg.
Ha a vizelet mennyiségét és összetételét összevetjük a szűrletével, jelentős különbségeket tapasztalunk. Ennek az az oka, hogy a szűrletből igen nagymértékű (99%) visszaszívás történik, mivel rengeteg olyan anyagot tartalmaz, amelyek a szervezet számára még szükségesek, pl. glükóz, aminosavak, sók és víz. A visszaszívó működések során a hasznos anyagok visszakerülnek a vérbe,
Az elvezető csatornák falának kiválasztó tevékenységének eredményeként a vizeletben megnő egyes anyagok koncentrációja a vérplazmához képest. A kiválasztást aktív transzportfolyamatok valósítják meg, ekkor az elvezető csatornát kísérő hajszálerekből olyan anyagok kerülnek a szűrletbe, amelyek
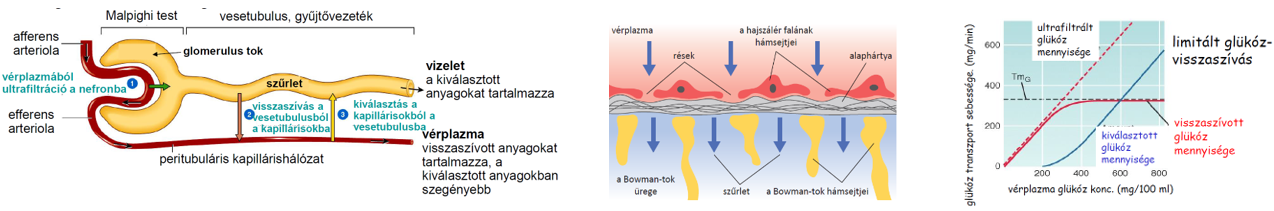 |
A szűrőműködés
A szűrőműködés során az érgomolyagból a vérplazma egy része, a vérnyomás hatására, a Bowman-tok üregébe jut. A folyamat eredménye az elsődleges szűrlet, amely gyakorlatilag fehérjementes vérplazma.
A szűrőberendezés az érgomolyagot és a Bowman-tok falát elválasztó negatív töltésű rostos alaphártya. A szűrő pórusai akkorák, hogy a vér alakos elemeit és a negatív töltésű nagy méretű plazmafehérjék zömét visszatartják. Tehát a szűrletben a vérplazma minden alkotója – a fehérjéken kívül – változatlan koncentrációban jelen van.
A szűrőműködés hatékonyságát növeli a hajszálérgomolyagban uralkodó nagy vérnyomás (kb. 50 Hgmm), ami annak köszönhető, hogy az érgomolyagból kivezető arteriola átmérője kisebb, mint az odavezető artelioláé.
A szűrőműködés az érgomolyag kapillárisaiban és a Bowman-tokban uralkodó nyomáskülönbségnek köszönhető, passzív folyamat, energiát nem igényel. A szűrés egymással ellentétes hatások eredőjeként valósul meg.
Tehát az eredő nettó filtrációs nyomás: Peff= Pv - Pcoll - PB
Ozmózisnyomásnak nevezzük azt a nyomást, amelyet a tiszta oldószerrel féligáteresztő hártyán át kapcsolatban lévő oldatra kell kifejteni ahhoz, hogy a dinamikus egyensúly beálljon. Az ozmózisnyomás az oldat koncentrációjával egyenesen arányos. Az ozmózisnyomás kialakításában az összes ozmotikusan aktív anyag részt vesz.
Az onkotikus (kolloidozmotikus) nyomás az ozmotikus nyomás azon része, amelyet a nem diffundáló kolloidok (főleg fehérjék, a vérben az albuminok) hoznak létre.
A Bowman-tokba jutó szűrlet (fehérjementes vérplazma) ozmotikus koncentrációja (ozmolalitása) lényegében megegyezik a vérplazmáéval, vagyis izoozmotikus. A vérplazma ozmotikus koncentrációjának döntő részét a kis ionok adják (elsősorban Na⁺ és a kísérő anionok), mivel a vérplazma fehérjekoncentrációja ezekhez képest csekély.
A Bowman-tok szűrlete tehát azért izoozmotikus a plazmához képest, mert az ozmolalitást nem annyira a fehérjék, hanem főleg a kis oldott részecskék adják.
Az oldatban lévő ozmotikusan aktív - amelyek nem jutnak szabadon át a féligáteresztő hártyán - oldott részecskék számát az ozmolalitással fejezzük ki. Minél nagyobb az ozmolalitás, annál nagyobb az ozmotikus nyomás. Az ozmózisnyomás kialakításában az összes ozmotikusan aktív anyag részt vesz.
A fehérjék nem a teljes ozmolalitást, hanem a kolloidozmotikus (onkotikus) nyomást határozzák meg.
Vérplazma teljes ozmolalitása: ≈ 285–295 mOsm/kg Ebből:
A glomeruláris kapillárisban a víz kilépése miatt nő a plazmafehérjék koncentrációja ezért folyamatosan nő a vér onkotikus nyomása (ozmózisnyomása):
Visszaszívó működés
A glükóz és az aminosavak visszaszívódása
Az átszűrődött glükóz
ezért egészséges, éhező ember vizeletében nem, vagy csak nyomokban mutatható ki cukor.
Amennyiben a vérplazmában – és a szűrletben – a glükóz koncentrációja meghaladja a 200 mg/100 ml-t, a cukor megjelenik a vizeletben, mivel az aktív transzporttal visszaszívást végző szállítómolekulák elérik kapacitásuk felső határát. A glükózhoz hasonlóképpen szívódnak vissza az aminosavak.
A sók és a víz visszaszívódása
A sók és a víz az elvezető csatorna csaknem teljes hosszában, ill. a gyűjtőcsövek területén is visszaszívódnak.
A proximális tubulus működése
A víz és a sók nagy része – kb. 70%-a – a proximális tubulusban szívódik vissza. A proximális tubulust körülvevő erekben található vér ozmotikus szívóereje jelentősen megnő, mivel a szűrőműködés során kilépett nagy mennyiségű víz miatt megnő a vérfehérjék ozmotikus koncentrációja, ami hajtóereje lesz a víz passzív mozgásának.
A proximális tubulus végén a szűrlet koncentrációja nagyjából megegyezik a vérplazma koncentrációjával, azaz izoozmotikus.
Bármely, a csatornában nem felszívódott ozmotikusan aktív anyag – pl. cukor – csökkenti a víz visszaszívódását, s emiatt a következő szakaszba érkező folyadék mennyisége megnő. A későbbi szakaszok nem képesek a megnövekedett vízmennyiséget visszaszívni, ezért a vizelet mennyisége megnövekszik. Ezt a jelenséget ozmotikus diuresisnek nevezzük, ezzel magyarázzuk a cukorbetegek nagyobb mértékű vizeletürítését.
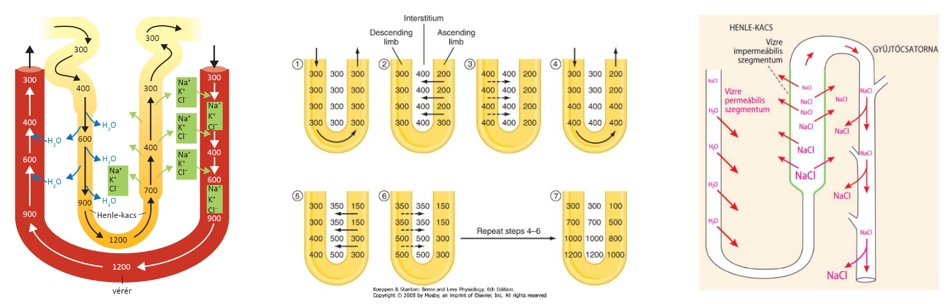 |
Henle-kacs működése
A Henle-kacs leszálló és felszálló ágában eltérő jellegű transzportfolyamatok működnek.
A leszálló ágban
A folyamatok eredményeként a leszálló ágban a szűrlet koncentrációja a vesepiramisok csúcsa felé egyre nő, a vérplazma koncentrációjához képest hiperozmotikussá válik. A víz passzív transzportját – ozmózist – az teszi lehetővé, hogy a leszálló ág csövei rendkívül nagy ozmotikus koncentrációjú szöveti környezetben haladnak.
A Henle-kacs felszálló ágának fala ugyanakkor
Mindezek miatt a felszálló ág folyadéka a kéregállomány felé egyre hígul, a disztális tubulusba érkezve hipoozmotikussá válik.
A felszálló ágból kiáramló sók tehát fokozzák a csövek körüli szövetnedv és az itt futó véredényekben a vérplazma koncentrációját, aminek köszönhetően a víz passzív transzporttal léphet ki a leszálló ágból és a gyűjtőcsatornákból, amelyek szintén ezen a területen keresztül futnak le. Az aktív transzportfolyamatoknak köszönhetően a vesepiramisok csúcsa felé egyre növekedő koncentrációgradiens jön létre.
A koncentrációgradiens kialakulásában elsődleges szerepet játszik a Henle-kacs felszálló ágában folyó NaCl-visszaszívás, amelyet nem kísér víz-visszaszívás. Ennek köszönhetően a szövetnedv NaCl-koncentrációja meghaladja a felszálló és a leszálló ág szűrletének koncentrációját. A szövetnedv NaCl-koncentrációjának további növelésére a leszálló és a felszálló ágak egymás melletti lefutásának köszönhetően kialakuló ellenáramú sokszorozás ad lehetőséget.
Ennek lényege, hogy
Az ellenáramú sokszorozást kiegészíti a csatornákat körülvevő, egymással és a csatornákkal párhuzamosan futó erek ellenáramú kicserélése. Az erekben áramló vér összetétele kiegyenlítődik a környezetével, a leszálló erekben áramló vér egyre hiperozmotikusabbá válik, a párhuzamosan felszálló erekben pedig átveszi a csatornákból kiáramló víz egy részét.
A disztális tubulus működése
A disztális tubulusban csak kismértékű só-(5%) (aldoszteron hatására) és jelentéktelen vízvisszaszívás történik. A folyamatok eredményeként a gyűjtőcsatorna felé a kezdeti szűrlet mennyiség 15%-a, kb. 27 dm3 folyadék halad.
A karbamid – urea – az aminosavak nitrogéntartalmú részének – aminocsoportjának – bomlásából jön létre a májban. Visszaszívódása passzív transzporttal történik. A proximális tubulusban kb. 50%-a szívódik vissza. A karbamid a gyűjtőcsövekből is jelentős mértékben lép ki, hozzájárulva a csatornák közötti tér növekvő koncentrációgradiensének kialakításához.
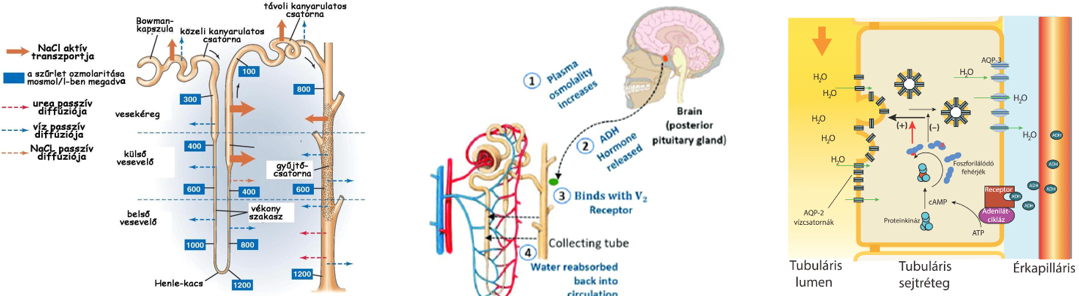 |
A kiválasztás szabályozása, a gyűjtőcsatornák működése
A gyűjtőcsatornák feladata a vesemedencében összegyűlő vizelet végleges mennyiségének kialakítása, összetételének és ozmotikus koncentrációjának a végső beállítása. A gyűjtőcsatornában a só és a víz visszaszívódásának a mértéke hormonálisan – ADH, aldoszteron – szabályozott a szervezet só és vízellátottságának megfelelően.
A gyűjtőcsövek a Henle-kacsokkal párhuzamosan futnak a vesepiramisok csúcsai felé, egyre nagyobb koncentrációjú szövetnedvvel körülvéve. A gyűjtőcsövek végére kialakul a végleges vizelet, amely az eredeti szűrletmennyiség kb. 1 - 0,5%-a, töménysége általában hiperozmotikus.
A gyűjtőcsövekben a víz visszaszívódását elősegítő hormon, az ADH (antidiuretikus hormon, vazopresszin) a hipotalamuszban termelődik és az agyalapi mirigy hátsó lebenyében raktározódik. Innen szabadul fel szükség esetén a hipotalamusz idegsejtjeinek hatására, melyek közvetve képesek érzékelni a vér koncentrációját (ozmózisnyomását).
Vízhiányos állapotban – amikor a vér ozmotikus koncentrációja (ozmózisnyomása) megnő –
A gyűjtőcsatornák falában az akvaporin-2 (AQP-2) típusú vizescsatornák teszik lehetővé a víz passzív transzportját, melyek beépülését a sejthártyába az ADH hormon szabályozza.
Az ADH a vízvisszaszívás fokozása révén a vérnyomást is emeli (vazopresszin). ADH hiányában nagy mennyiségű, híg vizelet keletkezik, a vérnyomás csökken.
A Na+ visszaszívódását – Na+-K+ csere révén – a disztális tubulusban és a gyűjtőcsövekben a mellékvesekéregben termelődő szteránvázas aldoszteron serkenti, amely sóhiányos állapotban termelődik fokozottabban. A sóvisszaszívódás növekedése esetén fokozódik a víz visszaszívása is, hiszen a víz passzív mozgását a növekvő koncentrációgradiens elősegíti, aminek köszönhetően emelkedik a vérnyomás.
Aldoszteron hiányában ezért nagyobb mennyiségű, koncentráltabb vizelet alakul ki, a vérnyomás pedig csökken. A hormon elválasztásának a mértékét – az ACTH mellett - döntően a vérnyomás határozza meg (renin-angiotenzin rendszeren keresztül), az alacsony vérnyomás fokozza a termelődését.
A vese szerepe a vér pH-jának szabályozásban
A vesékben a gyűjtőcsatornák sejtjei a vér savasodásakor szén-dioxidot vesznek fel. A sejtekben a szén-dioxid a vízzel szénsavat képez, ami bomlik. A H+- ionok a szűrletbe kiválasztódnak, s a vizelettel távoznak, míg a HCO–3-ionok visszakerülnek a vérbe.
A vizelet koncentrációja széles határok között változhat a körülményektől függően, napi mennyisége kb. 1,5 l, szalmasárga, átlátszó, pH-ja 6 körüli. Tartalmaz
A vizelet elvezetése
A vizelet a vesében a vesemedencében gyűlik össze, innen a húgyvezeték perisztaltikus mozgással továbbítja a húgyhólyagba, ahol időlegesen tárolódik, ill. szakaszosan ürül a húgycsövön keresztül.
A hólyag a falában található simaizomréteg összehúzódásával aktívan részt vesz a vizeletürítésben.
A húgycső a húgyhólyag alsó részéből indul ki, eredési helyén gyűrű alakú simaizom található, lejjebb pedig egy erős, harántcsíkolt záróizom, mellyel a vizeletürítés akaratlagosan szabályozható. A végbél záróizmához hasonlóan tehát a húgycsőnek is van egy simaizomból álló záróizma és egy harántcsíkolt záróizma. A húgycső a nőknél rövidebb, önállóan nyílik a külvilágba, férfiaknál hosszabb, a péniszben fut, ugyanis a vizeletelvezető és az ivarutak végső szakasza közös, a prosztatában a húgycső egyesül az ondóvezetékkel.
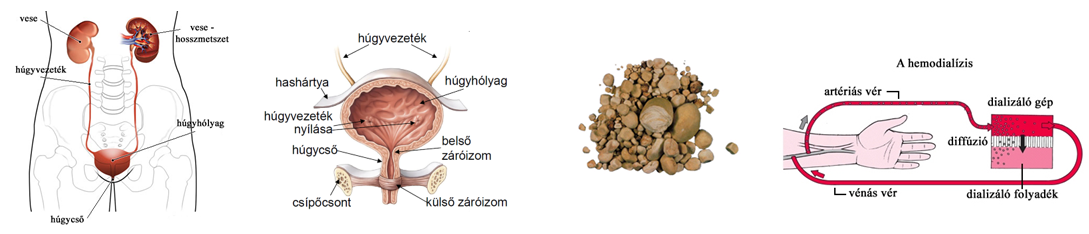 |
A kiválasztó szervrendszer egészségtana
Nemcsak a vizelet koncentrációja változhat széles határok között, hanem az összetételét és a mennyiségét egyaránt számos körülmény befolyásolja, mint pl.
A vizelet tulajdonságainak – mennyisége, színe, szaga, kémhatása, sűrűsége, kémiai összetétele – megváltozásából a kiválasztó-szervrendszer működésének zavarára és számos belgyógyászati problémára is fény derülhet. Ezért a vizelet vizsgálata számos betegség diagnózisához elengedhetetlen.
A vizelet mennyiségének kóros megnövekedése egyes hormonális rendellenességekre hívhatja fel a figyelmet.
Mindkét betegség esetén a betegek vízfogyasztása a vízvesztés következtében jelentősen megnő.
Egészséges éhező ember vizelete cukormentes, hiszen a szűrletbe került glükóz teljes mennyisége aktív transzporttal visszaszívódik, azonban cukorbetegség esetén a magas vércukorszint miatt a glükóz megjelenik a vizeletben.
Nagyobb mennyiségű fehérje megjelenése a vizeletben a vese, ill. a húgyutak gyulladásos, fertőzéses folyamataira utalhat, csakúgy, mint a fehérvérsejtek, ill. a vér jelenléte. Albuminokat akkor lehet kimutatni a vizeletből, ha a szűrőberendezés negatív töltése megszűnik, mivel ekkor már nem képes visszatartani a szintén negatív töltésű fehérjemolekulákat.
A véres vizelet továbbá jelezhet vese-, ill. húgyúti tumort és vesekövet, a hemoglobin megjelenése pedig hemolízisre utalhat.
Vesekő abban az esetben alakul ki, ha a vizelet kissé lúgosabb, ill. töményebb az optimálisnál, ekkor az egyébként oldott sók kikristályosodva apró szemű vesehomokot vagy egy nagyobb méretű vesekövet képeznek. A vesekő kialakulhat a vesemedencében vagy a húgyhólyagban egyaránt. Megfelelő mennyiségű folyadék fogyasztásával csökkenteni lehet a sók kiválásának a kockázatát.
Csökkent mennyiségű vizeletet eredményezhet a veseelégtelenség, a vese működésének súlyos zavara. Ebben az esetben a különféle toxikus anyagok nem tudnak távozni a szervezetből, s emiatt a vérben felhalmozódnak. A betegséget okozhatja a vese ereinek a szűkülete, károsodása – pl. cukorbetegség következtében –, ami miatt a vesén áthaladó vér mennyisége, ill. nyomása annyira lecsökken, hogy nem megfelelő a szűrőműködés hatékonysága. A vese szöveteinek a károsodása gyulladásos vagy mérgezéses okok miatt szintén veseelégtelenséghez vezethet.
A betegség kezelésére megoldás lehet a művesekezelés vagy a veseátültetés.
A művesekezelés – dialízis – az elégtelen veseműködések pótlására szolgáló eljárás. Többféle formája ismert. A hemodialízis dialízisközpontokban történik, hetente 3 x 4 óra időtartamban. A kezelés során a beteg vérét a dializáló gépbe vezetik, ahol egy féligáteresztő falú csőben áramlik egy dializáló folyadékkal körülvett térben, majd a berendezésből a vér salakanyagoktól megtisztulva kerül vissza a szervezetbe. Működési elve a diffúzió, a mérgező anyagok a koncentrációkülönbségnek megfelelően jutnak ki a vérből a féligáteresztő hártyán keresztül a csöveket körülvevő steril dializáló oldatba.
38. tétel: Kémiai érzékelés |
Szaglás és ízérzékelés
Közös jellemzők:
Szaglás
Szaglópálya
 |
Ízérzékelés
Nemcsak a nyelven, de a szájpadláson, a torokban, a gégefedőn és a nyelőcsőben is találhatók ízlelőbimbók és mindegyik terület érzékeli mindegyik ízt. A nyelven van némi különbség abban, hogy a nyelv különböző részein található ízlelőbimbók melyik alapízekre érzékenyebbek, de ez az eltérés lényegében elhanyagolható.
A receptorok 5-féle alapízre érzékenyek:
 |
Egyéb kemoreceptorok szerepe különféle élettani folyamatokban
A nyúltvelői szén-dioxidra (H+-ionra) érzékeny kemoreceptorok légzés szabályozásában betöltött szerepe
Az agytörzs területén – nyúltvelő és a híd – találhatók a légzés szabályozásáért felelős idegi központok. A nyúltvelőben megkülönböztetünk belégző és kilégző központot.
A légzés szabályozásában
A kémiai ingereket
A belégzésben a vér szén-dioxid koncentrációja, ill., ami ezzel arányos, az idegszövet H+-koncentrációja a legfontosabb inger, mivel ezek a belégző központ működését serkentik.
Az agykamrák falánál elhelyezkedő agyterületek Na+-koncentrációt érzékelő receptorai és ezek szerepe a szomjúság és a hipotalamikus vazopresszin (ADH) termelés folyamatában
Szomjazáskor, a szervezet vízhiánya esetén
A testfolyadékok koncentrációjának emelkedése – elsősorban a Na+-koncentráció - az a fiziológiás inger, ami felelős a szomjúságérzet kialakulásában ill., ami kiváltja a vízkereső magatartást.
A vér összetételének a megváltozását a hipotalamuszban, az agykamra falánál található neuronok kemoreceptorként képesek érzékelni. Ez a terület a szomjúságközpont, mely a szomjúságérzet és a vízkereső magatartás kialakításáért felelős, aktiválódásának ingere – a csökkent folyadékbevitel miatt – a vér ozmotikus koncentrációjának a megemelkedése.
Ugyanezen az idegrendszeri területen találhatók az ADH – vazopresszin – elválasztásáért felelős neuroszekréciós sejtek, melyek a vér Na+-koncentrációjának megnövekedésekor növelik hormon elválasztásukat. A fokozódó ADH termelés hatására nő a vízvisszaszívás mértéke a vesék gyűjtőcsatornáiban, csökken a vízleadás, aminek köszönhetően a szervezet igyekszik vízkészleteit megőrizni.
A vérkeringés – a vérnyomás és a szív működésének – szabályozásában szerepet játszó kémiai ingerek
A vérkeringés szabályozásában hormonális és idegi (vegetatív: szimpatikus, paraszimpatikus) szabályozó rendszerek vesznek részt.
A vérnyomás szabályozásában a különféle szabályozó mechanizmusok tulajdonképpen az erek szűkületét vagy tágulatát, így a vérnyomás emelését vagy csökkenését eredményezik.
A szívműködés – frekvencia, pulzustérfogat - szabályozása lényegében a szinuszcsomó működésére gyakorolt serkentő, ill. gátló hatások révén történik.
Az idegi szabályozóközpontok az idegrendszerben számos helyen megtalálhatók. Az elsődleges központok az agytörzsi hálózatos állomány nyúltvelői szakaszában helyezkednek el, működésüket kémiai és mechanikai ingerek befolyásolják. Itt kétféle, ellentétes hatású központ van.
A vérkeringés szabályozásában szerepet játszó kemoreceptorok
a) Perifériás:
b) Központi:
A táplálkozás szabályozásában szerepet játszó kemoreceptrorok
Az éhség egy motivációs állapot, a táplálék felkeresésére, a táplálék elfogyasztására irányuló belső késztetés. Kiváltásában többféle élettani tényező - pl. kémiai ingerek - játszik szerepet.
A táplálkozással kapcsolatos idegi szabályozó mechanizmusok központjai a hipotalamuszban vannak. Itt található
A fiziológiás éhség érzetét elsősorban az
A jóllakottság kémiai jelzései
A vér összetételének megváltozásait – vércukorszint, inzulinszint, ghrelinszint, leptinszint – a hipotalamuszban található szabályozóközpontok idegsejtjei kemoreceptorként érzékelik.
39. tétel: A látás |
 |
40. tétel: A hallás és egyensúlyérzékelés |
 |
41. tétel: A pajzsmirigy szerepe |
A pajzsmirigy elhelyezkedése
A pajzsmirigy a pajzsporc alatt és a légcső előtt található, két lebenyből áll.
A pajzsmirigy szerkezete
Mikroszkopikus szerkezetére jellemző, hogy működési állapotától függően különböző magasságú sejtek által határolt hólyagocskák építik fel. A hólyagok belsejét a sejtek által termelt hormonelőanyag tölti ki.
A hólyagok rendkívül gazdagon erezett lazarostos kötőszövetbe ágyazódnak. A hólyagok között a kötőszövetben kis sejtcsoportok találhatók, amelyek kalcitonint termelnek.
A pajzsmirigy egyik legfontosabb hormonja a tiroxin (T4), amely egy jódtartalmú aminosav-származék.
A T4 előállításához szükséges jódot az ivóvízből és a táplálékból vesszük fel. Ahol az ivóvíz nem tartalmaz elég jódot, a sót KI-dal egészítik ki.
 |
A tiroxin élettani hatásai
A tiroxinnak - a növekedési hormonhoz hasonlóan - nincs kitüntetett célszerve, hatása a legtöbb szövetre érvényesül. Hatásait alapvetően 2 területen fejti ki.
A tiroxin elengedhetetlen a normális egyedfejlődéshez, a sejtek differenciálódásához. A legnagyobb hatást a tiroxin a csontfejlődésre és az idegrendszer fejlődésére gyakorolja. A fehérjék szintézisét serkenti, így STH-al együttesen biztosítja a fejlődést, növekedést.
Általános hatása, hogy emeli az oxidatív lebontó-anyagcsere mértékét a mitokondriumok membránja áteresztőképességének növelésével.
Az oxidációs folyamatok serkentése miatt emeli a testhőmérsékletet.
A tiroxin elválasztását az agyalapi mirigy elülső lebenyének TSH-ja szabályozza. A szabályozás negatív visszacsatoláson alapszik.
A tiroxin valamennyi hatásának alapja a génátírás szabályozása, egyes gének átírását megindítja és fokozza, más gének átírását gátolja.
 |
A pajzsmirigy betegségei
A hipotireózis a pajzsmirigy csökkent működése, aminek oka általában jódhiány.
A) Felnőtteknél a hipotireózis a mixödéma nevű betegséget eredményezi. Ennek tünetei:
Hipotireózisban a tiroxin negatív visszacsatolása csökken, a TSH nagy koncentrációja miatt a pajzsmirigy kötőszövetes állománya megnagyobbodik, amit golyvának (strúmának) nevezzünk (hipofunkciós golyva).
B) Amennyiben a hipotireózis az embrionális fejlődés szakaszában lép fel, a kretenizmus nevű rendellenesség alakul ki. A betegek szellemileg súlyosan károsodottak, csontrendszerük aránytalan, a nemi érettséget gyakran nem érik el.
A hipertireózis a pajzsmirigy túlműködése, betegsége a Basedow-kor.
A megnövekedett tiroxinszint miatt a tünetei:
A fentieken túl jellegzetesen kidüllednek a szemek, és a pajzsmirigy fokozott működése miatt megnő a mirigyes állomány (hiperfunkciós golyva).
A Basedow-kor egy autoimmun betegség, mivel egy kórosan képződött immunoglobulin szerkezete a TSH-al megegyező, így a hormonhoz hasonló hatást gyakorol a pajzsmirigyre.
A pajzsmirigy állapotát ultrahanggal lehet vizsgálni, ennek során elsősorban azt figyelik meg, hogy a normálistól eltér-e a mérete, gyulladt-e, tartalmaz-e cisztát, daganatot.
42. tétel: A cukoranyagcsere szabályozása |
A cukoranyagcsere szabályozásában többféle hormon játszik szerepet. A különféle hormonok többnyire a máj szénhidrátforgalmát befolyásolják.
Növekedési hormon (STH, szomatotrop hormon, GH)
Növeli a vércukorszintet.
Tiroxin
Általános hatása, hogy emeli az oxidatív lebontó-anyagcsere mértékét a mitokondriumok membránja áteresztőképességének növelésével.
Kortizol
A kéregállomány középső rétegében elsősorban szénhidrát-anyagcserét befolyásoló hormonok termelődnek (glükokortikoidok), melyek közül a legfontosabb a kortizol. A kortizol elválasztását az ACTH szabályozza.
A kortizol az éhezéshez történő alkalmazkodásért felel:
Összességében az inzulin antagonistája.
Adrenalin
Az adrenalin kémiailag aminosav-származék, hatását tekintve ún. stresszhormon, nyugalomban a vérben szintje minimális, azonban megterhelés esetén a szervezet izgalmi állapotának a kialakításáért felelős.
Hatása:
Inzulin
Az inzulin egy 51 aminosavból álló polipeptid. Az inzulin az anyagcserét anabolikus irányba tolja el, a tápanyagok, testépítő vegyületek beépülését, raktározását serkenti.
A vércukorszint szabályozása
A máj szénhidrátforgalmát egyrészt idegi, másrészt hormonális tényezők befolyásolják.
A cukorbetegség (diabetes mellitus (mézédes húgyár))
Alapvetően kétféle cukorbetegséget szokás megkülönböztetni:
A diabetesz elsődleges tünetei
A tünetek hátterében a magas vércukorszint és az ennek ellenére fennálló sejtéhezés áll (mivel a sejtek inzulin hiányában nem képesek felvenni a glükózt (éhezés terített asztalnál)).
Összefoglalva:
Az inzulin elősegíti a glükóz felvételét, lebontását, zsírokká (máj, zsírszövetek) vagy glikogénné (máj, vázizom) alakulását.
Inzulin hiányában
Hosszútávon a cukorbetegségnek számos szövődménye lehet, aminek hátterében az áll, hogy a magas vércukorszint miatt a szervezet fehérjéihez a normálisnál sokkal több cukor kapcsolódik, ami megváltoztatja a szerkezetüket, rontja a működésüket.
Ezért jellemző:
Az 1-es típusú cukorbetegség mai tudásunk szerint nem előzhető meg, mivel kialakulásában örökletes tényezők döntőek.
A 2-es típusú cukorbetegség kialakulásában örökletes és életmódbeli tényezők is szerepet játszanak.
Életmódbeli tényezők:
Kezelés
Az 1-es típusú diabetes kezelése diétával és az inzulin bőr alatti zsírszövetbe juttatásával történik a végtagok, ill. a hasfal területén. Az inzulint be kell fecskendezni, szájon át jelenleg nem adható, mert peptid lévén a gyomorban kicsapódik, ill. lebomlik.
A 2-es típusú cukorbetegségben a betegség kezdetén ajánlott az életmód változtatás, szénhidrátszegény diétával, testsúlycsökkentéssel, sporttal. Ha ez nem elégséges gyógyszerek segítségével, amelyek csökkentik a vércukorszintet, növelik az inzulin hatását, fokozzák a sejtek inzulinérzékenységét. Ha a béta sejtek kimerültek, az inzulint teljesen pótolni kell.
43. tétel: A kalciumanyagcsere szabályozása |
A kalciumháztartás szabályozása
A kalciumion az egyik legjelentősebb szervetlen ion a szervezetünkben, mivel
A szervezetünk optimális kalcium ellátásában többféle szervünk is közreműködik:
A fenti szervek működésének összehangolását, a vér megfelelő kalciumszintjének kialakítását 3 hormon végzi,
E hormonok termelődésének mértékét a vér Ca2+-szintje határozza meg.
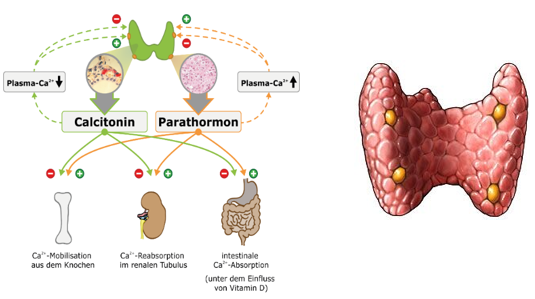 |
Mellékpajzsmirigyek
A pajzsmirigy csúcsán, annak állományába ágyazódnak be, számuk általában 4. A mellékpajzsmirigy a parathormont termeli, amely a vér Ca2+-szintjét emeli.
A parathormon:
A kalcitriol a D-vitamin aktív alakja. A kalcitriol a bélben növeli a kalcium-, valamint a foszfátfelszívódást, továbbá fokozza a csontok Ca2+ felvételét.
A pajzsmirigyben termelődő kalcitonin a vér Ca2+-szintjét csökkenti. Hatása nagyjából a parathormonéval ellentétes, főleg a csontrendszerre hat (fokozza a csontépítő sejtek működését).
Végül meg kell említenünk a nemi hormonok, az ösztrogén és a tesztoszteron a csontok összetételére gyakorolt hatását. Az ösztrogének csontvédő hatásúak, gátolják a fokozott csontbontást, segítik az építést. Szabályozzák a kalcium útját a tápláléktól a véren keresztül a csontokig és visszafelé, a csontokból a vérbe. Szabályozzák a kalcium kiürülését a veséken át. Az ösztrogén csontmegőrző hatásának pontos mechanizmusa nem tisztázott. Valószínűleg közvetlenül és közvetetten is szabályozza a csontanyagcserét, többek között úgy, hogy csökkenti a parathormon hatását, ugyanakkor növeli a kalcitonin aktivitását.
44. tétel: A gerincvelő működése |
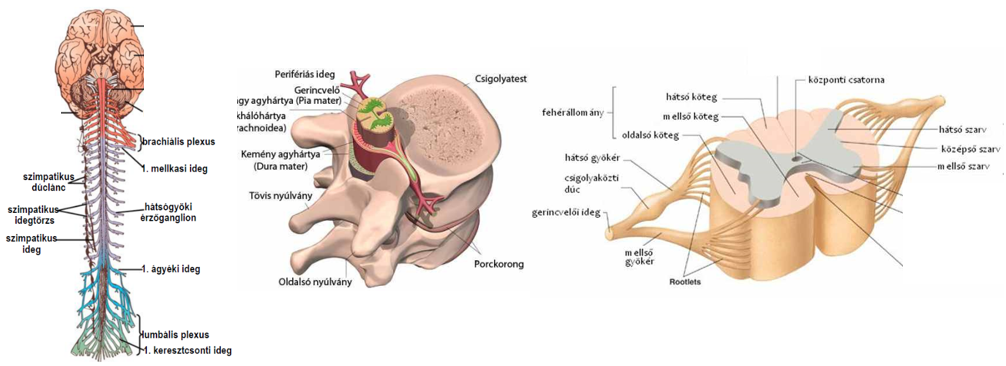 |
A gerincvelő keresztmetszete
Kétféle részt különböztetünk meg.
Szürkeállomány
Részei:
Fehérállomány
A gerincvelő szelvényezettségét a belőle kilépő, ill. belépő idegrostok, az ún. gerincvelői gyökerek okozzák.
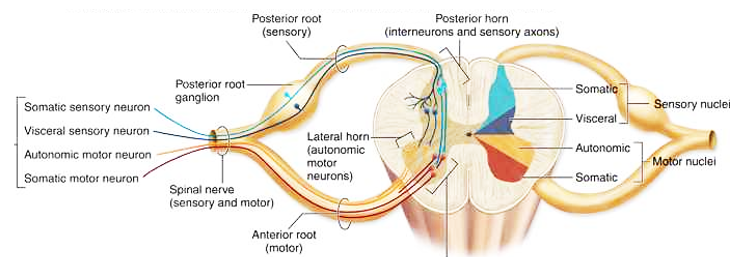 |
Mellső gyökér: idegrostjai a mellső szarvi mozgatóneuronokból és az oldalsó szarvi központi vegetatív neuronokból erednek, így kizárólag mozgatórostokattartalmaznak.
Hátsó gyökér: külső részén egy duzzanatot, az ún. csigolyaközti dúcot találjuk, benne pszeudounipoláris érzőidegsejteket, melyek centrális nyúlványai hozzák létre a hátsó gyökeret. Így a hátsó gyökérben kizárólag érzőrostok – szomatikus és vegetatív - futnak a receptorok felől a központok felé.
A két gyökér a gerinccsatornából való kilépés előtt gerincvelői ideggé egyesül, amely már kevert, érző- és mozgatórostokat egyaránt tartalmaz (31 pár). A gerincvelői idegek szelvényezetten hagyják el a gerincvelőt, s rostjaik szelvényesen hálózzák be a testet.
A szelvényezettség a dermatómák kialakulásában is megnyilvánul. A dermatóma a bőr, egy adott gerincvelői ideggel beidegzett területe. A neurológiában a gerinckárosodás területének a megállapítására szolgál. A dermatómák ismerete a gerincbe adott érzéstelenítés esetén is kiemelt fontosságú, hiszen az orvos a dermatóma-térkép alapján dönti el, melyik ideggyököt kell blokkolni a műtéti terület érzéstelenítéséhez.
A gerincvelő működése
A gerincvelő feladata kettős:
A gerincvelő működése és felépítése az evolúció során alig változott, csupán egyre többet vesztett önálló működéséből.
A gerincvelői reflexek típusai
Szomatikus: a végrehajtószerv valamilyen vázizom (ugyanakkor nem akaratlagos).
A receptor helye szerint:
Vegetatív: a végrehajtóegység zsigerekben található simaizom, ill. mirigyszövet.
Izomeredetű szomatikus reflex: feszítő reflex vagy nyújtási reflex (monoszinaptikus)
Ilyen a térd-, ill. a könyökreflex. Amennyiben reflexkalapáccsal a combfeszítő izom térdkalács alatti inára ütést mérünk, a lábszár kilendül. Az ütés következtében – mivel az inak nem nyújthatók - a combfeszítő izom megnyúlik, aminek hatására - a reflexkörnek köszönhetően - az izom aktív összehúzódással válaszol. A reflex vizsgálata tájékoztatást nyújt a gerincvelő állapotáról.
A receptor az izomorsó,
A reflexív lefutása
A bevezetőszárt az érzőidegsejt centrális nyúlványa képezi, mely belépve a gerincvelőbe 3 felé ágazik:
1. felszállópályán a hátsó kötegben az agyvelőbe tart, ill.
2. az azonos szelvényben elhelyezkedő mellső szarvi mozgatóneuronhoz kapcsolódik, melynek rostjai az elülső gyökéren keresztül érik el ugyanazt az izmot (combfeszítő izmot), kiváltva annak összehúzódását (a láb kilendül),
3. továbbá az alatta lévő gerincvelői szelvényben található hátsó szarvi gátló interneuronnal képez szinapszist, amely a hajlítóizom beidegzését végző mozgatóneuront gátolva, a hajlítóizom elernyedését váltja ki (elkerülendő az izomszakadást).
A nyújtási reflex feladata:
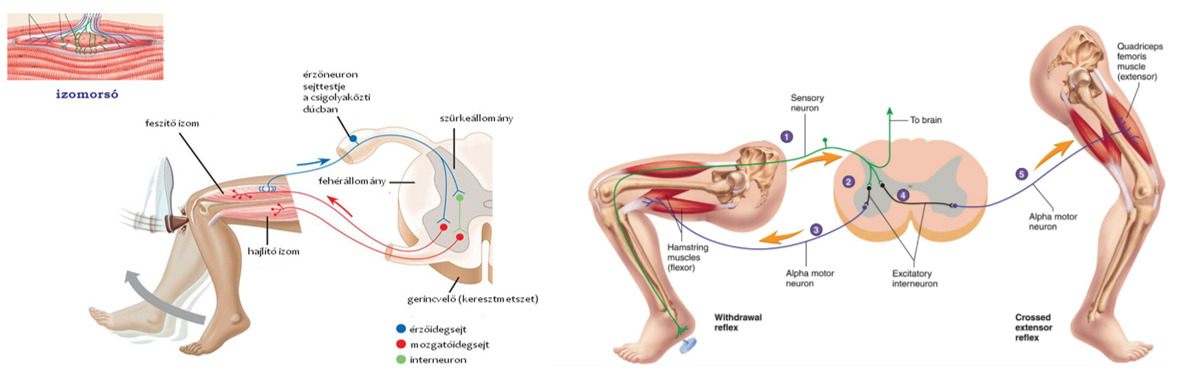 |
Bőreredetű szomatikus reflex, azonos oldali hajlító-keresztezett feszítő reflex
A reflex feladata a végtagok elhajlítása a fájdalmas ingerektől, a veszélyforrástól. Védekező reflex.
A reflexív lefutása
A reflexív működésének az eredménye a veszélyeztetett végtag hajlítóizmainak összehúzódása mellett, a másik oldali végtag feszítőizmainak összehúzódása.
A reflex feladata:
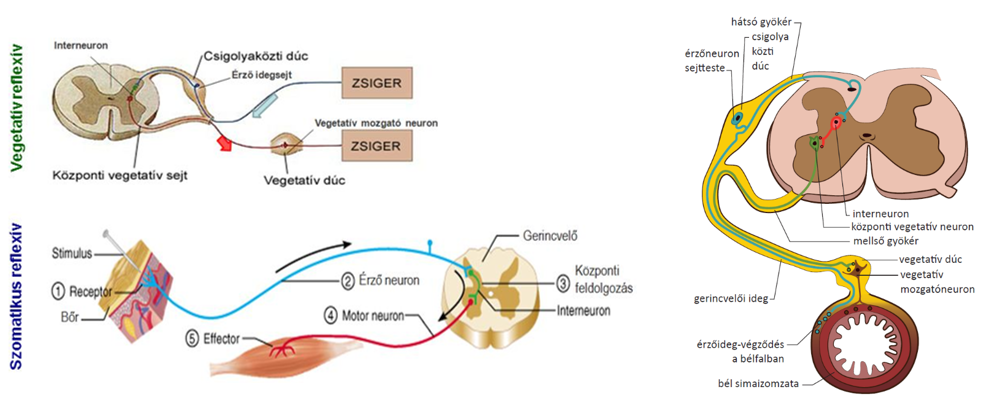 |
A vegetatív reflexek
Fontos különbség a szomatikus reflexhez képest, hogy a mozgatóneuron nem a gerincvelőben található, hanem a testben elhelyezkedő perifériális vegetatív dúcokban.
A reflexműködés a zsigerekben található simaizmokra, mirigyekre hat.
A reflexív lefutása:
A gerincvelő vegetatív működései (reflexei):
45. tétel: Szinapszis és membránpotenciálok |
Membránpotenciál
Minden élő sejtre jellemző, hogy a sejthártya két oldalán a különböző töltéssel rendelkező ionok eloszlása eltérő, aminek köszönhetően a sejten belüli tér (ICF) és a sejten kívüli tér (ECF) között potenciálkülönbség alakul ki, azaz feszültség mérhető.
Ezt a feszültséget – ami szinte minden élő sejtre jellemző - membránpotenciálnak nevezzük, értéke 20 és 100 mV közé esik.
A feszültséget a sejt belsejébe és a sejten kívüli térbe helyezett mikroelektródák között mérhetjük mV-ban.
Az idegsejtek nyugalmi állapotában (hatásoktól mentesen) a nyugalmi potenciál mérhető, melynek értéke átlagosan –70 mV (-50 és -90 mV között).
Az ICF-ban található elektród negatív az ECF-ban lévő elektródhoz képest, ezért a membránpotenciál értékét negatív előjellel vesszük (megállapodás alapján).
A sejten belüli tér jellemzői:
A sejten kívüli tér jellemzői:
A nyugalmi potenciál értéke az ingerlékeny sejtekben megváltozhat, ez különösen fontos idegsejtek és izomsejtek, érzékhámsejtek esetén.
A membránpotenciált az egyenlőtlen ioneloszlás hozza létre. Megváltozását az ionáramok megváltozása okozza. Mindkét folyamatot a membránban elhelyezkedő ioncsatornák és ionpumpák eredményezik.
Na+-K+ pumpa (ATP-áz, azaz ATP bontó enzim)
ATP-áz enzim, - amely minden állati sejtben megtalálható - aktív transzporttal 1 ATP hidrolízise mellett 3 Na+-t juttat ki és 2 K+-t visz be a sejtbe.
Nyugalomban a sejtek Na+ és K+ készletei állandóak (pl. nem fogynak el a Na+-ionok), mivel a sejthártyában található ún. szivárgási csatornákon keresztül a sejt folyamatosan K+-t veszít és Na+-t vesz fel diffúzióval (a Na+ diffúziója kismértékű).
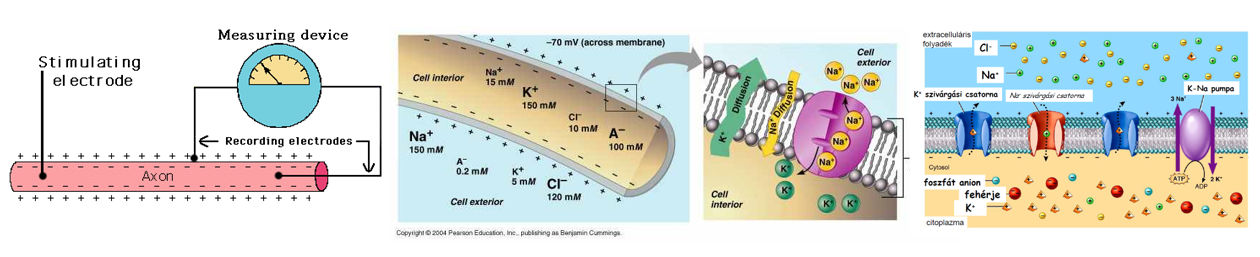 |
Ioncsatornák
Többféle passzív transzportot lebonyolító ioncsatorna létezik:
A nyugalmi potenciál megváltozhat:
Inger: külső, belső környezeti hatások, amelyek megváltoztatják egyes sejtek működését, anyagcseréjét, membránpotenciálját.
Ingerület: inger hatására bekövetkező anyagcsere-változás (membránpotenciál-változás).
Ingerküszöb: az a membránpotenciál-érték, amely kiváltja az akciós potenciált.
A különböző környezeti és pszichológiai hatások módosíthatják az ingerküszöböt.
Receptor: olyan sejt vagy idegvégződés, mely különféle ingereket képes felfogni azáltal, hogy a környezeti hatások ingerületi folyamatokat, potenciálváltozást eredményeznek működésében.
Adekvát inger az az inger, mellyel szemben a receptornak legkisebb az ingerküszöbe, ennek megfelelően megkülönböztetünk mechano-, foto-, termo-, kemoreceptorokat.
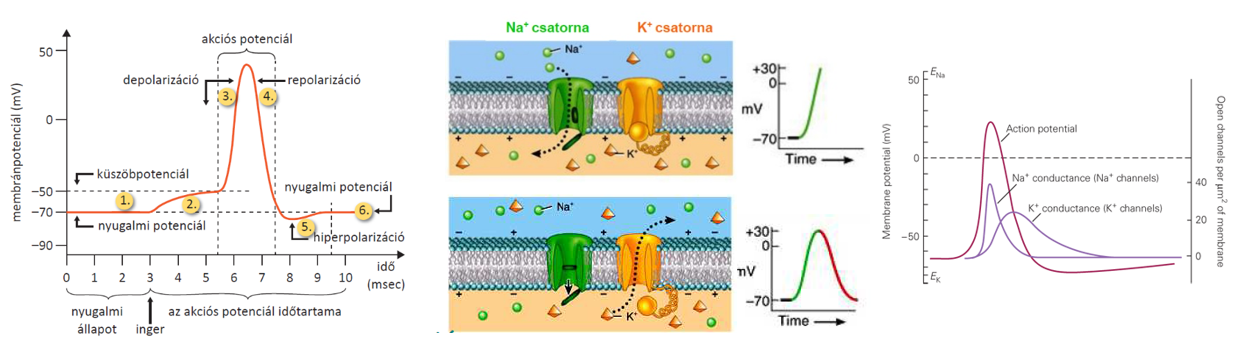 |
Akciós potenciál
Idegsejtek, izomrostok, izomsejtek, érzékhámsejtek inger hatására ingerületi állapotba kerülhetnek, azaz képesek a nyugalmi potenciáljuk gyors megváltoztatására és helyreállítására. A folyamat néhány ms alatt lejátszódik.
A nyugalmi potenciált (1) követő szakaszok:
Az akciós potenciál szakaszai:
Utópotenciál:
A folyamatokat leíró görbe az oszcilloszkóp képernyőjén tüske alakú.
Az akciós potenciál lezajlása
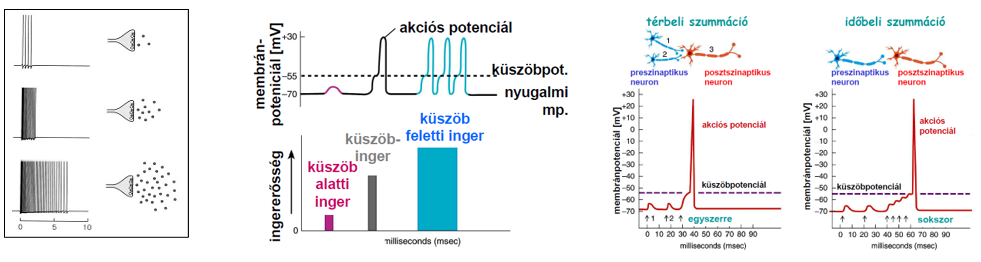 |
A posztszinapszison kialakuló helyi potenciálváltozás arányos az ürült neurotranszmitterek mennyiségével, így
A jelenség hátterében az áll, hogy a sejttesten és a dendriteken ligandfüggő, míg az axoneredésen és az axonon feszültségfüggő ioncsatornák találhatók.
Az inger erősségét az idegsejt a csúcspotenciálok sorozatának szaporaságában/frekvenciájában kódolja, erősebb ingerek nagyobb frekvenciájú akciós potenciálokat eredményeznek.
A preszinapszisból ürülő átvivőanyagok mennyisége arányos a preszinapszis membránján terjedő akciós potenciálok frekvenciájával.
Tehát, csúcspotenciál(ok) akkor alakulhat(nak) ki az axoneredésen, ha a helyi potenciálok eredője – térben ill. időben összegződve - az axondombon meghalad egy küszöbértéket, az ingerküszöböt.
A serkentő szinapszisok ún. lassú Na+-csatornákat nyitnak meg, amelyek helyi hipopolarizációt okoznak, ezek egyenként sosem érik el az ingerküszöböt, azonban hatásuk összegződhet és kiválthatnak akciós potenciált (gyors, feszültségfüggő csatornák megnyitásával az axondombon).
A gátló szinapszisok Cl--csatornák, ill. K+-csatornák megnyitása révén hiperpolarizálják a membránt.
Minden vagy semmi törvénye
Amennyiben egy inger erőssége eléri az ingerküszöb értékét, a kialakuló akciós potenciál értéke maximális, amplitúdója tovább nem fokozható (minden). Az erősebb ingerek nem az akciós potenciál amplitúdóját, hanem a sűrűségét, frekvenciáját növelik (frekvenciakód).
Azonban, ha a helyi potenciálok nagysága nem éri el az ingerküszöb értéket, nem alakul ki akciós potenciál (semmi).
Két fő jeltípust különböztetünk meg: analóg és digitális jelet.
A helyi potenciálok mértéke a belépő Na+ mennyiségétől függ, azaz mennyi ligandfüggő Na+-csatorna nyílik meg a szinapszisokban, amit viszont a felszabaduló neurotranszmitterek mennyisége határoz meg (analóg jel).
Az akciós potenciál lezajlásában az axonon feszültségfüggő csatornák vesznek rész, melyek ionáteresztőképességét már a sejt elektrokémiai tulajdonságai (kémiai és elektromos potenciál) határozzák meg, az áramló ionok koncentrációviszonyai egy adott értékig változtathatók, ami adott mértékben határozza meg a membránpotenciál nagyságát (digitális jel).
Ingerületvezetés
Az ingerület – potenciálváltozás – a sejttesten keletkezési helyétől minden irányba szétterjed a membránon, ugyanakkor az axonon visszafelé nem terjedhet. Sebessége néhány cm/s - 120 m/s-ig változhat.
A depolarizáció hatására a membrán síkjában is elmozdulnak az ionok.
Így az ingerelt hely repolarizálódik, a környező területek hipopolarizálódnak, amely elegendő az ingerküszöb eléréséhez, így ott is megnyílnak a feszültségfüggő csatornák, kialakul az akciós potenciál (köráram-teória).
Az idegrostok esetén a depolarizáció csak a gliasejtek közötti ún. befűződésekben jöhet létre, így az ingerület az idegroston ugrálva terjed, ami sokkal gyorsabb ingerületvezetés tesz lehetővé.
Gerinctelenekben nincs velőshüvely borítás, az akciós potenciál pontról pontra lassabban terjed, ezért ott az axon vastagságának növelésével fokozzák a vezetés sebességét. Ilyen pl. az alaposan tanulmányozott óriásaxon a tintahalakban. Tehát minél nagyobb az idegrost átmérője, annál gyorsabb a vezetés, mivel az axon vastagságának növekedésével nő annak felülete, ill. a felületen található ioncsatornák mennyisége, ill. minél nagyobb az axon átmérője, annál kisebb a sejten belüli vezető közeg ellenállása.
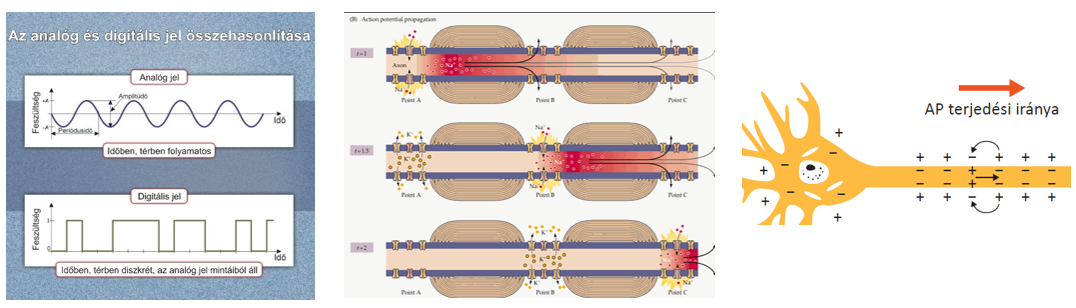 |
A szinapszis
Szinapszisoknak nevezzük az idegsejtek kapcsolódásait más idegsejtekkel, izom-, ill. mirigysejtekkel, a neuro-kommunikáció helyei.
Ismertek kémiai és elektromos szinapszisok.
Elektromos szinapszis (rés kapcsolat)
A kapcsolódó sejtek membránja között fehérjék alkotta csatornákon keresztül valósul meg az ingerületátadás egyik sejtről a másikra. Jellemzői:
Leggyakoribb előfordulási helyek:
 |
A kémiai szinapszis
Egy idegsejt
Az idegsejteket egyszerre többféle, egymással ellenkező hatás – serkentő és gátló – érheti. A helyi potenciálok összegződnek, eredőjük határozza meg a fogadó sejt membránpotenciál értékét.
 |
Az idegsejteket egyszerre többféle, egymással ellenkező hatás – serkentő és gátló – érheti. A helyi potenciálok összegződnek, eredőjük határozza meg a fogadó sejt membránpotenciál értékét.
Neurotranszmitterek lehetnek aminosavak, aminosav származékok, peptidek:
Egy neuron csak egyféle neurotranszmittert képes termelni (Dale-elv).
A kiürült átvivőanyagok hatásukat rövid ideig fejtik ki, mivel
A drog általában olyan hatóanyag, ami megváltoztatja a szervezet működését.
A tudatmódosító vagy pszichoaktív szerek azok a természetes vagy mesterséges drogok, amelyek a központi idegrendszer működését változtatják meg (lásd részletesen még 4.8.2. fejezet).
A drogok ill. egyes mérgek befolyásolják – serkentik vagy gátolják - a szinapszisok működését.
Hatásukat kifejthetik
1. a jelátvivő anyag
Tehát a gyorsító – serkentő - drogok
A lassító – depresszáns - drogok, nyugtatók
2. Receptormódosítás - a receptorokra ható agonista-antagonista hatás változása - révén.
Agonistának nevezzük azokat a hatóanyagokat, ún. ligandumokat, amelyek receptorokhoz képesek kötődni aktiválva működésüket. Ezek lehetnek
Tehát a drogok egyes fajtái is az idegsejtek receptoraihoz kötődve fejtik ki a hatásukat.
Antagonisták azok a vegyületek, amelyek valamilyen módon gátolják az agonisták hatását. A receptorokhoz kötődve inaktiválhatják azokat, ezáltal gátló hatást válthatnak ki.
A rendszeres droghasználat során függőség – addikció - alakul ki, azaz a szer csökkent bevitele vagy abszolút hiánya megvonásos tünetekkel jár, mivel a negatív visszacsatolás miatt a droghoz hasonló hatású, ugyanahhoz a receptorhoz kötődő, a központi idegrendszerben termelődő természetes transzmitterek termelődése csökken.
További következménye a különféle idegrendszerre ható anyagok – drogok - folyamatos alkalmazásának a tolerancia kialakulása, melynek során a szer tartós használata miatt annak hatása egyre csökken, s az anyag ugyan azt a hatást csak egyre nagyobb dózisban képes kifejteni. A tolerancia hátterében egyrészt a drogot lebontó enzimek aktivitásának növekedése, másrészt a receptorok számának a csökkenése áll.
46. tétel: A vegetatív idegrendszer |
Vegetatív érző- és mozgatórendszerek
Az idegrendszer funkcionálisan (a beidegzett területek alapján) felosztható
A vegetatív idegrendszer az idegrendszer azon működő része, amely az akaratunktól független szabályozza (autonóm módon), összehangolja a belső szervek működését. Szerepe az állandóan változó körülmények között a szervezet homeosztázisának fenntartása. A belső szervek reflexműködésén alapuló irányító rendszer (vegetatív reflexek).
Feladatai:
A vegetatív idegrendszer mind anatómiai, mind funkcionális értelemben két részre tagolható, szimpatikus és paraszimpatikus részekre. A szimpatikus és a paraszimpatikus rész ellentétes hatásokat vált ki a legtöbb szerven, ezért a két részt élettanilag antagonistáknak tekintik. Ugyanakkor mindkét rész egymással összehangoltan működik és az aktivitásuk közötti egyensúly az, ami fenntartja a stabil belső környezetet.
Vegetatív központok
Emeletszerűen szerveződnek, azaz a gerincvelői és a nyúltagyi elsődleges központokat a hídi és a középagyi másodrendű központok szabályozzák. Ezen felül az egész vegetatív rendszer működését hipotalamikus, ill. nagyagyi – limbikus rendszer - központok hangolják össze.
A gerincvelő oldalsó szarvában elhelyezkedő központi vegetatív neuronok irányítják a
Az agytörzsi hálózatos állományban
Hipotalamusz
A nagyagy integráló szerv, központjai főleg a limbikus rendszer területén találhatók.
Környéki része
Idegek
Mozgatóneuronokat tartalmazó dúcok
Receptorok
Fajtái:
Végrehajtók, célszervek a bőr, a belső szervek simaizmai, ill. mirigyei és a szívizom.
 |
Gerincvelő vegetatív reflexei
Gerincvelő által végrehajtott reflexek, melyek felső szabályozója a hipotalamusz. Ilyen például
Bevezető szár
Ugyanolyan, mint a szomatikus reflexek esetén, a csigolyaközti dúcokban található érzőidegsejtek nyúlványai vezetik be az ingerületet a hátsó gyökéren keresztül.
Központ
A gerincvelő oldalsó szarvában.
Kilépő vegetatív rostok
Szimpatikus idegrendszer
Általában a szervezet
A stresszelmélet szerint a stresszhatások (erőfeszítések, megpróbáltatások, fenyegető környezet, bekövetkezett ártalom, vagy nagy fizikai erőkifejtés) olyan fiziológiás változásokat okoznak a szervezetben, amelyek mindig azonos jellegűek, függetlenül attól, hogy mi volt a kiváltó ok. A különböző megterhelő ingerekre a szervezet azonos módon reagál, ún. szimpatikus túlsúly alakul ki, mely tónusos jellegű, az egész szervezetre kiterjed. E megváltozott fiziológiás állapotot Cannon- féle vészreakciónak nevezzük.
Cannon-féle vészreakció során:
A szimpatikus hatást közvetítő idegrostok a gerincvelő háti-ágyéki szakaszán lépnek ki a központi idegrendszerből.
Szimpatikus vegetatív mozgatódúcok a gerinc két oldalán találhatók (paravertebrális dúclánc).
Paraszimpatikus idegrendszer
A szervezet pihenését, regenerációját, energiakészletének feltöltését segíti (visszaállítás). Hatása kevésbé tónusos, mint a szimpatikus idegrendszeré, inkább helyi jellegű.
Hatásában közreműködő idegrostok
A paraszimpatikus vegetatív dúcok többnyire a beidegzett szerv falában találhatók.
Paraszimpatikus funkciók:
Belső szerveink kettős beidegzésűek, vagyis szimpatikus és paraszimpatikus rostokat egyaránt kapnak, kivéve a mellékvesevelőt, amely kizárólag a szimpatikus idegrendszerhez tartozik, mivel tulajdonképpen nem más, mint módosult szimpatikus dúc, benne átalakult, nyúlványaikat vesztett, hormonok – pl. adrenalin - termelésére specializálódott idegsejtek találhatók.
A kétféle hatás mértékét a hipotalamuszban található magvak hangolják össze, kialakítva az adott helyzethez szükséges optimális arányt.
47. tétel: Stressz és a hormonok |
A stressz
A stressz folyamatos feszültség, mely rendszerint negatív ingerekre adott tartós válaszreakció a szervezet részéről. A tartósan fennálló stressz akár komoly egészségkárosodást eredményezhet, mivel gyengíti a szervezet ellenállóképességét, folyamatos megterhelést jelent, stresszbetegségek – főleg szív és érrendszeri, ill. emésztőrendszeri - kialakulását eredményezheti.
A szervezetünk működésének a megváltozásáért részben
Kiváltó okok
Típusai
Tünetek megnyilvánulásának hátterében, részben fiziológiás, részben érzelmi történések állnak. A stressz általános tünetei:
A stresszortól függetlenül testünk automatikusan felkészül a veszedelemre. A stressz okozta tünetek hasonlóak a Cannon-féle vészreakcióhoz.
A fenti okok miatt fontos a stresszoldás lehetőségeinek ismerete, ilyen pl. aktív testmozgás, sportolás, relaxáció, minőségi alvás, pozitív társaskapcsolatok működtetése, pszichoterápiás kezelések igénybevétele.
Stresszhormonok
Adrenalin
A mellékvese velőállományának sejtjei átalakult idegsejtek, melyek nagymértékű hormonelválasztásra specializálódtak. A nyúlványaikat vesztett legömbölyödött sejtek kétféle hormont termelnek:
E hormonok elválasztása különféle stresszhatások, pl. erőfeszítések, megpróbáltatások, megterhelések (hideg, támadás, vérvesztés, jelentősebb fizikai munka stb.) hatására fokozódik.
Az adrenalin kémiailag aminosav-származék, hatását tekintve ún. stresszhormon, nyugalomban a vérben szintje minimális, azonban megterhelés esetén a szervezet izgalmi állapotának a kialakításáért felelős.
Hatása:
Az adrenalin a különféle szervekre – vázizomzat, bélcső - eltérő hatást gyakorol. A jelenség hátterében az áll, hogy a hormonhatást erőteljesen meghatározza a sejtek felületén elhelyezkedő adrenalinreceptor típusa.
A mellékvese velőállományát az idegrendszer közvetlen idegi úton szabályozza.
Kortizol
A kéregállomány középső rétegében elsősorban szénhidrát-anyagcserét befolyásoló hormonok termelődnek (glükokortikoidok), melyek közül legfontosabb a kortizol. A kortizol elválasztását az ACTH szabályozza.
A kortizol leginkább az éhezéshez történő alkalmazkodásért felel:
Az adrenalin az akut stressz esetén, a kortizol inkább a krónikus, hosszantartó stressz esetén választódik el nagyobb mértékben.
48. tétel: Az immunrendszer, immunválasz típusok |
A fehérvérsejtek
A fehérvérsejtek - szemben a vér alakos elemeinek másik két csoportjával - sejtmaggal rendelkező, teljes értékű sejtek. Számuk a szervezet állapotától függően széles határok között változhat, általában 4000 - 10 000/mm3, fertőzés, gyulladás esetén számuk jelentősen emelkedhet. A fehérvérsejtek az immunitásban játszanak szerepet, felépítésük és további szerepük alapján a sejteket 3 csoportba osztjuk.
A granulociták
Nevüket a sejtplazmában megtalálható nagy mennyiségű szemcséről (granulum) kapták, melyek membránkötött enzimek, és főként a bekebelezett részecskék lebontásában vesznek részt. A granulociták amőboid mozgásra képesek, átlépnek a hajszálerek falán, a szövetek közötti határfelületeken és a fertőzés irányába vándorolnak kemotaxissal. Az ún. nem specifikus (veleszületett) immunválaszban vesznek részt. A kórokozókat bekebelezik – kis falósejtek - és a sejtekben levő nagyszámú szemcse segítségével elpusztítják. Amikor a granulociták a kórokozókkal kapcsolatba kerülnek, belőlük is sok elpusztul. A fertőzött sebek körül keletkező genny az ilyenkor elhalt nagy tömegű granulocitától, kórokozótól és szétesett szövetelemektől származik. Viszonylag kis méretük és fagocitáló képességük miatt mikrofágoknak is nevezzük őket. Csupán néhány napig élnek. Gyulladásban fontos szerepet töltenek be.
A granulociták minden típusa a vörös csontvelőben termelődik.
A limfociták (nyiroksejtek)
Elnevezésük onnan származik, hogy a sejtek képződése részben vagy egészben, ill. elhelyezkedésük a nyirokrendszerhez kapcsolható (nyirok = lympha). Élettartalmuk néhány héttől több évig is terjedhet (memóriasejtek).
A sejteknek érési helyük és funkciójuk alapján két nagy csoportjuk van.
1. Természetes ölősejtek (NK), melyek a vírussal fertőzött sejtek, ill. a tumorsejtek elpusztításában vesznek részt.
2. Kis limfociták, melyeknek további 2 típusa ismert:
A vörös csontvelő és a tímusz ún. elsődleges nyirokszervek, az itt megért – antigén felismerésére képes - limfociták a vér és a nyirokkeringés segítségével a másodlagos nyirokszervekbe kerülnek, melyek a lép, a nyirokcsomók, a mandulák és a szervezetben elszórt nyiroktüszők.
A másodlagos nyirokszervekben az érett – antigén felismerésére képes - nyiroksejtek készenlétben állnak és kapcsolatba lépnek a kórokozókkal, miáltal aktiválódnak, megindítják az immunválaszt.
A monociták – makrofágok – antigénbemutató sejtek (APC)
A monociták nagyméretű, fagocitáló, vándorló sejtek, azonban ezeket a képességüket nem a vérben fejtik ki, hanem átlépve a hajszálerek falán a szövetekben makrofágokká alakulva végzik működésüket. Szintén a vörös csontvelőben jönnek létre.
Funkciójuk alapján hívjuk őket antigénbemutató sejteknek is, mivel bizonyos T-limfociták számára ők teszik lehetővé az idegen anyagok (immunogének) felismerését.
A makrofágok alapvetően fontos szerepet játszanak az immunválasz minden fázisában.
A granulocitákat és a makrofágokat szokták falósejteknek is nevezni.
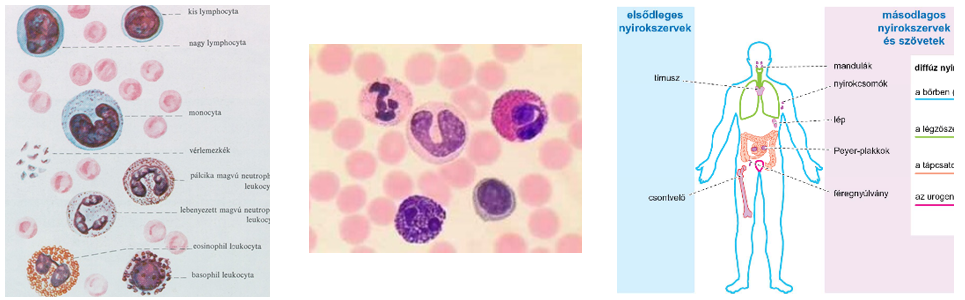 |
A szervezet egységes működésének feltétele többek között az is, hogy
Ezt a fontos feladatot rendkívül sokféle típusú sejt bonyolult együttműködése révén létrejövő biológiai védekezőrendszer, az immunrendszer látja el. A külső környezetből származó veszélyforrásokkal – paraziták, kórokozók - szemben a biológiai védelem mellett léteznek fizikai akadályok és kémiai védelmi mechanizmusok.
Fizikai védelem
Kémiai védelem
Szervezetünk kémiai védelmét
Biológiai védelem
A biológiai védelmet
Mikrobiom
Az emberi szervezetben élő, sajátos ökológiai rendszert alkotó mikrobák összességét, genetikai állományukat, kölcsönhatásaikat mikrobiomnak nevezzük. Ide tartoznak a bélrendszerünkben és különféle testfelületeinken megtalálható szimbióta, kommenzalista, parazita baktériumok, gombák, vírusok, egyéb mikroorganizmusok, mint pl. egysejtűek. A mikrobiom optimális összetétele nélkülözhetetlen az emberi szervezet egészséges működése szempontjából. A mikrobiom alkotói
A mikrobiom egyensúlyának megváltozása bizonyos betegségek kialakulását eredményezheti, mint pl. bélproblémák, elhízás, cukorbetegség, bőrproblémák, szorongás, depresszió. A Crohn-betegséget (gyulladásos bélbetegség), mely ma még ismeretlen eredetű autoimmun betegség, összefüggésbe hozzák a bélflóra összetételének kóros megváltozásával.
Immunrendszer
Az immunrendszer működésének az alapja, hogy a kivitelezésben résztvevő immunsejtek tudják megkülönböztetni a szervezet saját anyagait a testidegen anyagoktól. A sejtek felismerése a sejtköpenyben levő fehérje-szénhidrátláncok – jelölő fehérjék, markerek - alapján történik, melyek az adott szervezetre specifikusak, genetikailag meghatározottak.
Azokat az anyagokat, melyeket az immunrendszer sejtjei receptoraik segítségével felismernek, összefoglalóan antigéneknek nevezzük, makromolekulák, leggyakrabban fehérjék, poliszacharidok, nukleinsavak.
Tehát a már megismert anyagokat – tolerogének - az immunrendszer eltűri, tolerálja, azonban azokkal az objektumokkal – immunogének - szemben, amelyeket az immunrendszer nem ismer fel sajátként, megindul az immunválasz. Amennyiben az immunogén
Az autoantigénnel szemben ún. autoimmunfolyamat indul be, amely kedvező, ha a saját sejtek valamilyen oknál fogva rendellenesen jönnek létre, pl. daganatos sejtek esetén.
Abban az esetben, ha az immunreakció normális anyagokkal szemben indul meg, a folyamat kóros és különböző autoimmun-betegségeket eredményez, mint pl. az I. típusú cukorbetegség, a sclerozis multiplex, gluténérzékenység, a Basedow-kór, egyes ízületi gyulladások, vérlemezke hiányos vérzékenység stb.
A heteroantigéneket a következőképpen lehet csoportosítani.
Immunválasz típusok
Az immunválasz lehet:
Mindkét esetben lehet:
A nem specifikus – veleszületett - immunválasz
A természetes, avagy veleszületett immunitás elemei már jóval a születést megelőzően megjelennek a szervezetben, gyors, az antigén fajtájától független (nem antigén specifikus), általános védekezést nyújt a kórokozókkal és más káros anyagokkal szemben. Folyamatosan jelenlévő elemekből áll, ezért megindulása azonnali, ugyanakkor nem alakít ki immunmemóriát.
A nem specifikus humorális immunválaszt különféle vegyületek alakítják ki.
Komplementrendszer
Ezek a vérplazmában keringő inaktív fehérjék, enzimek (30-féle), melyek láncreakcióban aktiválják egymást.
Az interferonokat a vírussal fertőzött sejtek bocsátják ki. A még egészséges sejtek megkötik, bennük olyan enzimek képződését váltják ki, amelyek akkor keletkeznek, mikor bekövetkezik a fertőzés. Ezek az enzimek pl. bontják a vírus mRNS-t, gátolják a fehérjeszintézist stb.
A nem specifikus sejtes védekezésben a szervezet
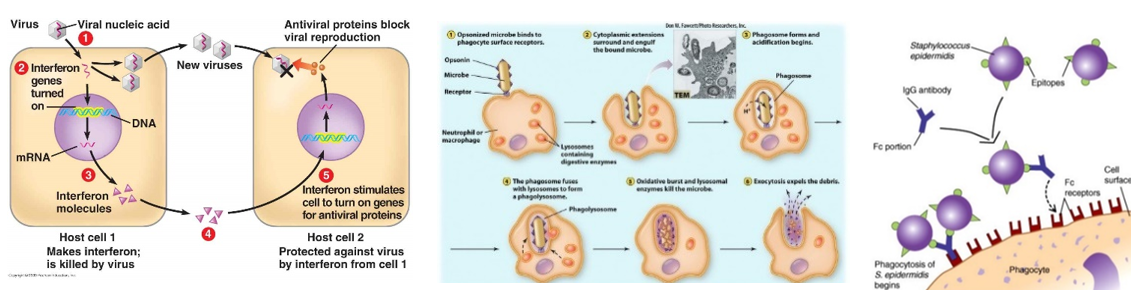 |
Hogyan semmisítik meg az immunogéneket a veleszületett immunválasz sejtjei?
A falósejtek bekebelezik az antigéneket, majd sejten belül elpusztítják őket.
A természetes ölősejtek
Hogyan ismerik fel a természetes immunválasz sejtjei az immunogéneket?
A veleszületett immunrendszer sejtjei
A vérben keringő és a szövetekbe vándorló természetes ölősejtek (NK) a szervezet egészséges sejtjeinek felszínén meglévő MHC-I-et ismerik fel. Az MHC-I felismerése egy tiltó jelet vált ki az NK-sejtekből, megakadályozva a saját sejtek megölését, ezért a saját MHC-I hiánya, módosulása az NK-sejtek ölő- (citotoxikus) mechanizmusát indítja be. A sejtek daganatos elfajulása az MHC-I hiányát okozza, ami főleg az NK-sejtek révén kiküszöbölésüket jelenti.
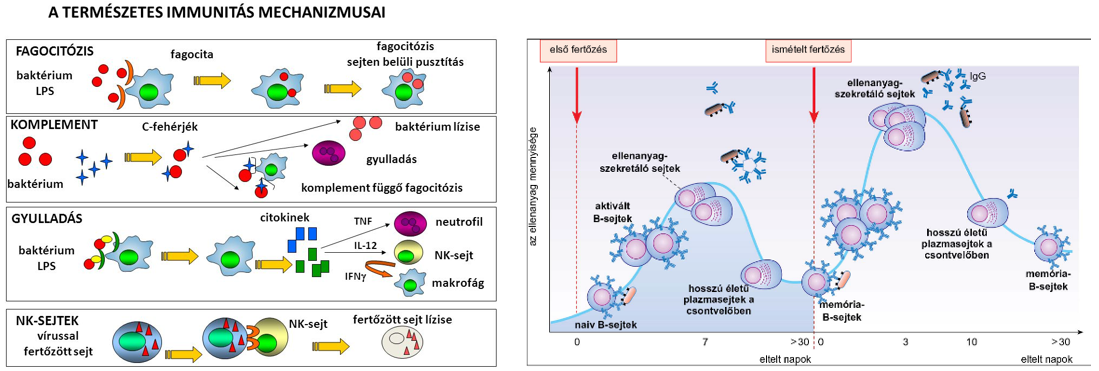 |
A specifikus vagy adaptív vagy szerzett immunválasz
A specifikus immunválasz adott típusú antigénre irányul, antigénenként különféle módon. Az adaptív immunitás az újszülöttekben gyakorlatilag még nincsen jelen. Az adaptív immunrendszer kis mértékben folyamatosan termel igen sokféle (kb. 1 milliárd!) antigén specificitású limfocitát. Ha egy adott antigén a szervezetbe kerül, az a néhány sejt, ami felismeri, intenzív osztódásba kezd, végrehajtó, ún. effektor sejteket hozva létre. Tehát az immunválasz idomul (adaptálódik) az aktuális fertőzésekhez. Az immunválasz során hosszú életű memóriasejtek jönnek létre, melyek védettséget biztosítanak a fertőzésen átesett szervezet számára. Az adott antigénnel való legközelebbi találkozáskor az immunrendszer már készen áll az antigén elleni specifikus immunválasszal, gyorsabban, intenzívebben és hatékonyabban reagál rá.
Mivel a sejtosztódások időigényesek, az elsődleges immunválasz lebonyolításához az antigénnel való találkozást követően akár egy-két hét is szükséges lehet. A későbbi újabb találkozások esetében azonban a másodlagos válaszreakció a memóriasejtek jelenléte következtében lerövidülhet néhány napra (3-5 nap).
A specifikus immunválasz kialakításáért a specifikus antigénreceptorral rendelkező limfociták felelősek.
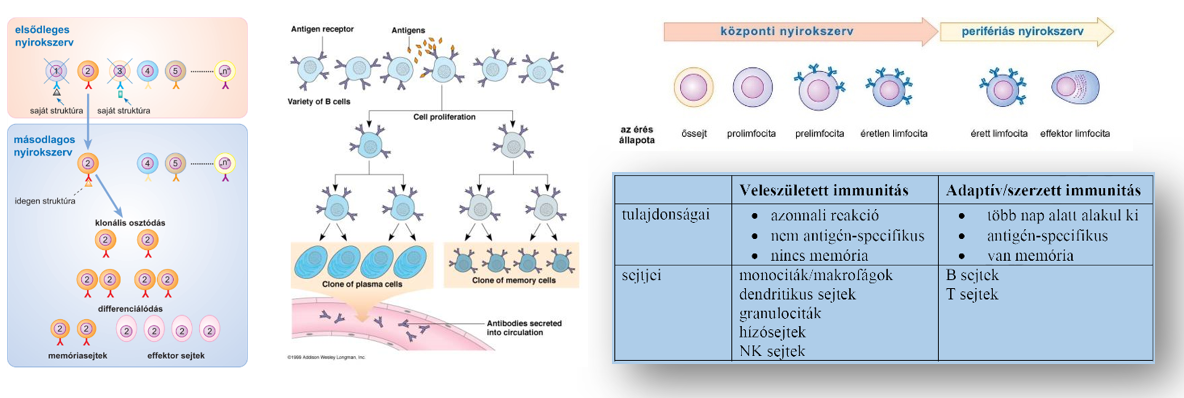 |
A limfociták érése
Az érési folyamatok során tanulják meg a nyiroksejtek megkülönböztetni a sajátot az idegentől. A steril közegű elsődleges nyirokszervek – vörös csontvelő, tímusz - a limfociták érésének színterei, itt jön létre a csak limfocitákra jellemző antigénfelismerő receptorok (109 nagyságrendű) nagyfokú sokfélesége.
B-limfociták klónszelekciója
A B-limfociták antitesteket termelnek. A vörös csontvelőben érnek be, mely érés során a sejtek sejthártyáján antigénreceptorok jelennek meg, melyek megegyeznek azzal az ellenanyaggal (antitest), melyet az illető sejtpopuláció elő tud állítani.
Tehát a B-limfociták az antigént a felszínükön található receptorként működő antitestekkel ismerik fel.
A B-limfocita érése során az antitestek kb. egymilliárd variációjából kiválaszt egyet, ezt kiteszi a sejthártyájára. Amelyik B-limfocita kapcsolódni képes valamely saját anyaggal, az apoptózissal elpusztul (klónszelekció).
T-limfociták klónszelekciója
A T-limfociták előalakjai a vörös csontvelőben fejlődnek, majd a sejtek a vérárammal a tímuszba – csecsemőmirigybe - kerülnek, ahol befejezik érésüket.
A T-sejtek érésük során „megismerkednek” a szervezet saját anyagaival mielőtt a keringésbe kerülnek. A csecsemőmirigyben eltöltött időszakot csak azok a T-sejtek "élik túl", amelyek képesek az MHC-molekulák felismerésére, miáltal képesek a saját-idegen megkülönböztetésére (klónszelekció).
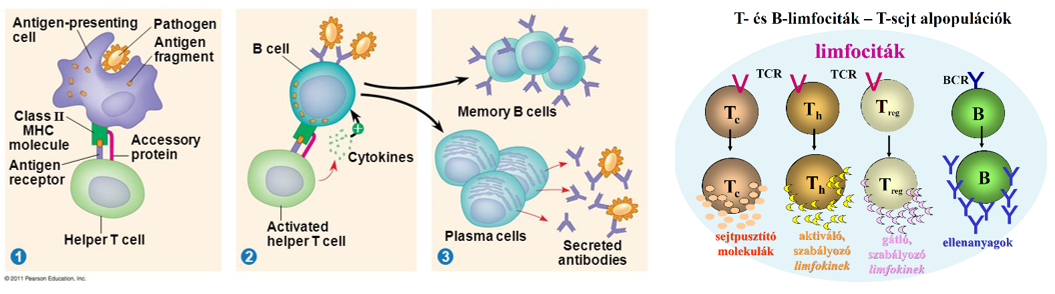 |
A limfociták aktiválódása
A megért – speciális antigénfelismerő receptorokkal rendelkező - T- és B-limfociták az elsődleges nyirokszervekből a keringés útján átkerülnek a másodlagos vagy perifériás nyirokszervekbe – nyirokcsomók, lép -, melyek a kórokozók lehetséges behatolási kapuinak megfelelően helyezkednek el a szervezetben. A limfociták a perifériás nyirokszervekben találkoznak a vér- és nyirokkeringés révén odajutó kórokozókkal, illetőleg azok jellegzetes molekuláival, aminek következtében aktiválódnak.
Az antigént felismerő, aktiválódott B-limfociták élénk osztódásba kezdenek és kétféle sejtpopulációt hoznak létre:
A nem aktiválódott sejtek rövidesen elpusztulnak (apoptózis).
Az aktiválódott T-sejtek többféle sejtpopulációt hoznak létre.
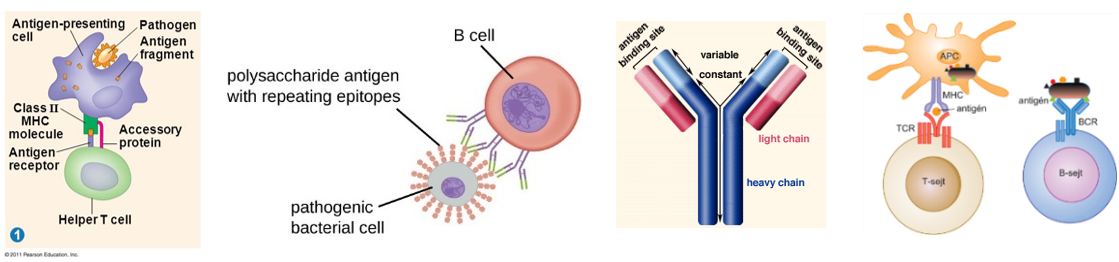 |
Hogyan ismerik fel az adaptív immunválasz sejtjei az immunogéneket?
T-limfociták kettős felismerése
Az antigének felismerésében nélkülözhetetlenek a makrofágok, mint antigénbemutató sejtek.
A makrofágok felismerik, bekebelezik, majd részben lebontják az antigéneket, majd valamilyen jellemző részletüket kijuttatják a sejthártya felszínére, pontosan az MHC struktúrákkal együtt.
Ennek a komplexnek – antigén-MHC - a kettős felismerése aktiválja az adaptív immunrendszer T-limfocitáit.
B-limfociták
A B-limfociták az antigént a saját maguk által termelt, felszínükön megtalálható, az antigén szerkezetével komplementer, receptorként működő antitestekkel ismerik fel.
Az antitestek (ellenanyagok, immunoglobulinok)
A B-nyiroksejtek által termelt antitestek molekulái Y alakúak.
Az antitest:
Az ellenanyagok termeléséért felelős B-limfociták és a sejtes immunitást hordozó T-sejtek eltérően ismerik fel az antigéneket.
49. tétel: Védőoltások |
Immunizálás
Immunizálás a fertőző betegségek megelőzésére, leküzdésére alkalmazott eljárás, melynek több formája ismert, lehet aktív vagy passzív, természetes, ill. mesterséges. Az immunizálás eredményeképpen a szervezetünk felkészül egy majdani lehetséges fertőzés következményeinek a leküzdésére, aminek köszönhetően megelőzhető a betegség kialakulása.
Az aktív immunizálás hátterében az immunmemória jelensége áll, mely a specifikus immunválasz során létrejött T- és B-memóriasejteknek köszönhető.
A memóriasejtek hosszúéletű limfociták, melyek megtalálhatók egyrészt az elsődleges nyirokszervekben, másrészt a periférián, a szövetekben, a másodlagos nyirokszervekben, a keringési rendszerben. Egy korábban - az elsődleges immunválasz során - már megismert antigén ismételt megjelenésekor intenzív osztódásba kezdenek és létrehozzák a különféle végrehajtó B- és T-sejteket, melyek gyorsan lebonyolítják a másodlagos immunválaszt.
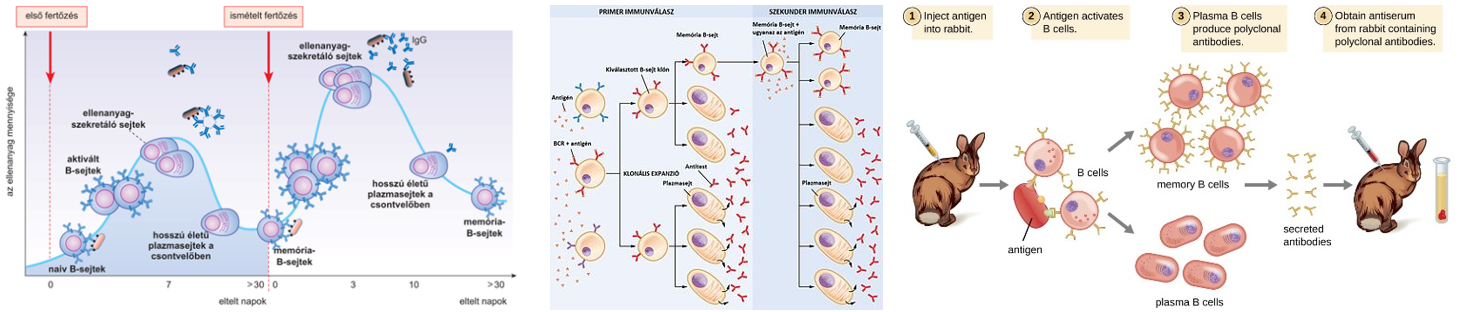 |
Mesterséges aktív immunizáláskor többnyire a kórokozó egy ártalmatlan, de ugyanakkor jellemző darabját juttatjuk be a szervezetbe oltóanyag formájában (ún. legyengített kórokozót), ezt az immunrendszer megjegyzi, kialakítja ellene a megfelelő immunválaszt, pl. ellenanyagot termel vele szemben, és amikor legközelebb a patogén megjelenik a testben, gyorsan lezajlik a kórokozót megsemmisítő immunfolyamat (másodlagos immunválasz) a kialakult immunmemóriának köszönhetően. Ezt a védőoltást gyakran kísérheti enyhe betegségtünet, láz, ami az immunrendszer működésének köszönhető. Ilyen pl. az influenza, hepatitis B elleni oltás. Hosszútávú védettséget biztosít, mint pl. BCG oltás.
Természetes aktív immunizáláson esünk át, mikor természetes úton megfertőződünk, majd az immunrendszerünk leküzdi a betegséget és kialakul az immunmemória, ilyen pl. a bárányhimlő ellen gyerekkorban megszerzett védettség.
Passzív mesterséges immunizálásról akkor beszélünk, ha más állatból kivont ellenanyagot juttatunk a szervezetbe, ilyen pl. tetanusz baktériumok elleni oltás, vérzéssel járó mélyebb sebek keletkezésekor. Ebben az esetben, mivel a saját immunrendszerünk nem játszik szerepet a védettség kialakításában (azaz passzív), nem alakul ki immunmemória, aminek következtében a hatás csupán néhány hétre korlátozódik, amig a beadott ellenanyagok a vérben le nem bomlanak. Ilyen továbbá az Rh-összeférhetetlenség esetén beadott vakcina.
Passzív természetes immunizálás folyamata során, természetes úton ellenanyag jut be a szervezetbe, pl. az anyatejjel csecsemőkorban, ill. méhlepényen keresztül a magzati korban.
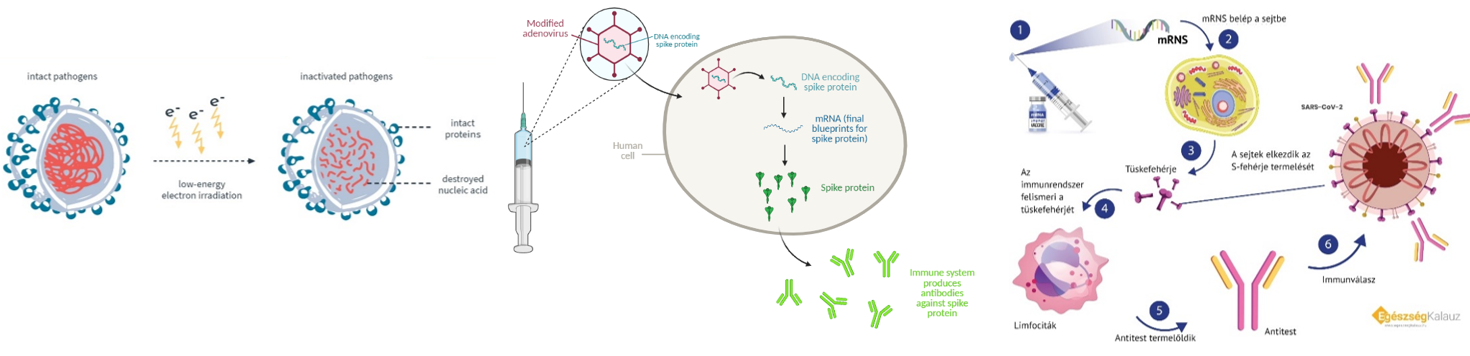 |
Első, másod- és harmadgenerációs vakcinák
Első generációs vakcinák: inaktivált kórokozó alapú vakcinák
Hagyományos, régi módszer, mely szerint inaktivált, ún. legyengített kórokozót juttatnak a szervezetbe, melynek felületén megtalálható molekulák beindítják az immunválaszt.
Az inaktivált kórokozók lehetnek genetikai anyagukban károsított patogének, melyek szaporodni nem képesek, de immunválaszt ki tudnak váltani. Ilyen pl. a veszettség elleni védőoltás.
Második generációs vakcinák
Ebben az esetben a kórokozó pl. vírus genetikai anyaga nélküli, immunválaszt kiváltó hatásos burokfehérjéket használnak. Erre példa a Hepatitis B elleni vakcina.
Harmadik generációs vakcinák
Ennek lényege, az immunválaszt kiváltó fehérjét a saját szervezetünk állítja elő, mivel csak a fehérje előállításához szükséges nukleinsav-molekulát juttatják be a szervezetbe.
Vektor alapú vakcinák
A vektorok – hordozók - olyan genetikai elemek, melyek képesek idegen DNS-t élő sejtbe juttatni. Vektoroknak kiválóan alkalmasak különféle vírusok, mint pl. az adenovírusok, melyek meghűléses tünetekkel járó, többnyire enyhe lefolyású légzőszervi megbetegedéseket okoznak.
Ebben az esetben a humán sejtekbe a vírus egyik burokfehérje génjét egy genetikailag módosított – ártalmatlanított – adenovírus közvetítésével juttatják be. A bejutatott vírus DNS szakasznak köszönhetően sejtek tehát legyártják a burokfehérjét. A termelődött fehérje – mint idegen anyag - megjelenve a szervezetben immunválaszt vált ki, aminek részeként pl. a B-limfociták nagy mennyiségben termelnek az idegen fehérje elleni antitesteket (ellenanyagokat), így amikor az igazi vírus megjelenik a szervezetben az immunrendszer már azonnal felismerni a vírus tüskéjét és gyorsan megindítja a fertőzés és a betegség elleni védekezést.
mRNS alapú vakcinák
A vírus egyik burokfehérjéjének genetikai információját egy lipidburokban található mRNS molekulával juttatják be emberi sejtekbe, melyek legyártják a burokfehérjét, amely a fent említett módon immunizálja a szervezetet.
50. tétel: A női nemi működések |
A nők belső nemi szervei
A nők nemi működése – szemben a férfiakéval – ciklusos, a petefészkek felváltva többnyire egy-egy petesejtet érlelnek havonta, melynek egymást követő fázisai az egész női nemi működésre, ill. a nemi szervek állapotára hatást gyakorolnak. A petefészkek, mint ivarmirigyek, a herékhez hasonlóan
A női nemi működés ciklusa kb. 28 napos, a pubertáskorban kezdődik, és kb. az 50. életévig tart, megszűnését klimaxnak vagy menopausának nevezzük.
A petefészek felépítése
A petefészek a medencében elhelyezkedő páros szerv, alakja és nagysága mandulára emlékeztet. A petefészeknek saját kivezetőcsöve nincs, a kilökődő petesejteket a petevezeték tölcsérszerűen kiszélesedő, a petefészekre ráboruló vége tereli magába.
A petesejtek képződése
A petefészekben, még az embrionális fejlődés alatt keletkező mikroszkopikus gömb alakú képződményeket, ún. elsődleges tüszőket találunk. Az elsődleges tüsző
Az elsődleges tüszők még magzati korban képződnek, a pubertáskorig, kb. 12-13 életévig változatlanok maradnak. Számuk a két petefészekben serdülőkorra kb. 200 000.
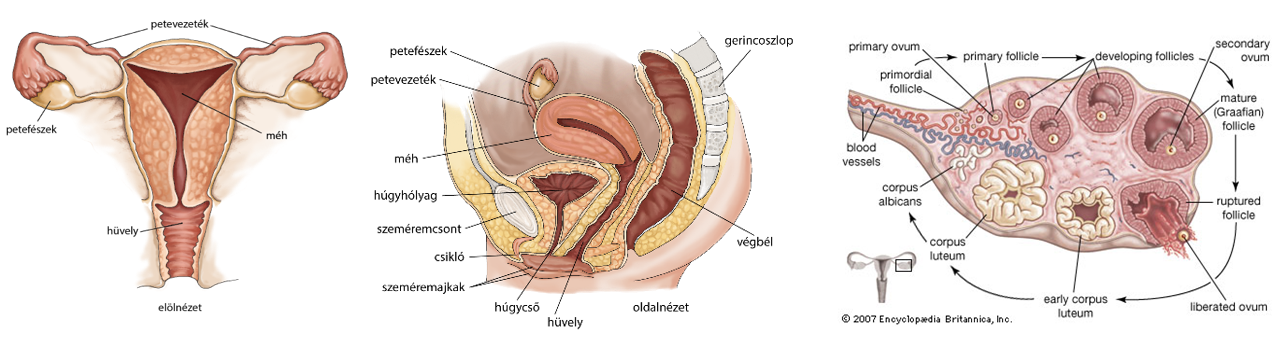 |
A tüszők érése
A tüszőérés folyamatában, a további fejlődésre kiválasztott (ún. besorozott) 10 tüszőből álló csapatból egyetlen (a legnagyobb FSH-érzékenységű) – néha több – tüsző ún. domináns tüszővé alakul, amely teljesen befejezi fejlődését. A csapat többi tagja apoptózissal elsorvad.
A domináns tüszőben az éretlen petesejt (elsőrendű oocyta) az első meiotikus osztódását befejezi és haploid érett petesejtté (másodrendű oocytává) alakul. A két petefészekben felváltva egy élet során kb. 400 petesejt érik meg.
Az érett haploid petesejt (másodrendű oocyta) kilökődik a petefészek felszínéről (ovuláció), és alkalmassá válik a megtermékenyítésre.
A peteérést és így az ivarérést az agyalapi mirigy tüszőserkentő hormonjának (FSH) növekedő koncentrációja váltja ki, ciklusosságát szintén a hipofízis működésének ciklusos jellege alakítja ki.
Tüszőérés fázisai
A pubertástól kezdve az FSH hatására a kiválasztott elsődleges tüszőben a petesejt térfogata megnő, a tüszőhámsejtek osztódni kezdenek, többrétegűvé válnak, létrejön a másodlagos tüsző (benne még diploid elsőrendű oocyta).
A tüszőhámréteg sejtjei tüszőfolyadékot termelnek, mely egy üregben halmozódik fel a tüszőhámsejtek között. A tüszőfolyadék tüszőhormonokat, ösztrogéneket tartalmaz.
A tüszőhámréteg tovább osztódik, a folyadékkal telt üreg egyre nagyobb lesz, melybe az éretlen „petesejt” (elsőrendű oocyta) egy réteg tüszőhámsejttel körülvéve bedomborodik (petedomb), ez a harmadlagos tüsző.
A tüszőrepedés előtti képződményt Graaf-féle tüszőnek nevezzük, benne az éretlen „petesejt” (elsőrendű oocyta) befejezi a meiózis I. fő szakaszát, és két sejtet hoz létre,
A meiózis második szakasza az ovulációkor kezdődik és a megtermékenyítéskor fejeződik be.
Lényeges különbség a petesejtek (oogenezis) és a hímivarsejtek (spermatogenezis) képződése között:
Az ovuláció (tüszőrepedés)
Az érett tüsző a petefészek felszínéhez közel helyezkedik el, azt erősen kidomborítja. A hipofízis sárgatestserkentő hormonjának (LH) hatására a tüsző fala felreped és az érett petesejt kilökődik a petefészekből. Ez a tüszőrepedés, az ovuláció, ami a ciklus kb. 14. napján következik be.
Az érett petesejt többnyire azonnal bekerül a petevezetőbe, annak szívó hatása miatt. A megtermékenyítés leggyakrabban a petevezető petefészekre boruló, kiszélesedő részén következik be, ha ez elmarad, akkor a petesejt kb. 24 óra múlva elhal, a tüsző visszamaradt része bevérzik, sejtjei osztódásnak indulnak, kitöltik a megmaradt üreget. Mivel a sejtekben élénksárga színű lipidcseppek halmozódnak fel, a kialakult képződményt sárgatestnek nevezzük, amely a progeszteron hormont termeli.
A sárgatest
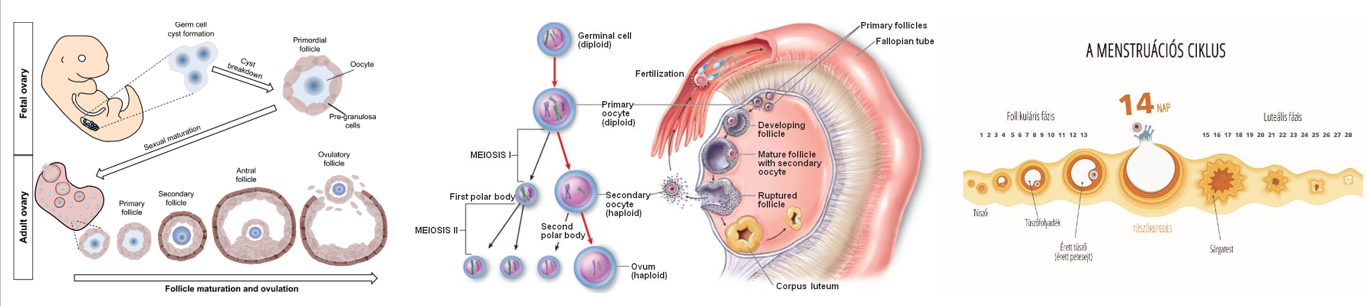 |
A petefészek hormonjai
A petefészek elsősorban szteroidhormonokat, ösztrogént és progeszteront, kisebb mennyiségben peptidhormonokat – inhibineket – termel. A petefészek hormontermelését
GnRH ↑ → (FSH ↑ + LH ↑) → petefészek szteroid- és peptidhormonok ↑.
A szabályozás a negatív visszacsatolás elvén alapszik.
A tüszőhormon (ösztrogén, pontosabban ösztradiol)
A progeszteron
Hatásai lényegében a terhesség elősegítésére és megtartására irányulnak.
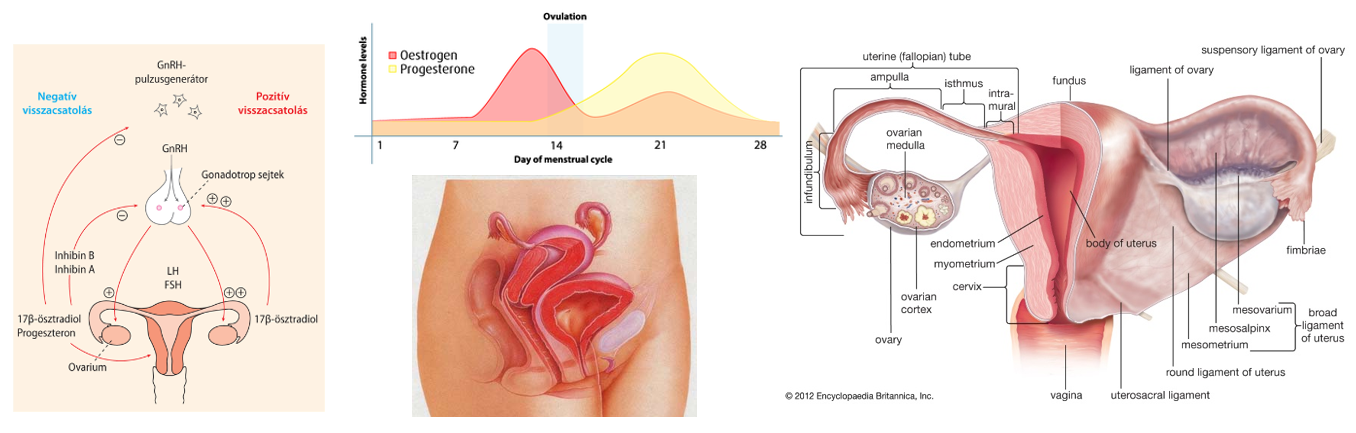 |
A petevezeték (méhkürt)
A petevezető a petefészket és a méhet összekötő, kb. 10-13 cm hosszúságú, ceruzavastagságú, izmos falú cső. A petefészekkel nincs összenőve, arra rojtos szélű, kiszélesedő végével ráborul. A petevezető hámja egyrétegű csillós hengerhám, ahol a csillók csapkodása a vezeték folyadéktartalmát a méh felé hajtja. A csillók csapkodása és a csőfal simaizomrétegének perisztaltikus mozgása miatt létrejövő szívóerő a petefészek felszínéről kilökődő petét beszippantja és azt a méh felé továbbítja.
A méh
Körte alakú, szűk üregű, a húgyhólyag fölé hajló izmos falú szerv. Megkülönböztetjük rajta a petevezetők nyílása felé kiszélesedő testet és a lefelé elkeskenyedő henger alakú nyakat. A méhnyak bedomborodik a hüvelybe, nyílását méhszájnak nevezzük.
A méh fala 3 rétegű,
A méhnyálkahártya hámja egyrétegű hengerhám, helyenként csillóval borítottan. A hám alatti kötőszövet összekapcsolja a hámot az izomréteggel, sejtdús, a hámból benyúló csöves mirigyekben gazdag kötőszövet. A méh nyálkahártyájának állapota nagymértékű változékonyságot mutat, melynek jellege ciklusos, fázisait a petefészekhormonok szintje határozza meg.
A méhnyálkahártya menstruációs ciklusa
A méhnyálkahártya állapota a 28 napos ciklus alatt folyamatosan változik. A ciklus első napja a menstruáció első napja.
Amennyiben
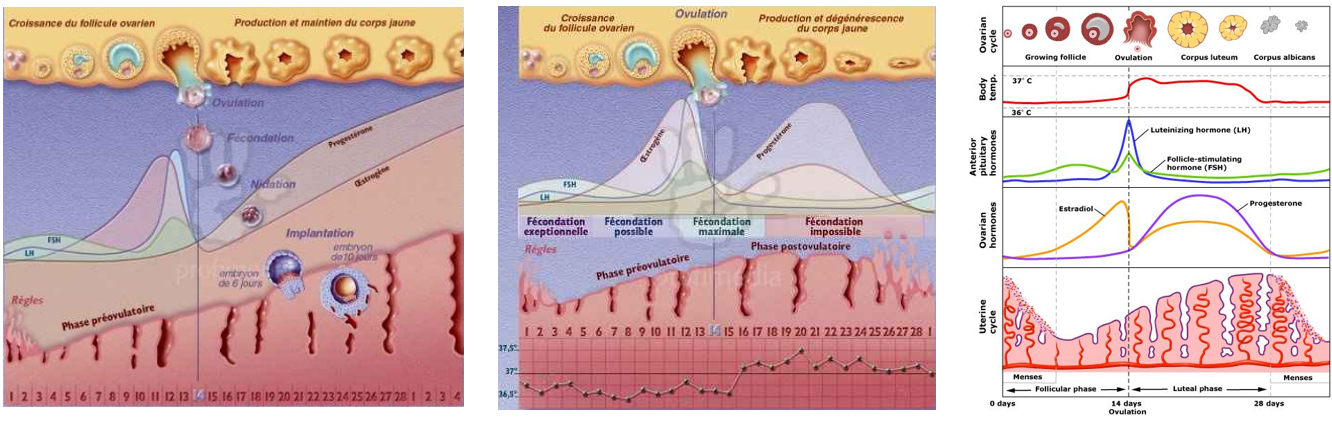 |
A női nemi működés hormonális szabályozása
A ciklus eseményeinek egymásutániságát a GnRH, az FSH, az LH és a három petefészekhormon koncentrációjának meghatározott, ciklikus változása eredményezi.
A különféle hormonok nemcsak egymás termelődésének a mértékére hatnak – pozitív vagy negatív visszacsatolással –, hanem befolyásolják a célsejtek receptorainak a számát, és ezzel a hormonérzékenységét.
A ciklus elején, a menses első napjaiban a petefészekhormonok – ösztrogén és progeszteron – koncentrációja a vérben alacsony.
Ennek következtében a hipofízis tüszőserkentő hormonjának elválasztása felszabadul a gátlás alól, szintje a vérben emelkedik, gyorsul a tüsző érése.
A fejlődő tüsző termeli az ösztrogént, ami a ciklus közepén pozitív visszacsatolás révén fokozza az FSH, ill. az LH elválasztását (úgy, hogy hatására nő az FSH-t és az LH-t elválasztó sejteken a GnRH receptorok száma).
A sárgatestserkentő hormon koncentrációja az ovuláció előtt 24 órával maximális, hatására bekövetkezik kb. a 14. napon a tüszőrepedés.
A létrejövő sárgatest megkezdi a progeszteron termelését.
A ciklus második felében a progeszteron és az ösztrogén negatív visszacsatolással gátolja az FSH és az LH termelődését, ezért ezek mennyisége csökken.
Eme tény az alapja a hormonális fogamzásgátlásnak; a tabletták megfelelő arányú ösztrogén- és progeszterontartalma gátolja a tüszőserkentő hormon, ill. a sárgatestserkentő hormon elválasztását, aminek köszönhetően a tüszőérés is gátlás alá kerül, így nem érik meg újabb petesejt, elmarad az ovuláció.
Mivel az LH mennyiségének a csökkenése miatt a sárgatest sorvadásnak indul, csökken a progeszteron- és az ösztrogénszint, ezért bekövetkezik a menses, a tüszőserkentő hormon termelése felszabadul a gátlás alól, szintje a vérben újra nőni kezd. A növekedő FSH-szint előkészíti az elkövetkezendő ciklusokban szerephez jutó tüszőket is.
A hüvely
Kb. 7-9 cm hosszúságú, tágulékony (szülés), izmos falú cső. Falát vastag simaizom alkotja, nyálkahártyájának hámja többrétegű, hormonhatásoktól (ösztrogén) változó mértékben elszarusodó hám. A hüvelyi ciklus lehetőséget biztosít arra, hogy hüvelykenet vizsgálatával a ciklus egyes fázisaira lehessen következtetni. A hüvelyváladék kémhatása savas (tejsavbaktériumok), ami megvédi a női ivarutakat elsősorban a gombás fertőzésektől.
Külső nemi szervek
A külső nemi szerveket lényegében a gáttájék hosszanti hasadéka alkotja, melyet a nagy és a kis szeméremajkak zárnak közre. A gát a medencefenék bonyolult izom- és kötőszöveti záró rendszere, amit a végbél, a hüvely és a húgycső fúr át (szűkebb értelemben a hüvely és a végbélnyílás közti terület).
A szeméremdombot és a nagy szeméremajkakat – ezek a férfi herezacskójának feleltethetők meg – másodlagos szőrzet fedi, ami a pubertáskorban alakul ki az ösztrogén hatására. A nagy szeméremajkak között találhatók a húgycső nyílását, ill. a hüvely bemenetét határoló kis szeméremajkak. A hüvelybemenetben található mirigyek váladéka nemi izgalom alkalmával megnedvesíti a hüvely nyílását, megkönnyítve a közösülést. A csikló a húgycső nyílása felett helyezkedik el, fejlődéstanilag a hímvesszőnek feleltethető meg, merevedésre képes, ingerlésével a nemi izgalom fokozható.
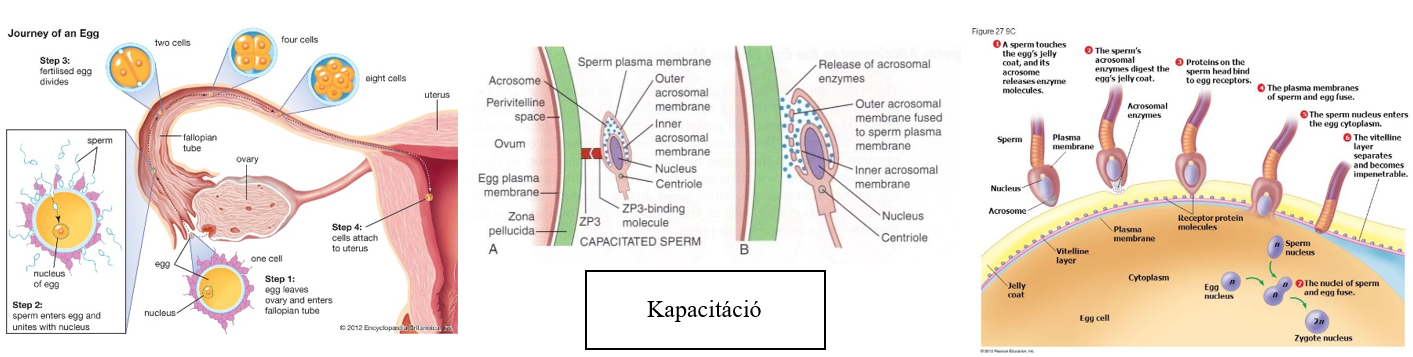 |
Az ember embrionális fejlődése
Az ember embrionális fejlődése 266 nap, az utolsó mensestől számítva a várandósság – melyet az utolsó menses első napjától számolnak - 280 nap.
Az egyedfejlődés korai szakaszai, ill. eseményei
A megtermékenyítést közvetlen megelőző folyamatok
A női nemi ciklus 14. napján bekövetkezik az ovuláció, melynek során a petefészek felszínéről kilökődik
(A petesejt a meiózis első osztódási fázisán túl van, a sejtek már haploidok, de még 2 kromatidás kromoszómákkal).
A petesejt a méhkürtbe – petevezetőbe - kerül, majd a petevezető perisztaltikus mozgásának és a csillós hám működésének eredményeképpen elindul a méh felé.
A hímivarsejtek a közösülés (koitusz) alkalmával általában a méhszáj környékére kerülnek.
A megtermékenyítéshez szükséges, hogy a hímivarsejtek érésének utolsó fázisa, a kapacitáció is végbemenjen, melyet a női ivari utak ionösszetétele, pH-ja és hormonális hatások váltanak ki.
A folyamat során a hímivarsejtek membránjában megváltozik a fehérjék összetétele, a petesejthez való kötődést elősegítő molekulák jelennek meg. A spermiumok anyagcseréje fokozódik és jelentősen megnő a Ca2+ felvétele, ami az akroszóma reakció végbemeneteléhez szükséges. A kapacitáció végén a spermium membránja destabilizálódik.
A megtermékenyítés leggyakrabban a petevezeték kiszélesedő rojtos részében, a petefészekhez közel történik. Abban az esetben, ha az embrió a méh üregén kívül – petefészek felszínén, petevezetőben, hashártyán stb. – tapad meg, méhen kívüli terhességről beszélünk.
Az ejakuláció alkalmával 1,5-5 ml ondó jut be a női ivarutakba, ml-enként kb. 150 millió hímivarsejttel, azonban a petesejtet csak töredékük éri el. A petesejt kb. 24, a hímivarsejtek kb. 48 óráig alkalmasak a megtermékenyítésre, utána elhalnak. Az ondó kémhatása lúgos (prosztata), amely kedvez a spermiumok mozgásának, azonban a hüvely savas váladéka gátolja azt, ugyanakkor hatására az ondófolyadék besűrűsödik, ami meggátolja a női ivarutakból történő kifolyást. A női orgazmus része, hogy a méh simaizomzata ritmikusan összehúzódik, elősegítve a hímivarsejtek feljutását a petevezető felé. A hímivarsejtek mozgásirányát kemotaxis és az áramlással szembeni haladás, a reotaxis határozza meg.
A megtermékenyítés
A haploid (23) hímivarsejt és a haploid (23) petesejt egyesülését megtermékenyítésnek nevezzük, melynek eredménye a diploid (46) zigóta.
A petesejt a 0,1 mm átmérőjével az emberi szervezet legnagyobb sejtje, citoplazmájában a szokásos sejtalkotókon kívül – belső membránrendszer, sejtmag, mitokondriumok stb. – szénhidrátokból, fehérjékből, és lipidekből álló tartaléktápanyag, az ún. szikanyag található. A petesejtet egy glikoproteidburok, a fénylő réteg veszi körül, amit a már említett tüszőhámréteg borít be. A tüszőhámréteg sejtjeit túlnyomórészt a hialuronsav nevű poliszacharid tartja össze.
A petesejtet egyszerre akár több száz spermium is körülveszi, azonban a petesejt belsejébe csak egy spermium juthat be, amit többféle mechanizmus biztosít.
Mivel a petesejtbe csak a spermium feji része kerül be, a nyak, a középdarab és a farok kívül marad, így a zigóta mitokondriumai kizárólag az anyától származnak.
Barázdálódás, a szedercsíra és a hólyagcsíra kialakulása
Magzatburkok
Az embriót, majd a magzatot az embrionális fejlődés alatt 2 burok veszi körül.
A külső magzatburok (irhahártya, chorion)
A szedercsíra külső sejtrétegéből alakul ki. Felszíne a sejtek osztódása miatt egyre bolyhosabbá válik (innen a neve). Feladata
A hCG a vizeletből a 10. nap után kimutatható, így megállapítható a várandósság. Pozitív próba esetén a várandós nő vizeletében megtalálható hormon összekapcsolódik a terhességi tesztek hCG-ellenanyagával, ami jól látható színváltozást eredményez, a kontroll mellett plusz csík vagy kör jelenik meg a teszten.
A belső magzatburok (magzating, amnion)
Az embriót tartalmazó amnionüreg fala. Termeli a magzatvizet, ami
A magzatvíz három óránként kicserélődik. A magzatvíz vizsgálata fontos információt nyújthat a magzat esetleges kóros fejlődéséről (a Down-szindróma a 16. héten vételezett mintából kimutatható). Később a két burok összenő és kialakul a közös magzatburok.
A méhlepény, placenta
A fejlődő magzat rögzítését, táplálását biztosító korong alakú, szivacsos, rendkívül vérbő szerv, mely progeszteront és ösztrogént termel. Kialakulásáig embrióról, utána magzatról beszélünk.
Fejlődése a beágyazódást követően megkezdődik, teljes fejlettségét a 3. hónapra éri el. A beágyazódást követően az embriócsomó feletti chorionsejtek intenzíven osztódnak, egyre elágazóbb bolyhok jönnek létre. A bolyhok növekedve felszakítják a méhnyálkahártya ereit, véröblök jönnek létre, aminek következtében az anyai vér körülmossa a chorion bolyhait (nyílt keringés).
A bolyhok hajszálereivel a magzati keringés a köldökzsinóron keresztül áll kapcsolatban.
A köldökartériák a magzat felől salakanyagokban gazdag vért szállítanak a bolyhokba, ahol a salakanyagok átjutnak az anyai keringésbe.
Az anyai keringésből oxigén és tápanyagok kerülnek a bolyhokon át a magzati vérbe. Az oxigénátadást a magzati hemoglobin nagyobb oxigénkötő-képessége biztosítja. A méhlepényben felfrissült vér a köldökvénán keresztül jut vissza a magzathoz.
A terhesség vége felé a méhlepény öregszik, leépül, ami veszélyessé teszi a túlhordást.
51. tétel: A férfi nemi működések |
A férfiak belső nemi szervei
 |
A herék
A herék, mint ivarmirigyek (gonádok), kettős feladatot látnak el,
Működésüket – hímivarsejt-termelést, nagyobb mennyiségű tesztoszteron termelését – 12-13 éves korban kezdik meg az agyalapi mirigy tüszőserkentő, ill. sárgatestserkentő hormonjai növekvő koncentrációinak hatására. A here működése folyamatos, a pubertáskortól kezdve egészen időskorig, az élet végéig tart.
A herék fejlődése
A herék a hasüregben jönnek létre az embrionális fejlődés 3 hónapját követően, majd a születés előtt a lágyékcsatornán keresztül a herezacskóba süllyednek, mivel működésük a testhőmérsékletnél 2-3 °C-kal alacsonyabb körülményeket igényel. A herezacskó falában található simaizomréteg hidegben összehúzódással, melegben elernyedéssel segíti az optimális hőmérséklet fenntartását.
A here felépítése
A here kanyargós lefutású, ún. herecsatornácskákból áll. A herecsatornácskák falában képződnek a hímivarsejtek. A csatornácskák közti kötőszövetben tesztoszteront termelő sejtcsoportok helyezkednek el, amiket Leydig-féle köztes sejteknek nevezünk.
A hímivarsejtek, a spermiumok képződése
A haploid hímivarsejtek a herecsatornák szélén található diploid, ún. csírahámsejtek meiózisával jönnek létre a nemi érést követően az agyalapi mirigyben termelődő tüszőserkentő hormon (FSH) hatására (kb. 70 nap alatt). A fejlődés különböző stádiumaiban levő egyes fejlődő ivarsejtformák a csatornácskák alapi részétől fokozatosan tolódnak felfelé, egészen a csatornák üregéig. A hímivarsejtek további fejlődése és érése a herecsatornák falában lévő Sertoli-féle dajkasejtek az üreg felé néző kehelyszerű tágulatában zajlik, itt alakul ki végleges formájuk. A Sertoli-féle dajkasejtek működését a tesztoszteron irányítja. Az FSH hormon ivarsejtképzést serkentő hatása is a Sertoli-sejteken keresztül érvényesül, ugyanis ezek tartalmazzák a hormon megkötésére alkalmas receptorokat.
A hímivarsejtek felépítése
A hímivarsejtnek alapvetően 3 fő része van: fej, középrész, farok.
A fej lényegében a haploid örökítőanyagot magába foglaló sejtmagot és az ún. akroszómát tartalmazza, ami a petesejtbe történő behatolást segítő enzimeket – hialuronidáz – tárolja.
A középrész tartalmazza a mozgáshoz energiát – ATP-t – szolgáltató mitokondriumokat.
A farok tulajdonképpen egy ostor, a hímivarsejtek mozgását teszi lehetővé.
 |
A mellékherék
A herecsatornácskák összeszedődve a mellékherékbe torkollanak, amelyek ív alakban simulnak a herék hátsó falára. A mellékherében tárolódnak (8-12 napig) és fejezik be érésüket – utóérés – a spermiumok, itt alakul ki a mozgásképességük (a Leydig-sejtek ösztrogénjének hatására) és itt válnak alkalmassá a megtermékenyítésre. Az utóérés során a spermium energiafogyasztása lecsökken, nyugalmi állapotba kerül.
Az ondóvezetékek és az ondóhólyagok
Az ondóvezetékek a mellékherékből folytatódó, a spermiumok továbbítására – perisztaltikus mozgással – szolgáló izmos falú csövek.
Az ondóhólyagok (seminal vesicle) az ondóvezetékekhez csatlakozó páros mirigyek. Az ondóhólyagok lúgos váladéka adja az ondó egy jelentős részét, tartalmaz fruktózt, a hímivarsejtek fő energiaforrását, ill. prosztaglandinokat, melyek serkentik a spermiumok mozgását és a méhfal összehúzódását, elősegítve ezzel a megtermékenyítést. E váladékkal a spermiumok csak az ejakulációkor keverednek, aminek hatására mozgásuk aktiválódik.
A prosztata (dülmirigy)
A prosztata a legnagyobb járulékos mirigye a férfiak belső nemi szervrendszerének, a húgyhólyag alatt helyezkedik el. Páratlan, sok simaizmot tartalmazó, lúgos kémhatású, a spermiumok mozgását fokozó, a hüvely savas kémhatását semlegesítő váladékot termelő szerv. Fala simaizomrétegének összehúzódása továbbítja az ondófolyadékot ejakulációkor a húgycsőbe.
A prosztata területén egyesül egymással a húgyhólyagból eredő páratlan húgycső és az ondóhólyag váladékát is felvevő páros ondóvezeték. Innen a húgyutak és az ivarutak közös szakaszban, a péniszben folytatódnak.
A prosztata után a húgycsőhöz kapcsolódnak a páros, borsó nagyságú Cowper-féle járulékos mirigyek, váladékuk az erekció alkalmával síkossá teszi a pénisz végét, elősegítve a közösülést, a koituszt.
Az ondó (sperma)
Az ondó a here és a járulékos mirigyek által termelt váladék, tartalmazza
Egy ejakuláció alkalmával kilövellt mennyisége 1,5-5 ml, ml-enként kb. 150 millió spermiummal.
Külső nemi szervek
A férfi külső nemi szerve a pénisz vagy hímvessző, amely merevedésre képes párzószerv. Három fő része van: a töve, a teste és a vége, a makk. A hímvessző bőre igen vékony, szőrtelen, a pénisz csúcsán visszahajlik, és egy redőt képez, ez a fityma. A fityma nem borítja be teljesen a makkot, részben visszahúzódik. A fitymát egyes népek rituális okokból eltávolítják (körülmetélés).
A péniszben ún. erektilis szövetből felépülő hengeres testek találhatók, ezek közül a barlangos testek páros képződmények, a húgycsövet körülvevő szivacsos test pedig páratlan.
Nyugalmi állapotban nagyon kevés vér folyik át a pénisz érrendszerén, mivel az artériák nagyrészt zárva vannak. Nemi izgalomkor az artériás véráramlás a verőerek tágulása miatt fokozódik, ugyanakkor a vénás visszaáramlás – a vénák összenyomódása miatt – csökken, aminek eredményeként az erektilis szövetek üregei megtelnek vérrel, a pénisz megmerevedik, ez az erekció. A merevedés kiváltásában elsősorban keresztcsonti paraszimpatikus idegek aktivitása játszik szerepet.
A nemi izgalom csúcspontján – szimpatikus hatásra – következik be az ondó kilövellése, az ejakuláció, amelyet egyrészt a gátizmok, másrészt a prosztata, az ondóvezetékek és az ondóhólyagok együttes összehúzódása tesz lehetővé.
 |
A férfi nemi működés hormonális szabályozása
A here, mint ivarmirigy, endokrin szervnek tekinthető, mivel
A tesztoszteron szteroidhormon,
A férfiakra jellemző másodlagos nemi jellegek:
A here működését – a spermiumok és a tesztoszteron termelődését – közvetlenül az agyalapi mirigy szabályozza negatív visszacsatolással.
A nemi működéseket, az ivari hormonok elválasztását az agyalapi mirigy FSH- és LH- termelődésének szabályozásával a hipotalamusz is befolyásolja közvetett módon, serkentő neuroszekrétumok – GnRH (Gonadotropin releasing hormon) – elválasztása révén.
A here tesztoszteron termelése az egyes életszakaszokban eltérő, nagyobb mértékben serdülőkorban kezd termelődni, elválasztása folyamatos és élethosszig tart. A hímivarsejtek termelése a pubertáskortól kezdődően – változó intenzitással – folyamatos, és szinte élethosszig tart. A plazma tesztoszteronszintjének statisztikai átlaga alacsonyabb idős-, mint fiatalkorban, ugyanakkor a szexuális aktivitás és az androgénhormonok plazmaszintje között nincs kimutatható összefüggés (Fonyó).
52. tétel: Populáció növekedési modellek, r- és K-stratégia |
A populáció (népesség)
A populáció egy adott szempont szerint azonosnak tekintett egyedek halmaza.
A faji minősítésű, genetikai meghatározás szerint,
A populáció fogalmának ökológiai meghatározása tágabb értelmezésű, valamilyen vizsgálati szempont alapján határozzuk meg az egyedek csoportját, pl. azonos szempont szerint táplálkozó, de több fajból álló állatok egy élőhelyen meglévő csoportja is lehet populáció.
A populáció tulajdonságai az egyedre nem, csak az egyedek csoportjára jellemzők.
A populációk változásai, populációdinamika
A populációkra szüntelen változás jellemző, legjellemzőbb a populációk időbeli egyedszám változása. A populációk egyedszám változása függ (a környezeti tényezők közvetlen hatását figyelmen kívül hagyva):
Természetesen, ezenkívül a populáció egyedszáma függ, közvetlenül
Születési ráta
A születések számát a születési rátával adhatjuk meg, amely az egységnyi idő alatt született utódok száma, pl.
A születési ráta függ,
 |
A populációk szaporodóképessége, korlátlan és korlátozott növekedési modellek
Alacsony egyedszám esetén, ideális körülmények között a születések számából, azaz a születési rátából következtethetünk a potenciális szaporodóképességre.
Környezet eltartóképessége (K) azt jelzi, hogy az adott környezetben mekkora egyedszámú populáció képes tartósan fennmaradni (N=K).
Az eltartóképességet egyedszámmal jellemezzük.
Kis egyedsűrűség esetén a populáció növekedése egy darabig exponenciális jellegű. Azonban az egyedsűrűség növekedésével a születések száma csökken, ami miatt a populáció növekedési görbéje ellaposodik, majd az egyedszám a K érték körül állandósul.
A környezet eltartóképességét egy adott területen egy adott populációra vonatkoztatjuk. Ez alapvetően faji jellegtől (méret, táplálékigény, territoriális viselkedési szokások) függ. A valóságban az egyedszám az eltartóképesség környékén ingadozik (fluktuál).
A szaporodási ráta (r)
A szaporodási rátát a születések és a halálozások különbségéből kapjuk meg. A szaporodási rátából a populációk szaporodóképességére, növekedésére következtethetünk.
A szaporodási ráta
A létszám (sűrűség) növekedése következtében a populáció
A születések és a halálozások száma – a szaporodási ráta - döntően befolyásolja a koreloszlást.
A populáció koreloszlása
A populációt alkotó egyedek különböző korosztályokba (korcsoportokba) sorolhatók, és ezek egymáshoz viszonyított nagyságai határozzák meg a korösszetételt. A korösszetételt a korfával ábrázoljuk, ahol a
Típusai:
Növekvő, vagy fiatalodó populáció:
Stabil populáció:
Kihaló, vagy elöregedő populációban a középkorú és idős egyedek száma jóval meghaladja a fiatalokét.
 |
Gradáció
A gradáció a populáció egyedszámának ugrásszerű emelkedése, melynek során a populáció nagysága jelentősen meghaladja a környezet eltartóképességét. A hirtelen növekedést, a kitörést mindig összeomlás követi az eltartóképesség korlátai miatt. Rovarokra (vándorsáska, levéltetvek, gyapjaslepke), egérfélékre, algákra (vízvirágzás) jellemző.
Szaporodási stratégiák
A szaporodási stratégiájuk alapján a populációk nagy része alapvetően két csoportba sorolható.
A K-stratégia (K – konstans (állandó))
Az r-stratégia (r – reproduktív)
53. tétel: Ökológiai kölcsönhatások |
Környezeti kölcsönhatások
Azt a térbeli helyet, ahol az életközösségek előfordulnak, élőhelynek (biotóp) nevezzük. Az élőhely egyes elemei – környezeti tényezői - nem egyformán hatnak az életközösségekre, egyesek közömbösek, mások pedig alapvetően meghatározzák az élőlények előfordulását, életlehetőségeit, ezért csak a ténylegesen ható tényezők jelentik a környezetet, amit ökológiai környezetnek nevezünk. Az élőlényekre ható környezeti tényezők lehetnek:
A környezeti tényezők értékei nem állandóak, térben és időben is változást mutatnak. A változások lehetnek periodikusak (pl. aszpektusok), ill. egyszeri, előrehaladók (pl. szukcesszió).
Az ökológiai tűrőképesség (tolerancia)
Grafikusan szemléltethető:
Szűktűrésűek (specialisták) azok a fajok, amelyek az adott környezeti tényező értékének megváltozását csak szűk tartományon belül képesek elviselni, azaz a tűrőképesség minimum és a maximum értékei közel vannak egymáshoz. Ilyenek az indikátor élőlények.
Az indikátor szervezetek azok az élőlények, amelyek jelenlétükkel (vagy éppen hiányukkal), egyedszámukkal, viselkedésükkel jelzik a környezet valamely tulajdonságát (ásványianyag-tartalom, kémhatás stb.). Az indikátor fajok szűktűrésűek egy adott tényezővel szemben.
Zuzmók a SO2-al, korallok a sótartalommal szemben szűktűrésűek.
Tágtűrésűek (generalisták, kozmopoliták)
A tágtűrésű fajok széles határok között képesek elviselni az adott környezeti tényező változását. Például, a cetek a hőmérsékletre nézve tágtűrésűek, tekintve, hogy többnyire a sarkvidék tengereiben élnek, de szaporodni a trópusi vizekbe járnak.
A tűrőképesség fajonként eltérő. A különféle populációk az egyes környezeti tényezőkkel szemben szűktűrésűek, ugyanakkor másokkal szemben tágtűrésűek. A foltosszárnyú maláriaszúnyog a hőmérséklettel szemben tágtűrésű, ugyanakkor a páratartalom változását csak nagyon szűk tartományban viseli el.
 |
Élettani és ökológiai tűrőképesség
Az élettani tűrőképesség a populáció egyedeinek kísérleti körülmények között meghatározott, az adott környezeti tényezővel szembeni élettani tűrőképességét mutatja. A vizsgálatok általában egy környezeti tényezőre vonatkoznak, miközben a többi környezeti tényezőt változatlanul hagyják. A fajok genetikailag kódolt fiziológiai sajátossága.
A természetben a populációk nem tudják teljesen kihasználni az élettani tűrőképességük által meghatározott lehetőségeiket, mivel az adott tartományban számos más hasonló tűrőképességű populáció lehet jelen. Így a populációk felosztják egymás között a hasonló környezeti tényezők által meghatározott teret, és az adott populáció az élettani tűrőképessége által meghatározott térnek csupán kis hányadát foglalja el, amit az ökológiai tűrőképességgel jellemezhetünk. Értékét általában befolyásolja a többi környezeti tényező is.
Tehát az élettani tűrőképesség sokkal szélesebben határozza meg a populációk előfordulási lehetőségeit, mint a ténylegesen megvalósuló ökológiai tűrőképesség.
A hasonló környezeti igényű populációk szüntelen versengésben vannak a korlátozott mértékben rendelkezésükre álló erőforrásokért. Rövidtávon mindkét fél számára korlátozó jellegű a verseny. Hosszútávon az egyik populáció kiszorulását eredményezheti.
 |
Ökológiai niche, ökológiai tér
A környezeti tényezők olyan n-dimenziós tere, ahol a populáció egyedei előfordulhatnak. Minden dimenzióját egy olyan környezeti tényező adja, amely a populáció előfordulását meghatározza. A niche (ejtsd: nis) az ökológiai térnek az a része, amelyet a populáció jelenlétével kitölt. A niche francia szó, fülkét jelent.
Egy populáció ökológiai niche-ét meghatározza
A populációk ökológiai niche-ét alapvetően tehát az egyedek különböző környezeti tényezőkkel szembeni tűrőképessége határozza meg.
Ebben az esetben is az előzőekhez hasonlóan különbséget teszünk élettani (fundamentális), ill. ökológiai (realizált) niche között.
Az élettani niche a populáció egyedeinek kísérleti körülmények között meghatározott, az adott környezeti tényezőkkel szembeni élettani tűrőképességétől függ.
Az élettani niche a többi fajjal (populációival) való kölcsönhatás nélkül, az ökológiai niche ugyanakkor e kölcsönhatások figyelembevételével mutatja meg a faj életlehetőségeit.
Az élettani niche sokkal szélesebben határozza meg a populációk előfordulását, mint a ténylegesen megvalósuló ökológiai niche.
Az ábrákon két-, ill. háromdimenziós ökológiai teret láthatunk.
Minden fajnak saját, rá jellemző ökológiai niche-e van; ha két populációnak azonos az ökológiai niche-e, akkor e két csoport között kompetíció alakul ki.
A hasonló igényű populációk felosztják egymás közt a hasonló élettereket úgy, hogy a különböző biotópok legalább egy lényeges tulajdonságukban különböznek. Ez a niche-szegregáció.
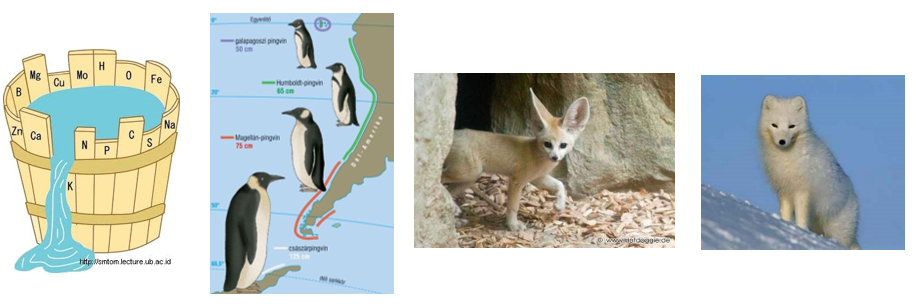 |
Gauze-elv, a kompetitív kizárás elve
A Gauze-elv szerint tartósan nem létezhet egy életközösségben (élőhelyen) két, teljesen azonos ökológiai igényű, azaz niche-ű populáció.
Másképp: két faj ugyanazt a niche-t nem foglalhatja el.
Minimum-elv
Justus Liebig növények fejlődését vizsgálva fogalmazta meg a minimumtörvényt: mindig az a tényező – tápanyag – határozza meg a növények fejlődésének az ütemét, amely a szükségletekhez képest a legkisebb mennyiségben áll a növény rendelkezésére. Hiába jut hozzá a növény az összes tápelemhez, mert ha, például a kalcium nem elegendő a talajban, akkor a növény végül elpusztul.
A hordó dongái jelképezik az egyes ásványi anyagok mennyiségét, a hordóba tölthető víz mennyisége pedig a növény fejlődésének a mértékét. Mint az ábrán látható a növény szükségleteihez képest a kálium található legkisebb mennyiségben, tehát ebben az esetben ez határozza meg a növény növekedését.
Liebig-féle minimumtörvény kiterjesztése: bármely biológiai folyamat sebességét az a tényező korlátozza, amely a szükségletekhez képest a
Ha bármelyik környezeti tényező eléri, vagy megközelíti a minimum vagy maximum értéket, akkor a többi tényező hatását is korlátozza.
A testtömeg, a testfelület, a testfüggelékek (fül, farok, végtagok) mérete és az élőhely átlaghőmérsékletének az összefüggése
Bergmann-szabály: A hidegebb területen élő fajok testmérete nagyobb a melegebb területen élő rokon fajokénál (a sarki pingvinek nagyobbak, mint az egyenlítőiek).
A hőtermelés a térfogattal (testtömeggel), a hőleadás a testfelülettel arányosan változik. A testméretek növekedésével a térfogat 3. hatvánnyal, a felület csak a 2. hatvánnyal nő. Tehát a felszín-térfogat arány alkalmazkodik annak a környezetnek az átlagos hőmérsékletéhez, ahol az állat él.
Allen-szabály: A hidegebb területek élőlényeinek testfüggelékei – melyek hőleadó felületek - kisebbek, mint a melegebb területen élő rokonaiké (a sivatagi róka füle nagyobb, mint a sarki rókáé).
A populációk kölcsönhatásait részletesen lásd 5.2. jegyzet.
 |
54. tétel: Egy erdő aszpektusai |
A társulások periodikus – visszatérően ismétlődő - időbeli megváltozása
A társulások évszakok váltakozásával kapcsolatos, periodikusan ismétlődő állapotai az aszpektusok. Az aszpektusokat a környezeti tényezők – fény, hőmérséklet, csapadék, páratartalom - periodikusan ismétlődő változásai eredményezik.
 |
A hazai erdőtársulások lehetnek klímazonálisak, intrazonálisak, ill. extrazonálisak.
Klímazonális erdők
Kialakulásukban a döntő környezeti tényező az éghajlat volt, elsődlegesen meghatározóak a hőmérséklet és csapadékviszonyok.
Az éghajlat magassági övei szerint kialakult egyes erdőtársulások:
A klímazonális erdőkben előfordulhatnak extrazonális, azaz zónán kívüli állományok. Hűvös hideg völgyekben, meredekebb északi hegyoldalakon a tölgyes és gyertyános-tölgyes erdőkben megjelenik az alacsonyabb hőmérsékletet kedvelő bükk.
Cseres-tölgyes
Gyertyános-tölgyes
Bükkös
55. tétel: Az életközösségek tér- és időbeli mintázatai |
A társulás – életközösség, biocönózis - egy adott helyen, adott időben együtt létező populációk életközössége. Stabil, önszabályozó rendszer.
A társulások térbeli mintázatai, a társulások szerkezete
A társulások szerkezetét a növénypopulációk határozzák meg. A társulások térbeli szerkezete tagolódhat
Függőleges szerkezet
A függőleges (vertikális) szerkezetet a növényeknek a fényért folyó versengése alakítja ki. Ez jelentkezik a társulás szintezettségében. Az egyes szinteken eltérőek az egyes környezeti tényezők értékei, pl. a fény mennyisége, hőmérséklet, páratartalom.
A szintezettség természetesen a vízi társulásokra is jellemző.
A gyökérszint
Az avarszint
A gyepszint
A cserjeszint
A lombkoronaszint
Vízszintes szerkezet
A társulás vízszintes szerkezetét, mintázatnak nevezzük.
A mintázatot a
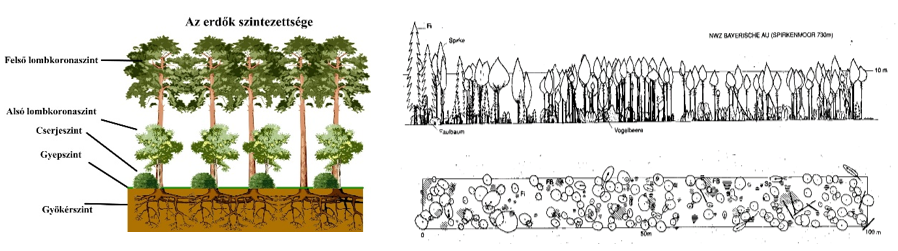 |
A társulások időbeli mintázatai, változásai
A társulások időbeli változásai lehetnek periodikusak, és lehetnek egy adott irányba haladók.
Periodikus – visszatérően ismétlődő - változások
A környezeti tényezők – fény, hőmérséklet, csapadék, páratartalom - periodikusan ismétlődő változásai eredményezik.
Napszakok szerinti változás
Évszakok szerinti változás
A társulások évszakok váltakozásával kapcsolatos, periodikusan ismétlődő állapotai az aszpektusok.
Egyszeri, előrehaladó változások: szukcessziók
A szukcesszió egy adott élőhelyen található életközösség átalakulási folyamata egy másik közösséggé.
Ismert
A természetes szukcessziónak alapvetően két típusát különböztetjük meg.
Szekuláris szukcesszió
Biotikus szukcesszió
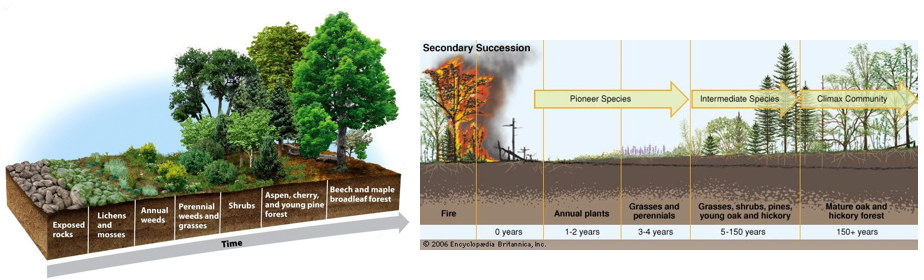 |
Példa a szukcesszióra, a sekély tavak feltöltődése
Időrendben a következő társulások jelennek meg egy sekély tó kialakulását követően.
A folyamat végére a tó feltöltődik, helyét egy puhafa-ligeterdő veszi át.
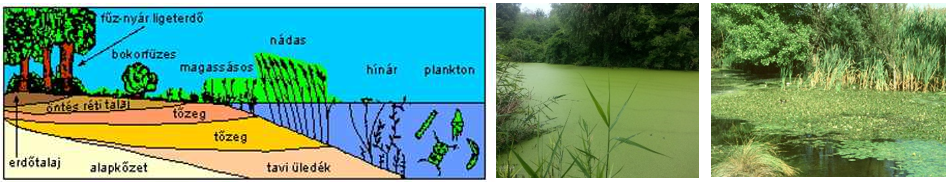 |
Az ember hatása a szukcesszióra
Az emberi tevékenység gyorsíthatja a szukcessziót. Ilyen pl. az eutrofizáció.
Eutrofizáció
Tápanyagoknak, különösen nitrogén- és/vagy foszforvegyületeknek a vízben való feldúsulása algák és magasabb rendű növényi életformák növekedésének felgyorsulását okozza, és ezzel nemkívánatos zavart eredményez a vízi élet egyensúlyi helyzetében, valamint a természetes vizek minőségében.
A szukcessziót lassító hatások
Degradáció
Különböző természetes rendszerek, társulások, biotópok állapotának leromlása. Okok:
56. tétel: A szén és az oxigén körforgása a természetben |
Anyagforgalom az ökoszisztémában, bio-geo-kémiai ciklusok
Az energia áramlása az anyagáramlástól az ökoszisztémákban elválaszthatatlan, hiszen az egyes táplálkozási szintek között az energia szerves anyagok formájában adódik tovább.
A két folyamat között azonban van egy alapvető különbség:
A körfolyamat lényege:
A szerves anyagokat
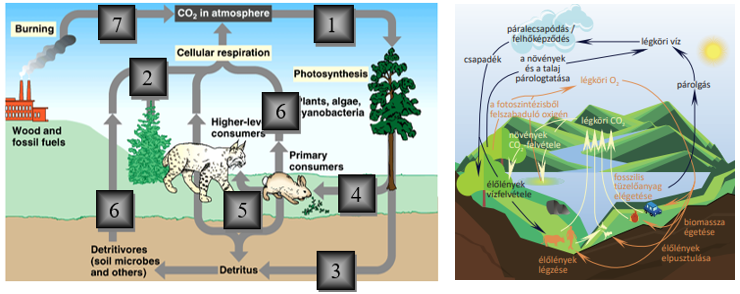 |
A szén körforgalma
Az oxigén körforgalma
A növények által felvett víz
57. tétel: A nitrogén körforgása a természetben |
A nitrogén körforgalma
A nitrogén nélkülözhetetlen alkotóeleme a fehérjéknek, nukleinsavaknak. Az elemi nitrogént kizárólag baktériumok és egyes gombák képesek közvetlenül a levegőből felhasználni. Az összes többi élőlény ezek tevékenységére van utalva.
Az előállított ammónia
 |
58. tétel: Táplálkozási hálózatok és piramisok |
Az ökoszisztémák négy összetevőből állnak:
Az ökoszisztéma elemeinek kapcsolata hozza létre
Táplálékláncok
A táplálékláncok egymással szoros táplálkozási kapcsolatban álló populációkból állnak, láncszemei az előtte levőből táplálkoznak és egyben táplálékul szolgálnak az utána következő láncszem fajainak.
1. Az anyag- és energiaáramlás első szintjét autotrófok – főleg növények - az ún. termelő vagy producens szervezetek képezik. Ezek fotoszintézissel, ill. kemoszintézissel szervetlen tápanyagokból (vízből és a levegő szén-dioxidjából) szerves anyagokat képesek előállítani, miáltal energiát építenek be szerves vegyületeikbe.
A többi fogyasztószervezet hasonló – autotróf – szerves anyag előállításra nem képes. Bennük a növények által előállított szerves anyagok áramlanak az új és új tápláléklánc-alkotók irányába.
2. A második szintjet a növényevő állatok, az elsődleges fogyasztók alkotják.
3. A további szinteken - másodlagos, ill. harmadlagos stb. fogyasztókban –, a ragadozó állatokban a szerves anyag egyre magasabb szintre kerül. A csúcsragadozónak az adott társulásban nincs természetes ellensége.
4. Az elhalt szerves anyagokat a lebontók vagy reducensek bontják le; ezzel a különböző anyagok (kémiai elemek) visszakerülnek szervetlen formáikba.
Az egyes populációk többféle táplálékláncnak is részét képezik, ezért helyesebb táplálékhálózatokról beszélni.
Táplálékhálózat
Egymással összekapcsolt táplálékláncok rendszere. Egy táplálékhálózatban egy meghatározott szervezet egynél több táplálkozási szinten is fogyaszthatja a táplálékait. Például
Minél bonyolultabb egy táplálékhálózat annál stabilabb a társulás.
 |
A táplálékláncok és -hálózatok szintjeit ennek megfelelően a
Az anyagok áramlása a táplálékhálózatokban egyirányú, vissza nem fordítható folyamat (a nyúl nem eszi meg a rókát, a fű meg a nyulat).
A peszticidek, más néven növényvédőszerek, olyan hatóanyagok melyek a kártevő élőlényeket – gombákat, gyomokat, állati kártevőket - távol tartják, terméketlenné teszik, ill. elpusztítják. A mérgező peszticideknek a helytelen alkalmazása
Még a távoli nemzeti parkok sem mentesek a peszticid szennyeződéstől, mivel a peszticideket a légáramlatok messze viszik - gyakran több ezer kilométerre - majd az esővel vagy hóval a talajra kerülnek. Ezt a jelenséget peszticid esőnek nevezzük.
A peszticideknek az emberek egészségére gyakorolt hatását két fő típusba lehet sorolni:
A táplálékláncokban az egyedek száma és össztömege a legalsó, termelői szinten a legnagyobb, és a legfölül elhelyezkedő csúcsragadozók szintjén a legkisebb. Az egymás után következő szintek csökkenő tömege, illetve egyedszáma alapján rajzolják meg az ún. táplálékpiramisokat.
Ez alól a parazitalánc kivétel, ahol a parazita szervezetek összegyedszáma meghaladhatja a tápláléklánc elején levő fogyasztó szervezetek egyedszámát.
 |
Biológiai produkció
Az ökológiai rendszerekben végbemenő szerves anyagtermelés és -átalakítás folyamata.
A biológiai produkció mértékegysége lehet pl. g/m²/év vagy kcal/m²/év.
A biológiai produkcióban termelődő szerves anyag sorsa:
Biomassza
A biológiai produkció hozza létre a biomasszát, amely
A biológiai produkció mértéke, a biomassza tömege mérhető, általában egy adott területegységre, meghatározott időre vonatkoztatva adják meg (g/dm3/év, g/m2/év, kg/ha/év stb.). Mivel a szerves anyagok energiatartalma kiszámítható, így a biológiai produkciót és a biomassza értéket energiatartalomban is meg lehet adni (lásd ábra). A különböző szintek biomasszája a termelők felől a csúcsragadozó felé egyre csökken, egy táplálékpiramisnak megfelelően.
Egy stabil, egyensúlyban levő társulás összes biomasszája közel állandó. A szukcesszió folyamatában az egymásután következő társulások biológiai produkciója és biomasszája egyre nő. A Földön a trópusi esőerdők biológiai produkciója és biomasszája a legnagyobb.
59. tétel: Energiaáramlás az ökoszisztémában |
Az ökoszisztémák nyílt anyagi rendszerek, a fennmaradásukhoz és a működésükhöz szükséges energiát kívülről kapják (napenergia). Az energia a különféle populációk élettevékenységeinek fenntartásához szükséges.
A külső forrásból származó energiát az autotróf (fototróf) vagy termelő szervezetek "hozzák be" a biocönózisokba, oly módon, hogy a napenergia felhasználásával – fotoszintézissel - saját energiadús szerves vegyületeiket építik fel energiaszegény szervetlen vegyületekből (szén-dioxidból és vízből).
Ezek a szerves anyagok haladnak végig a táplálékláncon, energiaforrásul szolgálva a heterotróf szervezetek, azaz a fogyasztók és lebontók számára.
A termelő szervezetek energiaforrásuk szerint fototrófok vagy kemotrófok lehetnek.
A mai termelő szervezetek túlnyomó többsége tehát zöld növény.
Az energiaforgalom szempontjából fontos a fotoszintetikus hatékonyság, az efficiencia is, amely a folyamat során felépített szerves vegyületek energiatartalma és a besugárzott energia hányadosa. Átlagosan a fotoautotróf szervezetek a rendelkezésükre álló sugárzó energiának alig kb. 0,5-1%-át építik be a szervezetükbe szerves anyagok formájában.
Egy fogyasztó által szerves vegyületek formájában felvett energia három útra terelődhet.
1. Beépül a fogyasztó testébe, annak testtömegét gyarapítja (~15%),
2. rögtön "elhasználódik", azaz lebomlik, eloxidálódik és a felszabaduló energia
3. ürülékkel vagy más salakanyagokkal együtt szerves vegyületekben kötött kémiai energia formájában távozik a szervezetből (~50%).
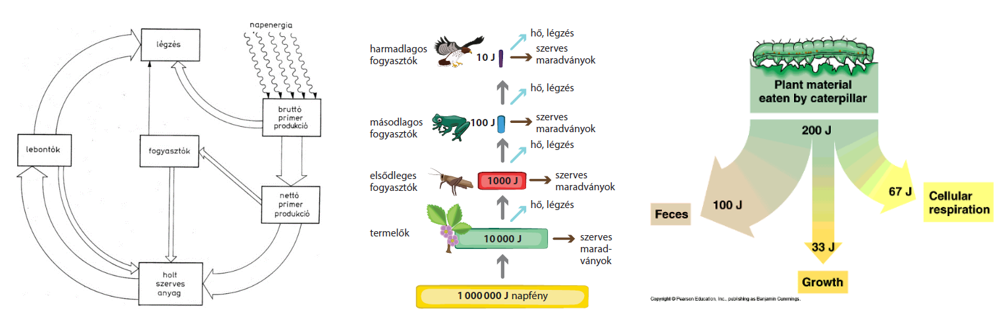 |
A fentiek szerint, az energia vándorlása során az egyes táplálkozási szinteken újabb és újabb energiaveszteséggel kell számolni (hő, salakanyag). A társulásba beépülő, majd szintről szintre szerves anyagként áramló energia tehát mind kevesebb lesz (lásd ábra).
A fény vagy kémiai energia formájában felvett energia minden esetben
Előbb vagy utóbb, a különböző szinteken, a szerves anyagok hőfelszabadulás közben elbomlanak és visszaalakulnak szervetlen anyagokká, melyek újra kiindulási anyagai lesznek az elsődleges biológiai produkciónak (autotróf asszimilációnak).
Az energia áramlása az anyagáramlástól az ökoszisztémákban elválaszthatatlan, hiszen az egyes táplálkozási szintek között az energia szerves anyagok formájában adódik tovább.
A két folyamat között azonban van egy alapvető különbség:
60. tétel: Környezetvédelem |
Globális folyamatok
Gaia-elmélet
Lényege, hogy a Föld összes élő és élettelen része egy szorosan összefüggő egységes és önszabályozó ökorendszert alkot, melyet egyetlen élő szervezetnek tekinthetünk.
A földi ökoszisztéma azért hasonlítható egy élő szervezethez, mert az élet egy olyan rendszernek a sajátossága,
Gaia tehát a Föld bioszféráját, atmoszféráját, vizeit és földjeit magába foglaló összetett rendszer, mely élőlénynek tekinthető, mert önszabályozó folyamatok a Föld sokféle jellemzőjét tartják állandó értéken, de legalábbis egy meghatározott tartományon belül.
Globális problémák
Földünkön az élő rendszereket az alábbi globális problémák fenyegetik, melyek hátterében bizonyítottan az emberiség környezetre gyakorolt káros hatásai állnak.
 |
Népességrobbanás, túlnépesedés
Az utóbbi 200 évben a népességnövekedés exponenciális tendenciát mutat.
A népességrobbanás okai:
A túlnépesedés áll a legtöbb globális probléma hátterében, gyökere tulajdonképpen az összes környezeti problémának, beleértve a környezetszennyezést, az élelmiszer- és energiagondokat, valamint nagyon sok gazdasági és társadalmi problémát.
Városiasodás
Az urbanizáció vagy városiasodás a városok térbeli és népességbeli növekedését, valamint a városi életforma terjedését jelenti.
A falvakból való elvándorlás okai a romló életfeltételek, a munkanélküliség és a biztonság hiánya.
A városiasodás társadalmi problémái
A városok ökológiai hatásai
Az úthálózatok ökológiai hatásai
Az autópályák, nagyobb autóutak
 |
Környezetvédelem
A környezet alatt
Az ökoszisztémát együttesen alkotják az élőlények, és az élettelen környezet (talaj, víz, levegő stb.).
Az épített környezetbe a települések, az utak, az infrastruktúra tartozik bele.
A környezet és természet nem szinonima fogalmak. A természet a környezet azon halmaza, amely nem tartalmazza az épített környezetet. Védelmük is különválik környezet-, valamint természetvédelemre.
Környezetvédelem
Környezetvédelmi Lexikon: "A környezetvédelem olyan céltudatos, szervezett, intézményesített társadalmi tevékenység, amelynek célja az ember ipari, mezőgazdasági, bányászati tevékenységéből fakadó káros következmények kiküszöbölése és megelőzése az élővilág és az ember károsodás nélküli fennmaradásának érdekében".
Természetvédelem
A természetvédelem a természeti területek értékeinek megőrzésére, bemutatására és helyreállítására irányuló társadalmi tevékenységek összessége.
Természeti terület az olyan terület, amely állapota természetközeli, nem érte jelentős emberi behatás.
Természeti értékek a természeti erőforrások, az élővilág és az élővilág fennmaradásához szükséges élettelen környezet.
A környezetszennyezés csökkentését ösztönző főbb gazdasági és jogi lehetőségek
Adók, tiltás, határérték, bírság, polgári per
A szabályozás lehet közvetlen és közvetett.
A közvetlen szabályozás adminisztratív tiltásokon, engedélyezési kötelezettségeken alapul, eszközei a kibocsátási engedélyek, környezetvédelmi normák, határértékek megállapítása.
A közvetett szabályozás gazdasági ösztönzőkkel tereli a piac szereplőit a kívánt környezettudatos magatartás irányába, pl. az adók, díjak bevezetésével, ill. környezetbarát technológiát alkalmazó támogatások rendszerével.
A kibocsátási díjakat a környezetbe juttatott szennyező anyag mennyisége alapján állapítják meg. A határértéket túllépő szennyezés esetén bírságot kell fizetni.
Környezetvédelmi bírság: aki jogszabályban foglalt, a környezet védelmét szolgáló előírást megszeg, vagy azokban megállapított határértéket túllép, az általa okozott környezetkárosítás mértékéhez igazodó környezetvédelmi bírságot köteles fizetni.
Egyes esetekben Hatályos Környezetvédelmi Törvény lehetővé teszi, hogy a környezetvédelmi, társadalmi szervezetek környezetveszélyeztetés, környezetszennyezés és környezetkárosítás esetén a környezethasználóval szemben polgári pert indítsanak a környezet védelme érdekében.
Határérték: A környezethasználat jogilag (egészségügyileg) elfogadható felső határa. A határértékek leggyakoribb fajtája a kibocsátási vagy emissziós határérték, melyet az érintett környezeti elem vonatkozásában a terhelő hatások, illetve szennyező anyagok fajtáira állapítanak meg (szennyvíznél pl. víz ólomtartalmára töménységben g/m3, zajhatásnál decibelben stb.).
Az intézkedések által elérni kívánt célok:
Környezetvédelmi adók
A környezet védelme érdekében az országok adórendszerébe be kell iktatni a környezeti adók tételét. Az adók alatt a környezetvédelmi adókat kell értenünk, mint például
Az ökoadót olyan termékekre, szolgáltatásokra (pl. olaj, gáz, áram), illetve olyan termelőtevékenységekre vetik ki, amelyek szennyezik a környezetet. Az adóztatás célja
61. tétel: Fenntarthatóság |
Fenntartható fejlődés
A fenntartható fejlődés olyan fejlődés, amely „kielégíti a jelen szükségleteit, anélkül, hogy veszélyeztetné a jövő nemzedékek esélyét arra, hogy ők is kielégíthessék szükségleteiket”.
A fenntartható fejlődés három alappilléren nyugszik:
Komplex megközelítéssel, egyszerre veszi figyelembe a természeti folyamatokat, a társadalmi igényeket, és a gazdasági fejlődés igényeit.
A fejlődés alapvető célja tehát a szociális jólét lehetőségének biztosítása, egyaránt a jelenlegi és a jövőbeli nemzedékek számára. Ez csak úgy lehetséges, ha közben fenntartható módon hasznosítjuk a természeti erőforrásokat, elkerüljük a környezet állapotában bekövetkező visszafordíthatatlan káros változásokat.
Alapelvek
Az ENSZ Fenntartható Fejlődés Bizottsága 1994-ben kidolgozta a fenntartható fejlődés mutatóit.
Az ökológiai lábnyom és a biokapacitás fogalma
Egy ember vagy egy adott terület népességének a természetre gyakorolt hatását, egy hektárban kifejezett mutatószámmal, az ökológiai lábnyommal lehet leírni.
Az ökológiai lábnyom az a terület, ami károsodás nélkül meg tudja termelni az életvitelünkhöz szükséges javakat (élelem, energia). Mértékegysége hektár/fő/év.
Kifejezi, hogy adott technológiai fejlettség mellett egy emberi társadalomnak milyen mennyiségű természeti javakra – pl. földre és vízre - van szüksége önmaga fenntartásához és a megtermelt hulladék elnyeléséhez. Ez az érték kiszámítható egyes emberekre, csoportokra, régiókra, országokra vagy vállalkozásokra is.
Az átlagos egy főre eső ökológiai lábnyom 2,2 hektár, 2,5-szer nagyobb, mint 1961-ben. Ám ha megnézzük, hogy a Földön 11,3 milliárd hektár biológiailag aktív föld- és tengerfelület van és 7 milliárd ember, akkor kiszámítható, hogy valójában minden emberre csak 1,6 hektár jut.
Ugyanakkor a biokapacitás megmutatja egy adott terület eltartóképességét, azon természeti erőforrások mennyiségét, amelyet a természet minden évben képes előállítani.
A biokapacitás alapján eldönthető, hogy valamely ország természeti erőforrásai elegendők-e az adott ország fogyasztási, illetve termelési tevékenységének a fenntartáshoz.
A biokapacitás és az ökológiai lábnyom különbsége azt a deficitet mutatja, amellyel lehetőségeinket túllépve más országokat vagy a jövő generációkat terheljük. Deficit esetén az ország ökológiailag fenntarthatatlan módon működik.
Magyarország ökológiai lábnyoma 2008-ban 3,59 gha volt. A fogyasztás ökológiai lábnyoma kb 0,9 gha-val haladja meg az ország biokapacitását, és kb. 2 Földre lenne szükség ahhoz, ha mindenki ugyanúgy akarna élni, ahogy mi Magyarországon.
 |
A természetvédelem
A természetvédelem a természeti területek értékeinek megőrzésére, bemutatására és helyreállítására irányuló társadalmi tevékenységek összessége.
Természeti terület az olyan terület, amely állapota természetközeli, nem érte jelentős emberi behatás.
Természeti értékek a természeti erőforrások, az élővilág és az élővilág fennmaradásához szükséges élettelen környezet.
A természetvédelem lehetőségei
A természetvédelmi tevékenység lehet passzív és aktív.
Ilyenek a kezeletlen védett területek, pl. az őserdők (olyan fás társulások, melyeket ember nem bolygatott meg). A védett területeken emberi beavatkozás nélkül történik a természetes megújulás és fenntartás.
A természetvédelem további lehetőségei
A természetvédelem alapvető eszköze – a fajok védelme mellett - a védett természeti területek létrehozása. Országos jelentőségű, egyedi jogszabállyal védett természeti területeknek nevezzük a természetvédelemért felelős miniszter által, rendeletben védetté nyilvánított természeti területeket. Ezek lehetnek
 |
Nemzeti parkok
A legmagasabb és legsokoldalúbb természetvédelmi kategória. Az ország legjelentősebb területeinek természetes állapotban tartása és bemutatása a cél.
Nemzeti park az ország jellegzetes, természeti adottságaiban lényegesen meg nem változtatott, olyan nagyobb kiterjedésű területe, melynek elsődleges rendeltetése a különleges jelentőségű, természetes növény- és állattani, földtani, víztani, tájképi és kultúrtörténeti értékek védelme, a biológiai sokféleség és természeti rendszerek zavartalan működésének fenntartása, az oktatás, a tudományos kutatás és a felüdülés elősegítése.
A nemzeti parkokat ütköző zónával kell körbevenni, amely megszűri a káros hatásokat, illetve ezeket, ha szükséges a park területén belül is ki kell alakítani.
A NP területén tudományos kutatómunka, oktatás és természetvédelmi nevelés folyik. Magyarországon jelenleg 10 nemzeti park található, melyek lefedik az ország egész területét. A nemzeti parkokban hagyományos gazdálkodási ágak maradtak fenn, vagy ma is hasznosítják a terület egy részét a gazdálkodásban.
Magyarország nemzeti parkjai
http://www.nemzetipark.gov.hu/
Pusztai jellegű nemzeti parkok:
Hegyvidéki jellegű nemzeti parkok:
Vizes jellegű nemzeti parkok:
A tájvédelmi körzet (TK) természeti, tájképi adottságokban gazdag nagyobb, általában összefüggő terület, tájrészlet, ahol a természet védelméről szóló 1996. évi LIII. törvény szerint „az ember és természet kölcsönhatása esztétikai, kulturális és természeti szempontból jól megkülönböztethető jelleget alakított ki, és elsődleges rendeltetése a tájképi és a természeti értékek megőrzése”.
A természetvédelmi terület (TT) az ország jellegzetes és különleges természeti értékekben gazdag, kisebb összefüggő területe, amelynek elsődleges rendeltetése egy vagy több természeti érték, illetve ezek összefüggő rendszerének a védelme.
Természetvédelem mellett szóló etikai, egészségügyi, kulturális és gazdasági érvek
Az etikai érv
A természet a belőle származó gazdasági érték mellett ún. belső értékkel rendelkezik, s mint olyan védelemre érdemes. Az élővilág egyedeinek joguk van az élethez, illetve minden fajnak joga van a túléléshez. Az élet egyszeri és megismételhetetlen.
A jóléti érv
A jóléti érv alapja a környezeti állapot romlása és az emberi egészség közötti összefüggés.
A rossz minőségű ivóvíz betegségek terjedéséhez vezethet, a rossz levegőminőség hozzájárul a légzési rendellenességek kialakulásához, a zaj pedig növeli a stresszt.
Az Európai Közösség Alapszerződésének 130. cikkelye szerint a környezetpolitika egyik fő célja az emberi egészség védelme.
A jóléti érv továbbá nem korlátozódik a jelenlegi generációra, hanem magában foglalja a jövő generációit is. A jövő generációival szembeni szolidaritás összhangban áll a fenntartható fejlődés célként való meghatározásával. Ez etikai érv is egyben.
Gazdasági érv
A környezetvédelmi szempontokat ma a gazdasági érvek uralják.
Nemzetközi egyezmények
62. tétel: A génműködés szabályozása |
Génműködés szabályozása a prokariótákban
A lac-operon
Differenciált génműködés folyamatát baktériumokban írták le először. A génműködés szabályozását leíró modell az operon modell. Az operon általi génműködés-szabályozás a sejtek gyors és hatékony alkalmazkodását teszi lehetővé a változó környezeti feltételekhez.
A baktériumok tápanyagok levesében úszva élnek. Sokféle különböző cukor fordulhat elő a környezetükben. Mindezek felvételéhez és lebontásához szükséges enzimek egyidejű termelése sok energiába kerülne. A baktériumok érzékelni képesek a rendelkezésre álló cukor típusát és csak akkor termelik a felvételhez és lebontáshoz szükséges enzimeket, ha szükség van azokra.
Az operon tehát a génműködés-szabályozás egysége. Megkülönböztetünk a DNS-molekulában
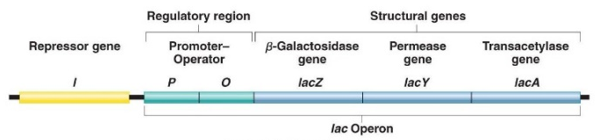 |
A lac-operon a következő elemekből áll:
Nem tartozik a lac operonhoz, - de alapvetően szükséges annak működéséhez -, az ún. lacI gén. A lacI gén kódolja a represszor fehérjét, amely tehát laktóz hiányában az operátorhoz kapcsolódva megakadályozza az RNS-polimeráz továbbhaladását, így az átírást.
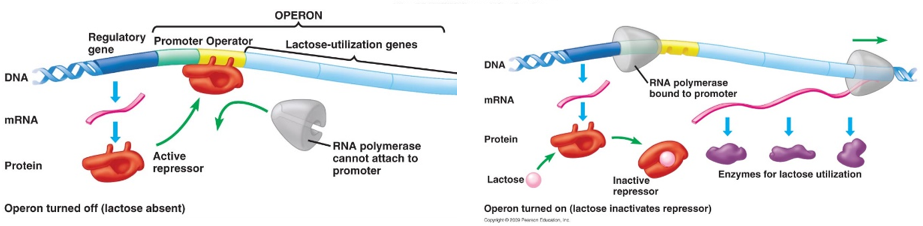 |
Laktóz hiányában
Laktóz jelenlétében
Video
https://drive.google.com/file/d/0B15KJwYa2uMFeGpaNW5lUVhZUlk/view?usp=sharing
63. tétel: Mutációk |
Mutáció
A mutáció az örökítő anyag (DNS) egyetlen nemzedéken belüli, ugrásszerű, öröklődő megváltozása. Óriási az evolúciós jelentősége, a populációk genetikai sokféleségének az alapja.
A genetikai változatosságot a mutáción kívül tovább növeli az ivaros szaporodás során bekövetkező, véletlenszerű tulajdonságkombinációk végtelen lehetősége:
A mutáció hatása lehet hátrányos (leggyakoribb), lehet közömbös, de lehet előnyös is.
Közömbös mutációról akkor beszélünk, ha a DNS megváltozását nem követi fehérjeszerkezeti változás. Erre példa,
Az evolúció során tehát az egyes allélváltozatok mutációval jöttek létre. Ma az élőlények az evolúció során felhalmozódott kedvező mutációkat hordozzák.
A mutáció mindig átadódik az utódsejtekre, de az utódokba (egyedekbe) csak az a mutáció öröklődik, amely az ivarsejteket (vagy az azokat képző sejteket) érinti. Ez a generatív mutáció (a klasszikus értelemben vett mutáció).
A különböző felépítésű és működésű testi sejtjeink genetikai információtartalma azonos, azonban a szomatikus mutációk, ami a testi sejtekben történik, ezt megváltoztathatják. A szomatikus mutáció nem öröklődő, mozaikos jellegű (pl. pigmenthiány a bőrön).
Szomatikus mutáció
A szomatikus mutációk tehát mozaikos fenotípust okoznak. A mutációt szenvedett testi sejtek utódai klónt alkotnak és gyakran a többi sejttől elkülöníthető mutáns szektorként jelennek meg. A mutáns szektor mérete annál nagyobb, minél korábban történt az egyedfejlődés során a mutáció.
Generatív, csíravonal mutáció
Az ivarsejtekben megjelenő öröklődő mutációkat csíravonal-mutációknak nevezzük. Egyes gének csíravonal-mutációi örökletes rákra hajlamosító tényezőt jelentenek. Ez a magyarázata a rákra való hajlam örökölhetőségének. A legismertebbek talán az öröklődő emlő- és petefészekrákkal összefüggő BRCA1 és BRCA2 gének mutációi (BRCA = breast cancer). Az emlőrákoknak kétféle típusa létezik, az öröklődő és a nem öröklődő emlőrák. A BRCA1 és a BRCA2 génekben előforduló mutációk növelik az öröklődő emlőrák kialakulásának kockázatát, mivel e két gén olyan fehérjéket állít elő, amelyek a DNS hibáinak kijavításáért felelősek. Mutáns gének esetén romlik a DNS javítómechanizmusainak hatékonysága, ezért nő a daganat kialakulásának a kockázata. Ha a BRCA1 és BRCA2 gének bármelyike mutáns formában kerül át az utódba, akkor már az átlagosnál jóval nagyobb eséllyel alakulhat ki a betegség. Az emlőrák kialakulásának általános kockázata 10%, ha azonban valaki mutáns BRCA-génnel rendelkezik, a valószínűség 80%-ra szökik fel.
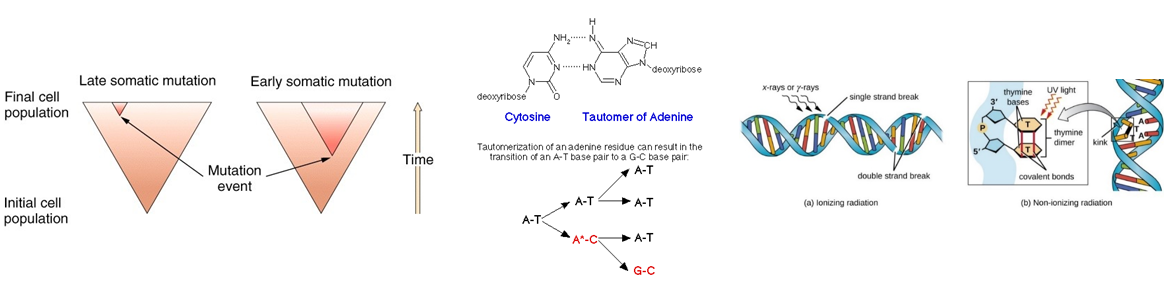 |
A mutációk okai
A mutáció során különféle okok miatt megváltozik a DNS-molekula nukleotidsorrendje.
Spontán mutáció az olyan mutáció, aminek a pontos kiváltó okát nem ismerjük.
Ilyenek a nemkívánatos kémiai reakciók, amelyek replikációs hibákhoz vezetnek. Bonyolult kémiai átalakulások okozzák. A változások hátterében az áll, hogy a különféle bázisok szerkezete könnyen átalakulhat olyan módon (tautomer átalakulás), hogy a változás következtében már a másik, egyébként nem komplementer bázissal képes kapcsolódni, pl. az adenin nem a timinnel, hanem a citozinnal. Így másoláskor már a timin helyére citozin kerül, újabb másoláskor pedig az eredeti adenin helyére guanin.
Indukált mutáció, melyeket ismert környezeti hatások (mutagének) váltanak ki. A mutagén olyan hatás vagy anyag, amely mutációt képes kiváltani, illetve a mutációk számát megnövelni a spontán mutációk gyakoriságához viszonyítva. Lehetnek:
Az UV sugárzás mutagén hatása
Az ultraibolya sugárzás a DNS-ben fotodimer termékeket eredményez, amelyek szomszédos timin bázisok kémiai összekapcsolódásával keletkeznek. Ezek a termékek torzulást eredményeznek a kettős spirál szerkezetében, és megakasztják a DNS-polimeráz haladását.
Kémiai mutagének
Például az alkiláló szerek (pl. a mustárgáz) úgy módosítják a bázisokat, hogy azok rosszul párosodnak.
Egyes gombákban termelődő aflatoxinok a DNS-hez kötődve megakadályozzák a hibátlan másolást, ill. pl. gátolják a daganatos sejtek kiküszöbölését végző immunválaszt.
Mutációk gyakorisága, a mutációs ráta
Az ivarsejtek képződésekor bekövetkező mutációk gyakoriságát fejezi ki. A mutációs ráta egy adott gén mutációinak gyakoriságát jelenti egy adott időegység alatt, általában generációnként. Pl. 10-3 érték azt jelenti, hogy 1000 ivarsejt képződésekor 1 mutáció történt.
A mutációs ráta
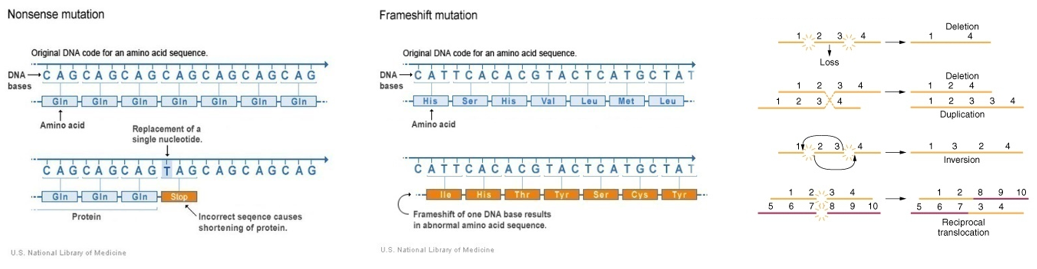 |
A mutációk típusai
Pontmutáció
Leggyakrabban a replikáció során következik be, többnyire egy nukleotidot érint. A mutagének legalább háromféleképpen indukálhatnak mutációt:
Kromoszómamutációk
Kromoszómamutációk azok a folyamatok, melyek
E változások többnyire mikroszkóppal is láthatók.
A kromoszóma szerkezeti változások meiózis során jönnek létre, ugyanis a folyamatban természetszerűleg törnek-egyesülnek a kromoszómák. Mutáció akkor keletkezik, ha a letört darab rendellenesen forr vissza.
Típusai
Számbeli kromoszóma-rendellenességek lásd még 6.2.1.
Egy faj alap-kromoszómaszáma a haploid kromoszómaszám (genom), ember esetén 23 kromoszóma.
Haploidok
Euploidok azok a szervezetek, melyek a haploid kromoszómakészlet egész számú többszörösét hordozzák (diploid (2x), triploid (3x), tetraploid (4x) stb.). Növények között fordul elő, hogy az ivarsejtek képződése során haploid helyett pl. diploid kromoszómaszerelvényű ivarsejtek jönnek létre. A diploid ivarsejtek összeolvadásakor tetraploid sejtek keletkeznek.
A triploidok jellegzetesen sterilek. A sterilitás oka, hogy a meiózis során páratlan számú kromoszómának kellene párba állni. Triploidokat diploid és tetraploid szülők keresztezésével állítanak elő. A termesztett banán triploid, nincs magja, mert nincs normális meiózisa.
Az aneuploidoknál néhány kromoszómával több vagy kevesebb van a genomban. A rendellenességek az ivarsejtek képződésekor a meiózis során a kromoszómák hibás szétválására vezethetők vissza. Ha meiózis első szakaszában a homológ kromoszómapárok tagjai nem válnak el egymástól, akkor a meiózis négy utódsejtje közül kettő az érintett kromoszómát nem tartalmazza, míg a másik kettő két példányban is tartalmazza. Az eggyel több kromoszóma - egyes gátló funkciójú enzimek magas szintje miatt - rendellenes állapothoz vezet.
A 21-es triszómia, a Down-szindróma főbb jellemzői:
A rendellenesség kialakulásának a valószínűsége az anya életkorával nő, hiszen a petesejtek osztódásukat már az embrionális időszakban megkezdik, melynek során az osztódó sejtek belépnek a meiózis I. főszakaszának a profázisába. A folyamat ezt követően megáll és csak a tüszőérés során fejeződik be, így a fejlődő petesejtekben a kromoszómák akár évtizedeket is összetapadva tölthetnek és emiatt nehezebben válnak szét.
Néhány, mutációra visszavezethető rendellenesség, betegség lásd 65. tétel
64. tétel: Sejten belüli információáramlás |
Az élővilágban a genetikai információ tárolója kivétel nélkül a DNS-molekula, ugyanakkor ismertek vírusok - retrovírusok, mint pl. a HIV, influenzavírusok, COVID19 (coronavirus disease - koronavírus okozta megbetegedés 2019) stb. - melyeknek az örökítő anyaga RNS. A nukleinsavakban az információkat gének hordozzák. Génnek nevezzük a DNS-molekula adott szakaszát, amely valamilyen öröklődő jelleg kialakulását megszabja egy adott fehérje kódolásán keresztül.
A genetikai információt tehát a DNS-molekula hordozza. Az információ továbbításáért pedig az mRNS-molekulák felelősek.A különböző tulajdonságokért specifikus fehérjék felelősek. Specifitásuk az aminosavsorrendjükben rejlik. A DNS a fehérjék aminosavsorrendjét nukleotidsorrend formájában kódolja, melyben az egyes aminosavaknak nukleotidhármasok, ún. tripletek feleltethetők meg.
A sejten belül az információáramlás a következő úton halad (centrális dogma):
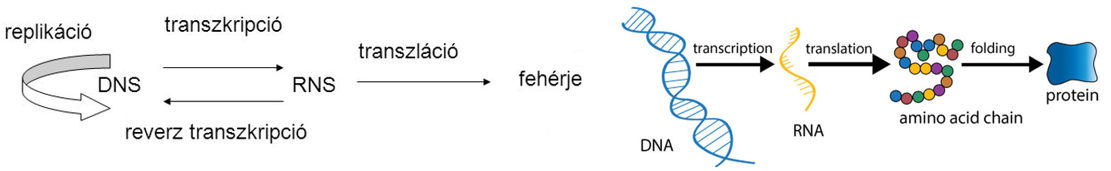 |
A DNS megkettőződése, a replikáció
A sejtek osztódásához szükséges a genetikai információ pontos megkettőződése és átadása az utódsejteknek, ez biztosítja ugyanis a biológiai információ lényegében változatlan megőrzését az utódnemzedékben. A DNS megkettőződés során a DNS-molekula pontos másolata jön létre.
A DNS-molekulában észlelhető szabályosság, a bázisok megfelelő kapcsolódása (komplementaritása) képezi az alapját a genetikai információ egyértelműen pontos másolásának, a replikációnak (adeninnel szemben mindig timin van, citozinnal szemben pedig guanin helyezkedik el).
A replikáció alatt a DNS-molekula két szála elválik egymástól és az így különváló láncok mintaként szolgálnak egy-egy újabb DNS-szál szintézisének.
Ennek megfelelően az utód DNS-molekulák egyik lánca a szülői DNS-ből származik, a másik komplementer lánc újonnan szintetizálódik hozzá (szemikonzervatív – félig megmaradó - jelleg).
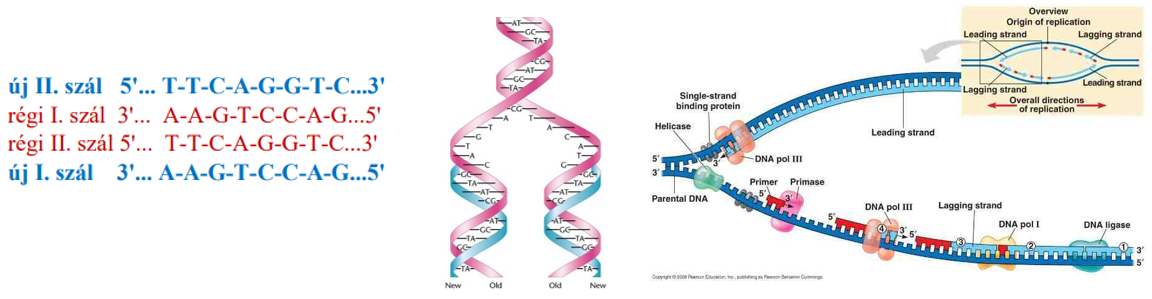 |
A replikáció mechanizmusa
A replikáció a DNS-szál szigorúan meghatározott részén, az ún. kezdő – iniciációs - ponton indul meg, amit replikációs origónak nevezünk.
1. A másolást megelőzően a kromoszómaszakaszhoz kicsavarófehérjék – helikázok - kapcsolódnak. A folyamatban résztvevő enzimek felszakítják a bázisok közötti H-kötéseket, aminek következtében a DNS-szálak elválnak egymástól.
A szabaddá vált DNS-szakaszon egy RNS-polimeráz (primáz) segítségével megindul a DNS-száléval komplementer, 50-100 ribonukleotid egységből RNS-primer szintézise.
2. A primer RNS 3'-OH végéhez a DNS-polimeráz III lépésenként dezoxiribonukleotid-trifoszfátokat A folyamat energiaigényét a lehasadó difoszfátok hidrolízise biztosítja.
Az új szál tehát 5' -) 3' irányba nő. Ahogy az új szál hosszabbodik, a kicsavaró fehérjék elválasztják a komplementer szálakat, kialakítják a replikációs villa újabb szakaszait.
Mivel a DNS két szála ellentétes polaritású, úgy szintézisük is ellentétes irányba történik, hiszen a szintézis iránya mindig 5’ -) 3’.
Minthogy a kicsavarófehérjék csak az egyik irányba haladnak, azaz a replikációs villa csak az egyik irányba nyílik szét, csak az egyik szál szintézise lehet folyamatos, az, amelynek növekedési iránya megegyezik a helikázok haladási irányával. Ebből következik, hogy a másik ellenkező irányba növő szál szintézise csak szakaszosan mehet végbe (Okazaki-fragmentek).
Az ellenkező irányba növekvő szálon a szintézis 1000-2000 RNS nukleotid egység összekapcsolásáig tart. Az RNS- primert a polimeráz I távolítja el, majd helyére komplementer dezoxiribonukleotidokat épít (kitölti a hézagokat).
3. Az egyes DNS-szakaszokat a ligáz kapcsolja össze úgy, hogy két nukleotid egység között foszfodiészter kötés kialakulását katalizálja.
https://drive.google.com/file/d/0B15KJwYa2uMFeDJsQjJQZHdJUEk/view?usp=sharing
A replikáció pontossága
A tulajdonságok megfelelő öröklődése szempontjából rendkívül fontos, hogy az örökítőanyagot a sejtek pontosan lemásolva adják át egymásnak. Ennek biztosítéka egyrészt a másolás pontossága, másrészt a replikáció során történt hibák kijavítása. Az emberi genom – haploid sejtenkénti DNS állomány – 3 milliárd bázispárból épül fel, replikációjakor átlagosan 6 hiba történik. A replikáció ilyen pontosságáért javítóenzimek felelősek. Az élő rendszerekben a hiba típusától függően a hibajavító mechanizmusoknak számtalan formája alakult ki.
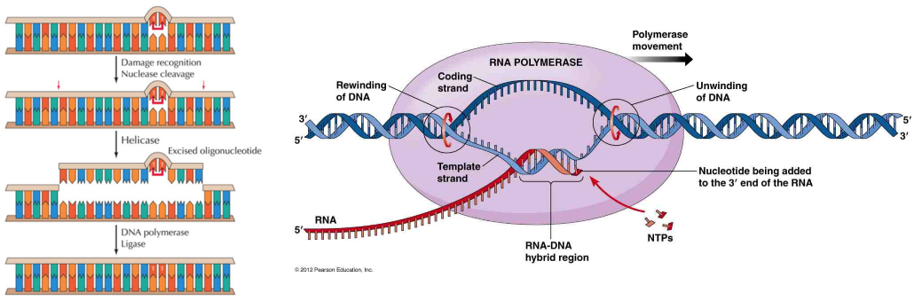 |
Transzkripció - átírás
Az élő szervezetek bármilyen életjelensége - anyagcsere, mozgás stb. - fehérjék működésén keresztül valósul meg. A különböző funkciókért specifikus fehérjék felelősek. Specifitásuk az aminosavsorrendjükben rejlik. Egy fehérje aminosavsorrend információját a DNS-molekula adott szakasza, az ún. struktúrgén hordozza.
Eukariótákban az információt hordozó DNS a sejtmagban helyezkedik el, ugyanakkor a fehérjék szintézise a citoplazmában folyik. A DNS tehát közvetlenül nem szolgálhat mintaként a fehérjék szintéziséhez. Ebből az következik, hogy a DNS az információt egy közvetítő molekulának adja át, amely kijutva a citoplazmába irányítja a fehérjék szintézisét. Ezt a szerepet a hírvivő, vagyis a messenger RNS (mRNS) látja el.
A DNS információjának RNS-molekulára átmásolását transzkripciónak nevezzük. E folyamattal nemcsak az mRNS szintetizálódik, hanem az összes többi RNS fajta is (tRNS, rRNS, miRNS).
Az információtároló DNS és az információt közvetítő RNS igen hasonló, közöttük a kémiai különbség csekély:
Az átírás – transzkripció - mechanizmusa
Az RNS-ek szintézisét DNS-mintaszálon az RNS-polimeráz enzim végzi úgy, hogy az RNS nukleotidokat lépésenként 5’ -) 3’ irányban összekapcsolja a mintául szolgáló DNS bázissorrendjének megfelelően. A replikációhoz hasonlóan az RNS-polimeráz nukleotidtrifoszfátokat kapcsol egymáshoz, miközben egy difoszfátot hidrolizál le, s az ennek során felszabaduló energia fedezi a szintézis energiaigényét.
Az RNS-polimeráz felismeri és hozzákötődik az átírás kezdőpontjához a promoterhez. A kapcsolódáskor a DNS két szála elválik egymástól, majd az értelmes – aktív szál, mintaszál, átíródószál - szálon megindul az átírás a bázisok párképződési szabályainak megfelelően, A = U, G = C.
A DNS kettős hélixnek csak az egyik szála íródik át (értelmes szál), a másik soha (néma szál). Az értelmes szál DNS-szakaszonként – génenként - változhat, de egy adott génre mindig állandó. A szintézis addig tart, amíg az RNS-polimeráz eléri a szintézis befejezését jelentő stopjelet.
A lánc végén egy U-ban gazdag szekvencia található, amely előtt egy hajtűhurok kialakulva megakasztja az enzimet, miáltal az átírás befejeződik.
https://drive.google.com/file/d/0B15KJwYa2uMFNlE1TnhvYXhUNzA/view?usp=sharing
https://drive.google.com/file/d/0B15KJwYa2uMFNUJXRUI2NEhHejg/view?usp=sharing
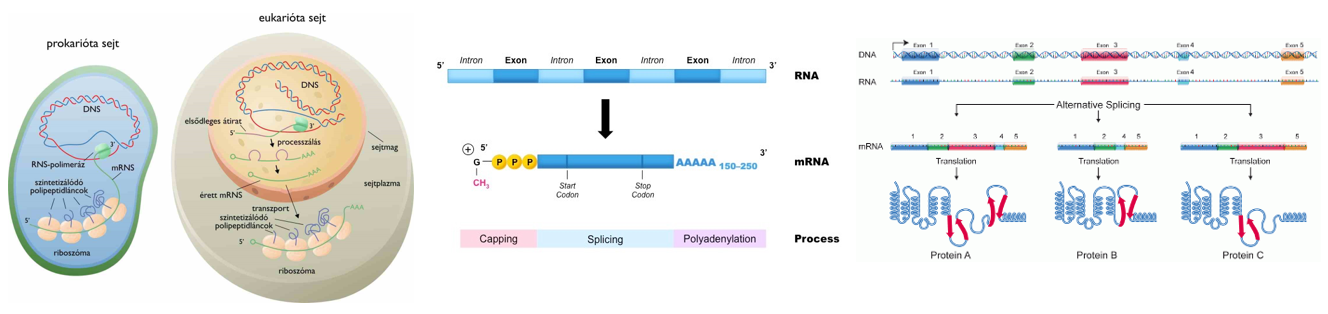 |
A prokariótáknál a transzkripció és a fehérjeszintézis csaknem szimultán folyamat, az elkészült mRNS 5’ vége még jóval a lánc szintézisének a befejezése előtt hozzákapcsolódik riboszómákhoz, s ezen a szakaszon már megkezdődik a fehérjeszintézis.
Eukariótáknál az RNS-polimeráz hatására kialakuló mRNS még nem teljes értékű végtermék, biológiai feladatának betöltése érdekében többféle átalakuláson, ún. érésen kell keresztülmennie, mielőtt kikerül a citoplazmába.
Az RNS érési folyamatai eukariótáknál (poszttranszkripciós módosítások)
1. Az mRNS két végéhez különböző szerkezeti elemek – sapka, poli-A farok - kapcsolódnak.
2. Az elsődlegesen elkészült mRNS-ből különböző méretű szakaszok, ún. intronok hasadnak ki. Ez a splicing.
1. Láncvédő szerkezeti elemek
A citoplazmába kikerülő mRNS-t nukleázok támadják, melyek a láncvégeken kezdik bontani a molekulát.
2. Az mRNS hasítása - splicing.
Sőt ezen túlmenően a gének tartalmaznak
Az exonokat egymástól 10-10000 nukleotidpárból álló intron szakaszok választják el.
A gén átírása folyamatos, a keletkező elsődleges átiratból egymást követő lépésekben hasadnak ki az üzenetet nem tartalmazó intron szakaszok és a szabadon maradt exonvégek egymással összeillesztődnek. A folyamatot splicingnak nevezik.
Az emberi gének száma mindössze kétszerese az ecetmuslicában meghatározottaknak. A komplexitás titka nem a gének számában, hanem a gének által meghatározott fehérjék összerakási módjában van. Az alternatív splicing mechanizmusa lehetőséget ad arra, hogy egy gén többféle fehérjét is meghatározzon. Egy gén kifejeződésekor nem minden exonszakasz marad benne a végső mRNS-ben, így annak megfelelően, hogy mely exonok illesztődnek össze, egyetlen gén többféle fehérjét is kódolhat.
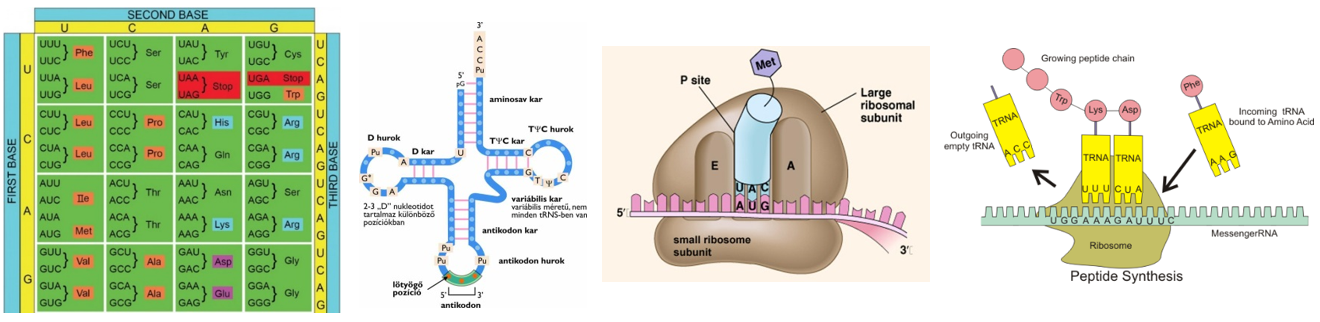 |
A genetikai információ kifejeződése, a fehérjék bioszintézise (transzláció, átfordítás)
A genetikai információt a DNS-molekula hordozza. Az információ továbbításáért az mRNS-molekulák felelősek. A kérdés az, hogy a DNS-ben nukleotidsorrend (bázissorrend) formájában foglalt információ hogyan határozza meg a fehérjemolekulák aminosavsorrendjét.
A választ a genetikai kód adja meg, amely egy olyan szabályrendszer, kódolási eljárás (nem anyagi egység), ami szerint a nukleinsavakban szereplő szöveg (nukleotidsorrend) lefordítható a fehérjékben szereplő szövegre (aminosavsorrend). Ennek a szabályrendszernek a feltárása a biológiai kutatások körében az egyik legnagyobb intellektuális teljesítmény volt.
A genetikai kód
A nukleinsavak ABC-je 4 betűs (DNS-ben pl. A,T,G,C), a fehérjéké 20 (aminosavak).
A genetikai információ egységeit, tehát DNS- vagy RNS-nukleotid hármasok ún. tripletek alkotják. Ezeket kodonoknak nevezzük. A kodonok jelentését a kodonszótár tartalmazza.
A genetikai kód jellemzői
1. Degenerált (redundáns), azaz egy aminosavat több triplet határoz meg, hiszen 20 aminosav beépülésére 64 kodon áll rendelkezésre.
A leggyakoribb aminosavakat 4-6 kodon is jelöli, pl. az Arg-nak, Ser-nek, Leu-nak 6,
2. Univerzális, mivel a genetikai kód a szervezetek fejlettségi szintjétől függetlenül egységes, minden élőlényre igaz (kivéve a mitokondriális DNS-t).
3. Átfedés- és kihagyásmentes, azaz egyetlen nukleotid sem lehet tagja két szomszédos kodonnak, ill. minden nukleotid tagja valamelyik tripletnek.
Ha egy vagy két nukleotid kiesik vagy beékelődik, az eltolja a leolvasási keretet, és ezzel értelmetlen aminosavsorrendet hoz létre, ami teljesen működésképtelen fehérjét eredményez (leolvasási kereteltolódás, frame shift mutáció).
A fehérjeszintézis, a transzláció – átfordítás, leolvasás - mechanizmusa
Az mRNS kodonjai közvetlenül semmilyen módon nem hozhatók kapcsolatba az általuk kódolt aminosavakkal, mivel szerkezetük nem komplementer. Ezért az aminosavaknak először egy olyan közvetítőmolekulához kell kapcsolódni, amely valamilyen módon kapcsolódni képes az mRNS kodonjaival.
A közvetítőmolekula a tRNS, amelynek feladata tehát kettős:
A tRNS szerkezete
A tRNS-ek a ribonukleinsavak között a legkisebb molekulák, mindössze 75-90 nukleotidból épülnek fel. Egy tRNS-molekulának specifikus háromdimenziós szerkezete van, amely biztosítja a maximális stabilitást. A molekula térbeli szerkezete lóherelevélre emlékeztet.
A molekulának a legfontosabb része az mRNS kodon felismerését biztosító, ún. antikodon rész. Az antikodon lehetővé teszi, hogy az adott tRNS-molekula, amely adott aminosavat szállít, a neki megfelelő mRNS kodonhoz kapcsolódjon.
Az mRNS kodon és a tRNS antikodon komplementaritása a biztosítéka annak, hogy adott mRNS-bázisszekvenciáról mindig azonos aminosav-szekvencia keletkezzék.
A riboszómák
A fehérjeszintézis helyszínei, rRNS-ekből és fehérjékből épülnek fel. Prokarióta sejtekben ezernyi, eukarióta sejtekben sok ezernyi található a citoplazmában, ill. előfordulnak egyes sejtszervecskékben, mint pl. mitokondriumokban, színtestekben. Működésük, hogy az mRNS nukleotidsorrendjében foglalt információt fordítják le aminosavsorrendre.
Szerkezetük egységes az egész élővilágban, egy nagyobb és egy kisebb alegységből állnak.
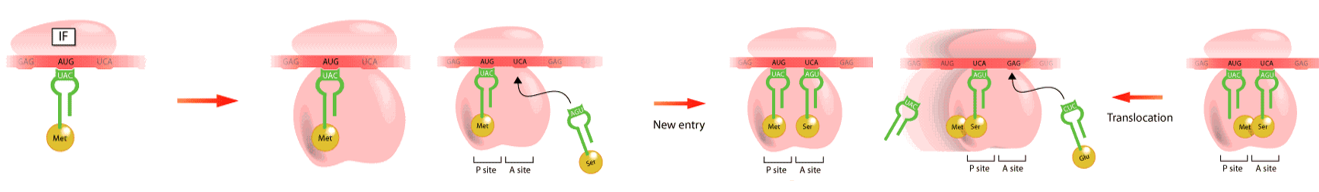 |
A transzláció lépései prokariótákban röviden
1. A start kodonhoz hozzákapcsolódik az első aminosavat – metionint- szállító tRNS.
2. A következő lépésekben a kodon felismerése révén a megfelelő aminosav-tRNS-ek egymást követően a riboszómához kapcsolódnak.
3. Az aminosavak között kondenzációval kialakul a peptidkötés.
4. A riboszóma lépésenként odébb mozdul az újabb tRNS megkötésének érdekében.
5. Ha a riboszóma elérkezik a stop kodonhoz, a transzláció befejeződik. A terminációnak az az oka, hogy a három stop kodonhoz nem tartozik aminosav, mivel a kodonoknak megfelelő komplementer antikodont tartalmazó tRNS nincs.
6. A stopjel felismerését követően a polipeptidlánc leválik a riboszómáról.
65. tétel: Öröklődő betegségek |
Példák autoszomális – testi kromoszómához kötött - domináns-recesszív öröklésmenetre
Monogénes autoszomális recesszív öröklésmenetek jellemzői (AR)
Recesszíven öröklődő autoszómális génmutációk
A fenilketonúria egy öröklődő, enzimhiányon alapuló szervezetszintű anyagcserezavar. A betegség a vérben és vizeletben felszaporodó aminosavról, a fenilalaninról ismerhető fel, amelyet újszülöttkorban vérminta útján vizsgálnak.
A fenilalanin az esszenciális aminosavak közé tartozik. A fenilalanin egy részét az egészséges szervezet májban termelődő egyik enzimje egy másik aminosavvá, tirozinná alakítja.
A fenilketonúriások szervezete - a májban az enzim hiánya vagy csökkent mértékű működése miatt – nem tudja átalakítani a fenilalanint, ezért az felszaporodik a vérben, ill. alternatív anyagcsereutak révén mérgező anyagcseretermékek szabadulnak fel.
A lebontatlan, így fölös mennyiségben jelenlévő fenilalanin a központi idegrendszer bizonyos területein fölhalmozódva idegrendszeri károsodást okoz, valamint az agy fejlődését károsan befolyásolja, kisfejűség alakul ki. Az értelmi fogyatékosságot okozó agykárosodás szigorú fenilalanin-mentes diéta nélkül az ötödik életév végére alakul ki.
Kezelése fehérjeszegény diétával történik. A fenilketonúriásnak tilos húsféléket, halat, tejtermékeket, gabonából készült termékeket, hüvelyeseket, tojást, szóját enni.
Albinizmus lényege, hogy a szervezetben a melanin termelődése csökken vagy teljesen hiányzik. Az 1-es típusú albinizmus esetén a tirozináz enzim hiánya miatt a tirozin aminosav nem tud melaninná alakulni, emiatt a pigmentáció is hiányos a bőrben, szőrzetben, hajzatban, továbbá a szivárványhártyában, aminek köszönhetően a szem színe a benne futó hajszálerekben keringő vértől piros színű.
Sarlósejtes anémiában a vörösvértestek kóros hemoglobint tartalmaznak, melynek következtében egyes vörösvértestek sarló alakúvá válnak. Mivel a károsodott sejtek merevek, a kis vérereken való áthaladáskor szétesnek, vérszegénységet, vérkeringési zavart és csökkent oxigénellátottságot okozva. A deformált sejtek rögökké tapadnak össze, melyek a lépet, a veséket, az agyat, a csontokat és a többi szervet is károsítják.
A rendellenesség hátterében egy, a hemoglobin β-lánc génjében bekövetkezett pontmutáció áll, aminek következtében a hemoglobinmolekulák összecsapódnak, kikristályosodnak, oxigénkötő képességük jelentősen csökken.
A tünetek homozigóta egyéneknél súlyosak, hiszen bennük kizárólag hibás hemoglobin található. A heterozigóták esetében a hemoglobinmolekulák egy része normális, így az összecsapódási hajlam is kisebb. Náluk csak oxigénhiány miatt jelentkezhetnek a tünetek, pl. magas hegyi tartózkodás, vagy erősebb fizikai terhelés esetén.
A sarlósejtes anémia rendkívül súlyos betegség, homozigóta recesszíveknél korai elhalálozást okoz (letális mutáció).
Ugyanakkor ismertek olyan speciális esetek, amikor a recesszív allél homozigóta formában letális, azonban heterozigótákban szelekciós előnyt biztosít a hordozójának. A jeleséget heterozigóta fölénynek nevezzük. Erre példa a sarlósejtes vérszegénységre nézve heterozigóták gyakori előfordulása Afrikának azon területein, ahol a malária pusztít.
A cisztás fibrózist a CFTR gén mutációja okozza, a gén egy fehérjét kódol, aminek a légúti és az emésztőrendszeri nyák megfelelő viszkozitásának kialakításában van szerepe. Mutációja esetén a nyák sűrű lesz, ami többek között helyi fertőzésre való hajlamot, a hasnyálmirigy elégtelen működését, a légzőrendszer krónikus gyulladását okozza.
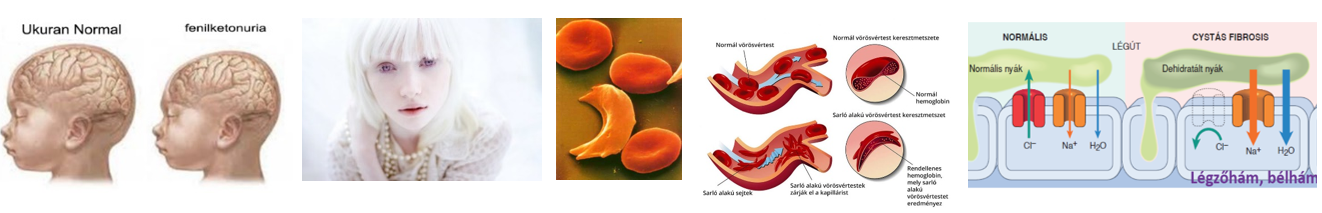 |
Monogénes autoszomális domináns öröklésmenetek jellemzői (AD)
Dominánsan öröklődő autoszomális génmutációk
Az achondroplasia a törpenövés egyik leggyakoribb fajtája. A betegség örökletes, egyetlen gén hibája okozza. Az érintett személyek heterozigóták, a homozigóta újszülöttek többsége légzési elégtelenség következtében korán meghal. Ha két átlagos termetű szülőnek születik achondroplasiás gyermeke, ennek az az oka, hogy valamelyik szülő ivarsejtjében mutáció következett be. A mutáció oka ismeretlen.
A rendellenesség főbb jellemzői.
 |
Példák gonoszomális – ivari kromoszómához kötött - domináns-recesszív öröklésmenetre
Az X-kromoszómához kötött recesszív öröklésmenet jellemzői (XR)
X-kromoszómához kötődő recesszív rendellenességek
A színtévesztő ideghártyájában az egyik csap nem működik megfelelően. Valamely színtartományban nem kellően érzékeny, vagy a színtartomány eltolódik valamelyik másik csap színtartománya felé. Leggyakoribb a zöld és a vörös receptorok hibája, a vörös csapocska színtartománya a zöld felé tolódik.
A hemofília ritka, veleszületett, egész életen át tartó vérzékenység, mely az egyes véralvadási faktorok rendellenes képződésével vagy hiányával kapcsolatos. A betegség lényege a vér alvadékonyságának csökkenése, melynek következtében a hemofíliás betegek sérülések alkalmával hosszabb ideig véreznek, mint az egészségesek.
A leggyakoribbak és legsúlyosabb problémát az ízületi- és izomvérzések jelentik. A belső vérzések által okozott feszítő nyomás miatt a hemofíliát az egyik legfájdalmasabb betegségnek tekinti az orvostudomány. A gyakori bevérzések következtében a betegek ízületei fokozatosan tönkremennek és izmaik sorvadnak.
Az alvadási faktorok mindegyikének leírták örökletes hiányállapotát. Leggyakoribb a
X-kromoszómához kötődő domináns rendellenességek (XD)
Y-kromoszómához kötött rendellenességek
A fül túlzott szőrössége az egyetlen férfiról férfira átadódó testi jelleg, a szőrös fül nem Y-hoz kötött, de génje és annak helye még nem ismert (Tóth Sára Genetika és Genomika).
Az Y-kromoszóma legnagyobb része nem hordoz információt, így mindössze néhány rendellenesség – pl. a férfi meddőség egyes formái - társulnak specifikusan ehhez a kromoszómához.
Az Y-kromoszómához kötött öröklődés jellemzői
Kizárólag férfiak érintettek.
66. tétel: Sejtciklus zavarai |
A folyamatosan osztódó sejtek esetében az osztódás és az osztódások közötti időszakok szabályosan követik egymást. Ebben az esetben sejtciklusról beszélünk. Ez jellemző az embrionális sejtekre, ill. az állandóan osztódó sejtekből álló szövetekre, mint pl. vörös csontvelő, bélhám, többrétegű elszarusodó hám, ivarszervek csírahámjai, továbbá ilyenek a tumorsejtek.
A sejtciklus alapvetően két részre osztható:
Az interfázis
Az interfázis alatt a sejtben számos olyan folyamat zajlik, amely a sejtosztódást készíti elő. Az interfázist három szakaszra osztjuk:
A G1 szakasz az osztódást közvetlen követi. A létrejött utódsejtek:
Azok a sejtek, amelyek már nem osztódnak tovább, a G1 szakaszból a G0 szakaszba kerülnek, ahol a sejt determinálódik és valamelyik differenciálódási pályán indul el. Maga a determináció, az elköteleződés azt a pillanatot jelenti, amelyben az illető sejt egy specifikus fejlődési pályára lép. Ez egy adott genetikai program beindulását jelenti.
A differenciálódás a sejtek sokféleségének az alapja, amelynek során az eredetileg egyforma sejtek más és más speciális képességekre tesznek szert.
Az S fázisban történik:
A kromoszómák az interfázisban is léteznek, csak ilyenkor a laza kromatinállományt alkotják, melyben a kromoszómák csak egyik kromatidája van jelen. Az S fázisban az egykromatidás kromoszómák DNS-e megduplázódik, és a keletkező másolat a kromoszóma másik kromatidáját alkotja. A kétkromatidás kromoszómákban tehát a két kromatida – a másolási hibáktól eltekintve – teljesen azonos információtartalmú.
A G2 fázis
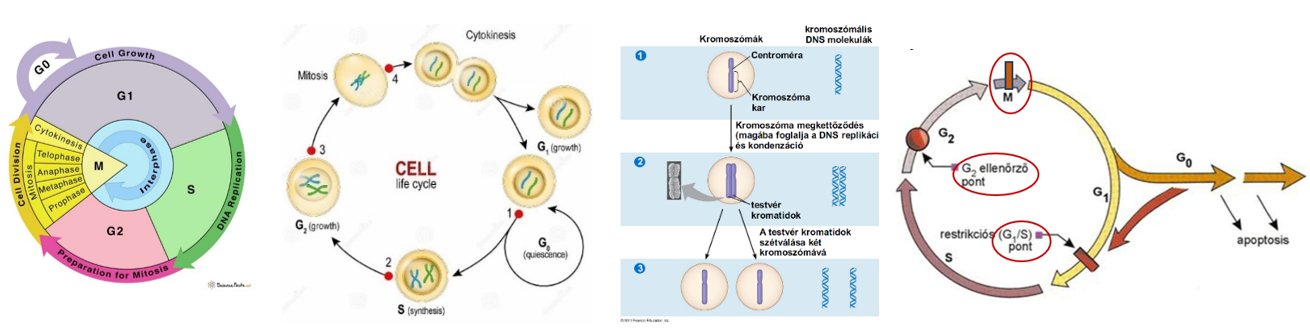 |
A sejtciklus szabályozása
Az eukarióta sejtek osztódási ciklusát szigorúan szabályozott folyamatok sorozata jellemzi. E folyamatok visszafordíthatatlan változásokat eredményeznek a ciklusban lévő sejtekben.
A soksejtűeknél a ciklus beindításához nélkülözhetetlenek az ún. növekedési faktorok, melyek jelátviteli útvonalak közvetítésével aktiválják az osztódáshoz szükséges géneket.
A ciklus meghatározott pontjain ellenőrző pontok működnek, melyek a részfolyamatok pontosságáért, ill. meghatározott sorrendjéért felelősek. Mutációs eredetű hibák a sejtciklus szabályozásának felborulását eredményezhetik.
Alapvető fontosságú, hogy a sejtciklus egyes fázisai a megfelelő sorrendben kövessék egymást, s a sejt csak akkor tudjon átlépni egy következőbe, ha az előzőt tökéletesen befejezte. Ellenkező esetben a genetikai anyag eloszlásában zavarok léphetnek fel. Elveszhetnek kromoszómák vagy kromoszómarészletek, vagy egyenlőtlenül osztódnak szét az utódsejtek között. A tumorsejteknél gyakran jellemzők ilyen elváltozások.
Ezeket a problémákat a sejtciklus ellenőrzési pontjai hivatottak kiküszöbölni azáltal, hogy leállítják a sejtciklust addig, amíg a hibák kijavításra kerülnek, vagy ha ez nem lehetséges, programozott sejthalálra, apoptózisra késztetik a sejtet. Ha azonban egyik sem következik be, akkor rákos elfajulás jöhet létre: a sejt irányíthatatlan módon osztódni kezd. Az eredeti hibás sejtből egy tumoros sejtvonal alakul ki, amely halhatatlanná válik.
Az első ellenőrzési pont, a G1 fázisból a szintézisbe (S) való átmenet a legfontosabb, hiszen ez az, amelyik a sejtciklusba való belépést, illetve a sejtosztódás megkezdését szabályozza (restrikciós pont).
Az átlépés feltétele rendszerint az osztódást serkentő jelmolekulák, az úgynevezett növekedési faktorok felszaporodása a sejt környezetében.
A második ellenőrzési pont a G2/M átmenetet felügyeli. Itt kell a sejtnek meggyőződnie arról, hogy a DNS másolása megfelelően zajlott-e le. DNS károsodás esetén a p53 tumorszupresszor fehérje felfüggeszti a sejtciklust.
A harmadik, utolsó ellenőrzési pont az M pont, ellenőrzi, hogy a kromoszómák mindkét kromatidája hozzákapcsolódott megfelelően a magorsóhoz.
A rák kialakulása és a sejtciklus zavarai
A rák kialakulását minden esetben az örökítőanyagot érintő mutációk felhalmozódása okozza. Azok a mutációk okoznak daganatot, amelyek a sejtciklust szabályozó fehérjék működését érintik. A daganatkialakulás szempontjából érintett gének
A rákos elváltozások kialakulásához általában öt-hat egymást követő mutáció szükséges.
A felnőtt szervezetben osztódásra általában csak a nem véglegesen elkötelezett őssejtek képesek, bár kivételképpen megemlíthetők a máj sejtjei, melyek szükség esetén mégis visszatudják nyerni osztódóképességüket (regeneráció). Korábban úgy gondolták, hogy a daganatok csak a véglegesen elkötelezett, differenciált sejtekből fejlődnek az elköteleződés "visszaprogramozódásával", azonban ma már tudott, hogy számos daganat kiindulópontjául a sejtciklust csak átmenetileg elhagyó, de abba bármikor könnyen visszabillenthető őssejtek szolgálnak.
Bár a daganatok kialakulása minden esetben az örökítőanyag meghibásodására vezethető vissza, a daganatos betegségeket a kiváltó okok szerint alapvetően két csoportra oszthatjuk,
A rák nem egyetlen betegség, hanem egy gyűjtőfogalom, amely száznál is többféle megbetegedést foglal magába. Megkülönböztetünk jó- és rosszindulatú daganatokat.
Az elsődleges daganatok, jó- és rosszindulatú daganatok
A daganat lehet jóindulatú (benignus). Bár ezek is kontrollálhatatlanul növekednek, de
A rosszindulatú (malignus) vagy rákos daganatoknál
A rosszindulatú tumorokat két alapvető csoportba sorolják:
Az egyes ráktípusokat a megtámadott szerv alapján is elnevezték (például emlő-, tüdő-, gyomorrák stb.).
A rák hatása a szervezetre
A rák számos módon fenyegeti a szervezetet. Maga a daganat fizikailag hatást gyakorolhat a környezetében lévő szervekre, vezetékekre vagy véredényekre, fájdalmat és egyéb tüneteket okozva. Például az agyban lévő tumor fontos agyterületeket nyomhat (agyműködési zavarokat okozva), vagy a hasnyálmirigyben növekvő daganat elzárhatja az epevezetéket.
Amikor a rák betör a környező szövetekbe, a megsérült véredények miatt vérzések keletkezhetnek, a megtámadott szervek működésében pedig problémák lépnek fel.
A tumorsejtek bejuthatnak a vér- és a nyirokkeringésbe, és távoli szervekhez is eljuthatnak. E szervekben megtapadhatnak, s osztódásukkal megindulhat a távoli áttétek (metasztázisok) kialakulása.
Sajnos számos daganat esetében az elsődleges tumor sokáig anélkül növekedhet, hogy a beteg által észrevehető tüneteket okozna. Pedig a gyógyulásra sokkal nagyobb esély van, ha a kezelések idejében, az áttétek kialakulása előtt megkezdődnek. Ezért elengedhetetlenek a rendszeres szűrővizsgálatok.
67. tétel: Mendeli öröklődés |
Alapfogalmak
A gén
A gén az öröklődés anyagi egysége.
A DNS működése alapján lehet:
A fentieken kívül a DNS nagyrészt ún. nem-kódoló szekvenciákat – „hulladék DNS-t” – tartalmaz. Ezek ismétlődő, ismeretlen funkciójú vagy nem kódoló szakaszok. Az ember esetén ez a genom kb. 90%-a.
Egyéb alapfogalmak
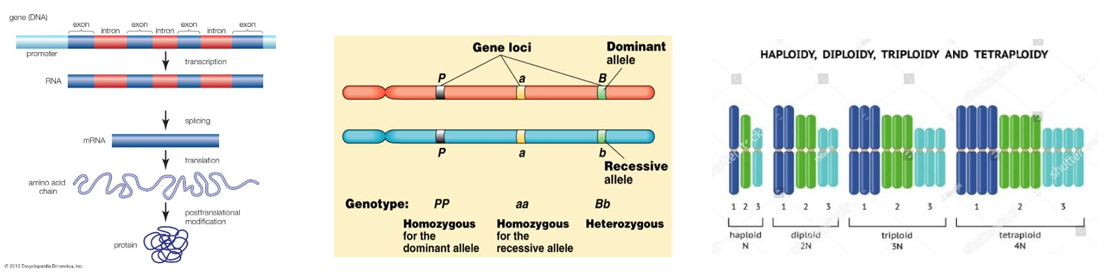 |
A diploid élőlények a nem osztódó testi sejtjeikben egy-egy kromatidából álló homológ kromoszómapárokat hordoznak – egy apai, ill. egy anyai változattal -, s így egy gén két kópiájával – alléljával - rendelkeznek.
Az allél lehet domináns (A) vagy recesszív (a).
 |
Mendeli analízis, klasszikus genetika, allélikus kölcsönhatások
Gregor Mendel (1822-1884)
Kerti borsóval dolgozott.
Többek között az alábbi tulajdonságokat vizsgálta:
Az élőlények alapvető molekuláris életfolyamatai, biokémiája, örökléstana igen hasonló, működéseik általános törvényszerűségeinek megismerését ún. modellszervezetek tanulmányozása teszi lehetővé. A modellszervezet tehát egy olyan élőlény, amelyet alapvető biológiai jelenségek megértése érdekében tanulmányoznak. A modellszervezetben tett felfedezések betekintést nyújtanak más élőlények működésébe. A tudománytörténet első genetikai modellszervezete a Mendel által tanulmányozott veteményborsó volt.
Előnyös, ha a modellszervezet
Genetikai modellszervezetek lehetnek:
 |
A mendeli öröklésmenet megállapításának módszere
(A tiszta vonalú egyedek egymás közötti keresztezésekor – pl. lilát lilával - az utódok ugyanazt a szülői fenotípust mutatják, elméletileg végtelen számú generáción keresztül, hiszen homozigóták.) A tiszta vonalú egyedeket Mendel több generáción keresztül önmegtermékenyítéssel hozta létre.
A mendeli szabályok hátterében az alábbi jelenségek állnak.
Mendel magyarázata
Mendel szabályai, törvényei
A Mendel-törvények általános érvényűek az élővilágban minden olyan élőlényre nézve, amelynek meiózisa van.
Egy haploid ivarsejt az adott tulajdonságpárból csak az egyik tulajdonságot hordozza.
Két eltérő – pl. lila-fehér -, tiszta vonalú – homozigóta - szülői vonal közötti keresztezés első utódnemzedéke egyöntetűen egyforma, uniform (heterozigóta). F1-ben nem fejeződik ki a fehér tulajdonság, mert a bíbor szín domináns, a fehér recesszív.
 |
Egy domináns fenotípusú egyed genotípusát ún. tesztelő keresztezéssel lehet megállapítani, melynek során egy recesszív fenotípusú – homozigóta - egyeddel kell keresztezni.
Amennyiben a vizsgált egyed
Az első utódnemzedék tulajdonságai függetlenek attól, hogy a tulajdonságot az apai vagy az anyai vonal hordozta. Mendel elvégezte a reciprok – fordított - keresztezéseket is, és ugyanolyan eredményeket kapott.
Az F1 egyedeit tovább keresztezve a második utódnemzedékben mindkét szülő tulajdonságai megjelennek 3:1 arányban.
Eddig egy allélpár által meghatározott tulajdonság öröklődését vizsgáltuk, az ilyen öröklésmeneteket egygénes vagy monohibrides öröklődésnek nevezzük.
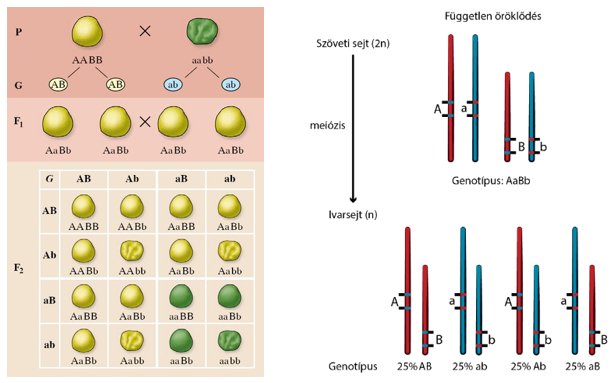 |
Két tulajdonságpárban – dihibrid - különböző növények öröklésmenete
Amely kimondja, hogy egyes tulajdonságok egymástól függetlenül öröklődnek. Ez azonban csak akkor igaz, ha a vizsgált tulajdonságokat meghatározó gének nem ugyanazon a kromoszómán, egymás közelében vannak. Ellenkező esetben ugyanis kapcsolt öröklődésről beszélünk, mert ilyenkor a kapcsolt tulajdonságok jellemzően együtt öröklődnek tovább, ebben az esetben nem érvényes ez a szabály.
A két tulajdonságpár:
P: AABB x aabb
sárga-sima zöld-ráncos
G: AB ab
F1: AaBb
sárga-sima
Eltérés a mendeli számarányoktól
A mendeli számarányok fenotípus szintjén való megnyilvánulása szigorú feltételekhez kötött. Mendel a róla elnevezett statisztikus számarányok (3:1, 9:3:3:1) és szabályok felfedezését a következő tényezőknek köszönhette:
Mendel a kísérleteiben valójában egyetlen fenotípus követésével egyetlen kromoszóma átörökítését „modellezte”. A mendeli törvények ezért a kromoszómák meiózisbeli viselkedését írják le, ami általánosan érvényes bármely gén öröklődési mintázatára.
Számos öröklésmenetet ismerünk, ahol az előbb felsorolt feltételek nem érvényesek, ekkor gyakran – de nem mindig - fenotípusszinten eltérést tapasztalunk a statisztikus mendeli 3:1 és 9:3:3:1 (tesztelő keresztezés esetén 1:1, ill. 1:1:1:1) számarányoktól.
A mendeli genetika kiterjesztése
Eltérések a mendeli szabályoktól
Az utolsó három esetben – V-VII – nem érvényesülnek maradéktalanul a mendeli szabályok.
 |
A dominanciaviszonyok lehetséges változatai
Nem teljes dominanciáról, akkor beszélünk, ha az egyik allél nem nyomja el teljesen a másik allél hatását, azaz a heterozigóta nem azonos az egyik homozigótával sem.
A nem teljes dominancia esete, az intermedier öröklésmenet
A tiszta vonalú – homozigóta - piros és fehér virágú csodatölcsér növények keresztezéséből származó F1 nemzedék virágai rózsaszínűek. Egyik szülő fenotípusa sem domináns. Ha az F1 nemzedék fenotípusa nem azonos egyik szülő fenotípusával sem, azok között átmeneti (köztes) jellegeket mutat, a dominancia nem teljes (intermedier jelleg).
Kodominancia
Akkor beszélünk kodominanciáról, ha a heterozigótában mindkét domináns allél egyformán megnyilvánul. Kodominancia esetén is a fenotípusok pontosan tükrözik a genotípusokat.
Ilyen például
A dominanciaviszonyok magyarázata
Egy gén két alléljának dominanciaviszonyaira a heterozigóták fenotípusából következtethetünk.
Többszörös allélek
Egy diploid szervezet egy génből csak két allélt hordozhat, hiszen a nem osztódó testi sejtekben két egykromatidás homológ kromoszóma van, ugyanakkor a populációban egy gén különböző alléljeinek száma nagyon nagy lehet, ezt nevezzük többszörös allélizmusnak. Egy gén alléljainak összességei allélsort képeznek. Az allélok lehetnek egyenértékűek, de mutathatnak változatos dominanciaviszonyokat is.
Az ember AB0 vércsoportjának négy vércsoportját egyetlen gén háromtagú allélsora határozza meg.
Letális allélek
Olyan allélok, melyek homozigóta formában a hordozójuk halálát okozzák.
Pleiotrópia
Azt a jelenséget, amikor egy gén egyszerre több tulajdonságot is befolyásol, pleiotrópiának nevezzük. A gének túlnyomó része pleiotróp. Erre példa a sarlósejtes vérszegénység, a fenilketonúria, cisztás fibrózis vagy az egerekben a bundaszínt meghatározó gén hatása, mivel a letalitást, a bundaszín kialakulását, továbbá elhízást, inzulinrezisztenciát, rákbetegségre való hajlamot egyaránt befolyásolja.
Allélizmus megállapítása
Egy fenotípusos jelleg meghatározását okozhatják:
Az allélizmus úgy állapítható meg, hogy a különböző jellegekre tiszta vonalakat (homozigóta) keresztezünk. Ha követik a mendeli egyfaktoros öröklésmenetet, (F2 = 3:1) akkor allélikus kölcsönhatásról van szó.
Több gén befolyása ugyanarra a jellegre (génkölcsönhatások)
A gének sokszor nem magukban, hanem más génekkel együttműködve, kölcsönhatásban alakítják ki a fenotípust.
Ha egy keresztezést olyan génekkel végzünk, amelyek mindegyike ugyanarra a fenotípusra hat, az megváltozott mendeli számarányokat eredményez. Ha a gének függetlenül öröklődnek, a számarány utalhat a génkölcsönhatás jellegére (lásd táblázat).
68. tétel: Kapcsoltság, nemhez kötött öröklődés |
A kapcsolt öröklődés
Mendel III. törvénye, a független kombinálódás törvénye, csak olyan tulajdonságokra vonatkozik, amelyek génjei különböző, nem homológ kromoszómán lokalizálódnak.
Ebben az esetben, amennyiben eltérő fenotípusú, homozigóta szülőkből indulunk ki, F2 nemzedékben a két tulajdonság szabadon kombinálódik, és a szülői fenotípusoktól eltérően új tulajdonságkombinációk jönnek létre (9:3:3:1).
Ennek oka, hogy F1 nemzedék egyedei 4-féle ivarsejtet hozhatnak létre azonos valószínűséggel (25%) a homológ kromoszómák véletlenszerű szétválása miatt.
Az emberi gének száma kb. 21 000, a genomban kromoszómák száma 23. Ebből következik, hogy kromoszómánként átlagosan 1000 génnek kell elhelyezkednie. Az egy kromoszómán elhelyezkedő gének úgymond kapcsoltan öröklődnek, ún. kapcsoltsági csoportot alkotnak, és mivel együtt öröklődnek (többé-kevésbé), nem vonatkozik rájuk a független kombinálódás törvénye.
Amennyiben a két gén egymás mellett van (teljes a kapcsoltság, nincs rekombináció) teljesen együtt mozognak, szemben a független öröklődéssel, a 4-féle ivarsejt helyett csupán 2-féle ivarsejt jön létre. Mivel a két gén együtt mozog, úgy viselkednek, mintha csupán egyetlen gén öröklődését vizsgálnánk.
Kapcsoltság esetén is megjelenhetnek a szülőktől eltérő rekombináns egyedek. A gyakoribb fenotípusok azonosak a szülők fenotípusaival, a szülőktől eltérő új kombinációk kisebb %-ban alakulnak ki. A jelenség a gének fizikai kapcsoltságával, továbbá a gének közötti rekombinálódással magyarázható, ami új rekombináns ivarsejtek kialakulását vonja maga után.
A rekombináció hátterében az átkereszteződés (crossing over) áll, amely folyamat a homológok között új allélkombinációt eredményez, aminek köszönhetően a heterozigóta egyedekben 4-féle ivarsejt jött létre.
 |
Rekombináció
A rekombináció során a meglévő tulajdonságokat kialakító örökítő tényezők újrarendeződnek, és ezzel új génkombinációkat hoznak létre. A rekombináció folyamata a meiózis során zajlik le, a meiózis eseményeihez köthető. Általában bármely folyamat, amely olyan új gén- vagy kromoszómakombinációkat eredményez, amelyek nem voltak jelen a kiindulási sejtben vagy annak elődeiben.
2-féle rekombinációt különböztetünk meg.
A folyamat során olyan haploid génkombináció jön létre a meiózis alatt, amely különbözik a haploid szülői génkombinációtól.
Ha a rekombinációs gyakoriság két gén esetén kisebb, mint 50%, akkor a két gén ugyanazon kromoszómán, kapcsoltan található.
Amennyiben a rekombinációs gyakoriság 50%, a két gén nem kapcsolt, és nagy valószínűséggel különböző kromoszómákon helyezkedik el.
A rekombinációs gyakoriságok ismeretében készíthetünk géntérképet. Rekombinációs gyakoriság: rekombináns utódok száma osztva az összes utód számával. A géntérképezés során a gének egymástól való távolságát, a genomban való viszonylagos elhelyezkedésüket határozzuk meg.
Minél távolabb van két gén a kromoszómán, annál nagyobb valószínűséggel játszódik le átkereszteződés közöttük a meiózisok során. Ehhez fel kell azonban tételezzük, hogy az átkereszteződés esélye a kromoszóma mentén mindenhol azonos.
A rekombináció eseményének valószínűsége nő a gének közötti távolsággal, ezért a rekombináció gyakoriságát mérve a gének közötti távolságra következtethetünk.
A géntérkép egysége (map unit, rövidítve m.u.) az a génpárok közötti távolság, amikor 100 utódsejtből egy rekombináns.
Másképp, 0,01 (vagy 1%) rekombinációs gyakoriság egyenlő 1 térképegységgel, melyet T.H. Morgan tiszteletére centimorgannak (cM) is neveznek.
Az ivarmeghatározás genetikája, nemhez kötött öröklődés
Az ivarmeghatározásnak az élővilágban igen sokféle formája ismert. A legtöbb esetben az ivarmeghatározásért különféle gének felelősek, melyek rendszerint speciális, ún. ivari kromoszómán helyezkednek el.
Az ivart meghatározó gének és az ivari kromoszómák
A legtöbb eukarióta élőlény sejtjeiben a kromoszómákat funkció szerint két csoportba oszthatjuk:
XX/XY rendszer
Ebben a rendszerben
Ez a rendszer jellemző az emlősökre és néhány rovarra (pl. kétszárnyúak, mint a muslica és a házi légy). Emberben a testi sejtekben 22 pár autoszóma mellet a 23. pár az ivari kromoszómapár.
A hím ivar meghatározásáért az Y-kromoszóma felelős, amely jóval kisebb, mint az X ivari kromoszómapárja, emberben hozzávetőleg 100 gént tartalmaz, amelyek főleg az ivarmeghatározásban játszanak szerepet. Legnagyobb része nem hordoz információt, így mindössze néhány rendellenesség – pl. a férfi meddőség egyes formái - társul specifikusan ehhez a kromoszómához.
Az X-kromoszómán ugyanakkor a női ivarmeghatározásáért felelős géneken kívül számos testi tulajdonságot kódoló gén található, ezek mutációi ún. X-kromoszómához kapcsolt rendellenességeket (vörös-zöld színtévesztés), ill. betegségeket (hemofília) eredményeznek.
Az X-, ill. az Y-kromoszómák eltérő méretük, alakjuk, ill. eltérő génállományuk miatt nem tekinthetők tipikus homológ kromoszómapárnak, bár vannak homológ szakaszaik néhány mindkét kromoszómán előforduló gén miatt. Ennek megfelelően a crossing over, ill. az intrakromoszomális rekombináció teljesen ki nem zárható, de kevéssé jellemző.
 |
Az ember ivarmeghatározása
Az utódokban 50%-os az egyes nemek kialakulásának a valószínűsége.
Az X-kromoszóma inaktivációja
A diploid szervezetekben általában egy bizonyos tulajdonságért felelős autoszomális gén mindkét allélja működhet. Ezzel szemben a nemi kromoszómák csak nőkben alkotnak homológ párokat (XX), a férfiakban az Y-kromoszóma nem funkcionális homológja az X-kromoszómának.
Míg az Y-kromoszóma kevés, főként a hím nem meghatározásáért felelős gént (pl. SRY) tartalmaz, addig az X-kromoszóma nagyszámú testi tulajdonságot meghatározó gént is hordoz. Ezért szükségessé válik a két nem eltérő X-kromoszomális géndózisait kiegyenlíteni. Ez a dóziskompenzáció, ami a női szervezetben az X-kromoszóma inaktivációja révén valósul meg.
Minden egyes sejtben a fejlődés során az egyik X-kromoszóma inaktiválódik, és ezt az állapotát aztán minden utódsejtben továbbörökíti. Ez az inaktiválódás véletlenszerű, vagyis akár az anyai, akár az apai eredetű X inaktiválódhat. A random X-inaktivációnak köszönhetően egy női szervezetben lesznek olyan sejtek, amelyekben az anyai, és lesznek olyanok, amelyekben az apai eredetű X-kromoszóma lesz inaktív. Vagyis emiatt a női szervezet egyfajta mozaikosságot mutat.
Az inaktiválódott X-kromoszóma egy kicsi, tömör kromatinszemcsét alkot, amely kitapad a sejtmaghártyájához és mikroszkóppal jól azonosítható. Ez a kromatinrög a Barr-test. Régebben szájnyálkahártya kaparékból vett mintából a Barr-test kimutatását a sportolók kromoszomális nemének meghatározására használták, mivel a kromatinrög csak nők testi sejtjeiben található meg (petesejtben nem, mert az haploid).
Ezzel a jelenséggel magyarázzuk a macskák vörös-fekete tarka szőrszín kialakulását. Ebben az esetben a szőrzetszín kialakításáért egy X-kromoszómához kötött gén kétféle alléja felelős. Az egyik allél vörös (Xv), míg a másik a fekete szín (Xf) meghatározását végzi. A vörös-fekete tarka macskák heterozigóták. Azokon a bőrfelületeken, ahol az Xv-kromoszóma inaktiválódik, a szőrszín fekete lesz, máshol, ahol az Xf inaktiválódik, a bőrterületek vörös színűek lesznek. Az előbbiekből következik, hogy a vörös-fekete tarka macska csak nőstény lehet.
Az X- és Y-kromoszómák mikroszkópban jól megkülönböztethetők: az Y-kromoszóma jóval kisebb az X-nél. Mindkét kromoszóma tartalmaz homológ és nem-homológ részeket.
Bár kevéssé jellemző, de az X- és Y-kromoszóma a homológ szakaszaik mentén képesek párosodni a meiózis I. főszakaszában, és ennek köszönhetően az adott gének esetleg rekombinálódhatnak.
A nem-homológ szakaszok olyan géneket hordoznak, amelyeknek nincs megfelelője a másik nemi kromoszómán, ezért férfiak esetén az ivari kromoszómákon található gének többségére – melyek pl. csak az X-kromoszómán találhatók meg - a genotípus feltüntetéséhez nem használhatjuk a homo-, ill. a heterozigóta fogalmakat, ebben az esetben az ún. hemizigóta kifejezést használjuk.
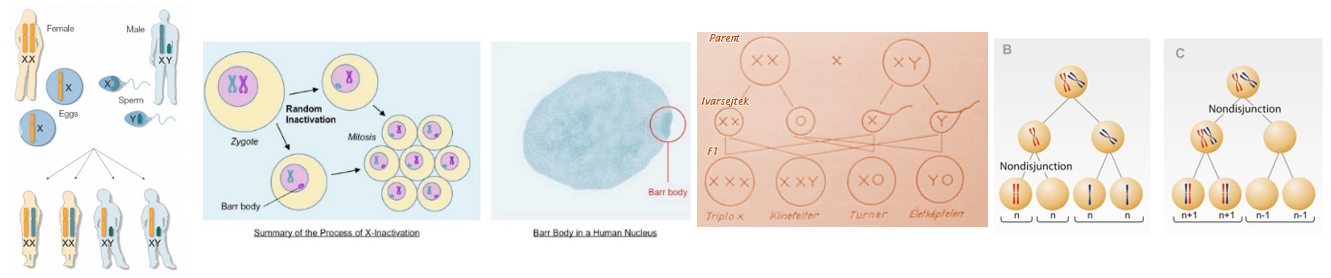 |
Nemhez kötött kromoszómamutáció
Ha az ivari kromoszómapár az ivarsejtek képződésekor zajló meiózis alatt nem válik szét – nondiszjunkció -, akkor lehetséges, hogy
A hibás esemény megtörténhet a meiózis első, ill. a második főszakaszában egyaránt. A rendellenesség mindkét szülőben előfordulhat.
Nemhez kötött öröklődés
Az X ivari kromoszómán az ivart meghatározó géneken kívül számos tulajdonság génje lokalizálható. Ezek a tulajdonságok nemhez – X-kromoszómához - kapcsoltan öröklődnek.
A megtermékenyítést követően
Az előbbieknek megfelelően az apa a lányának, az anya a fiának örökíti az X-kromoszómán lévő tulajdonságait. Ez a criss-cross szabály. Ezért recesszív jellegű betegségek esetén
Az X-kromoszómához kötött recesszív öröklésmenet jellemzői (XR)
X-kromoszómához kötődő recesszív rendellenességek
X-kromoszómához kötődő domináns rendellenességek (XD)
Y-kromoszómához kötött rendellenességek
A fül túlzott szőrössége az egyetlen férfiról férfira átadódó testi jelleg, a szőrös fül nem Y-hoz kötött, de génje és annak helye még nem ismert (Tóth Sára Genetika és Genomika).
Az Y-kromoszóma legnagyobb része nem hordoz információt, így mindössze néhány rendellenesség – pl. a hímivarsejtképzés zavaraival összefüggő férfi meddőség egyes formái - társulnak specifikusan ehhez a kromoszómához.
Az Y-kromoszómához kötött öröklődés jellemzői
69. tétel: Vércsoportok és öröklődésük |
Az emberi vércsoportok
A vörösvértestek felszínét borító sejtköpenyben megtalálható glikoproteidek – vércsoportantigének - alapján az emberi populációban többféle vörösvértest típus fordul elő. Ennek alapján több mint 40 egymástól független vércsoportrendszert különböztetünk meg, azonban a vérátömlesztés szempontjából, ebből csupán kettőnek van gyakorlati jelentősége, ez az AB0-rendszer és az Rh-rendszer, melyek az emberszabású majmokban is előfordulnak (ezenkívül MN, Langereis, Junior vércsoport stb.). Az emberi vércsoportokat a 20. század elején Landsteiner fedezte fel, amiért később Nobel-díjat kapott. A vörösvértestek sejthártyájának felszínén, a vércsoportok kialakításáért felelős glikoproteidek – vércsoportantigének - többnyire transzportfehérjék.
Az AB0 vércsoportrendszer
Ebben az esetben kétféle vércsoportot meghatározó molekulát – antigént - különböztetünk meg, az egyik típus az A, a másik típus a B vércsoport kialakításáért felelős. A vércsoportantigének elhelyezkedése és minősége alapján négy nagyobb vércsoportot különböztetünk meg, A, B, AB, 0. Az egyes típusokon belül még további altípusok ismertek pl. A1, A2, A3, A4, A5.
Egy adott vércsoport vérplazmájában - az AB kivételével - a másik vércsoport vörösvértestei ellen termelődő ellenanyagok, antitestek vannak.
Az A és B antigének akkor jelenhetnek meg a vértestek felszínén, ha egy speciális szénhidrát-molekulához, a H-antigénhez kapcsolódnak. A H antigén a 0-s vércsoportú emberek vérében is megtalálható, de ezekben a személyekben nem kapcsolódik hozzá sem A, sem B antigén.
Létezik egy olyan mutáció, amelynek következtében nem működik az az enzim, ami létrehozná ezt a H-antigént. Ezt a jelenséget Bombay-jelenségnek nevezik. Ebben az esetben az érintett személyek 0-ás vércsoportúak, mivel nem alakulhat ki sem A sem B vércsoport, tekintve, hogy az A ill. a B antigének csak akkor jelenthetnek meg a vörösvértestek felületén, ha jelen van a H antigén, amihez kötődhetnek.
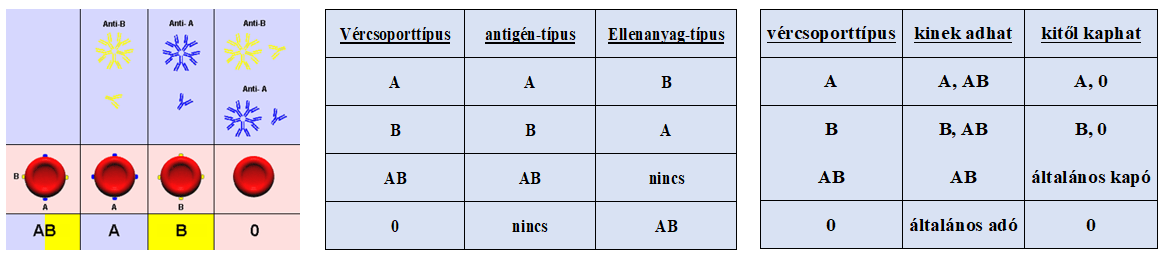 |
Vérátömlesztésnél elsődlegesen csoportazonos vért kell használni, ha ez nem áll rendelkezésre, különösen kell vigyázni és betartani az alábbi szabályokat, mivel pl., ha egy B vércsoportú ember A típusú vért kap, a vérplazmában lévő A ellenanyagok kicsapják a beadott A vörösvértesteket, ami a képződött vérrögök miatt súlyos, esetleg életveszélyes következményekkel járhat.
A beadott vérkészítmények vérplazmamentesek, mivel azt előzőleg eltávolították és a vörösvértesteket fiziológiás sóoldatba vitték, így csak arra kell ügyelni, hogy a beadott sejtek ne csapódjanak ki.
A Bombay-jelenségben érintett személyek nem kaphatnak vért még 0 vércsoportú donortól sem, mert a normális, nem Bombay-mutációs 0-s vércsoportban jelen van a H-antigén, ami idegen antigén a Bombay-mutációban szenvedők számára, ezért a vérplazmában megjelenő anti-H antitestek kicsapnák a beadott 0-s vörösvértesteket.
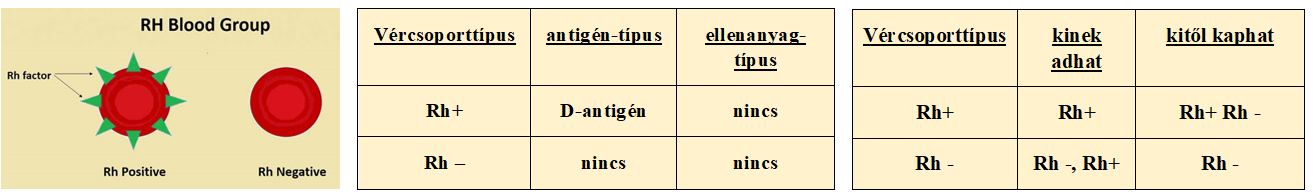 |
Rh vércsoportrendszer
A vércsoportrendszer nevét onnan kapta, hogy a Macacus Rhesus majomban fedezték fel.
Itt kétféle vércsoport van aszerint, hogy az Rh-faktor (D-antigén) megtalálható-e vagy sem a vörösvértestek felületén. Az első esetben Rh+-, a második esetben Rh-csoportról beszélünk.
Rh ellenanyag eredendően nincs a vérplazmában, csak abban az esetben termelődik, ha egy Rh- ember Rh+ vért kap, hiszen az Rh+ vörösvértestek - az Rh-faktor miatt - testidegen anyagként immunreakciót indítanak be.
Régebben súlyos gondokat okozott az Rh-összeférhetetlenség problémája. Amikor egy Rh- nő Rh+ gyereket szül, szülés közben a kétféle vér érintkezésekor az anya vérében megindul az ellenanyag képződés. Ez az első terhesség alatt azért nem okoz problémát, mert az anyai és a magzati vér nem keveredik egymással.
Azonban, ha a második gyerek is Rh+, az anya vérében már jelenlevő ellenanyag átjutva a méhlepényen kicsapja a magzat vörösvértesteit, annak életét súlyosan veszélyeztetve.
A szülést követően 72 órán belül az anyának Rh+ ellenanyagot adva megakadályozható az immunreakció kialakulása.
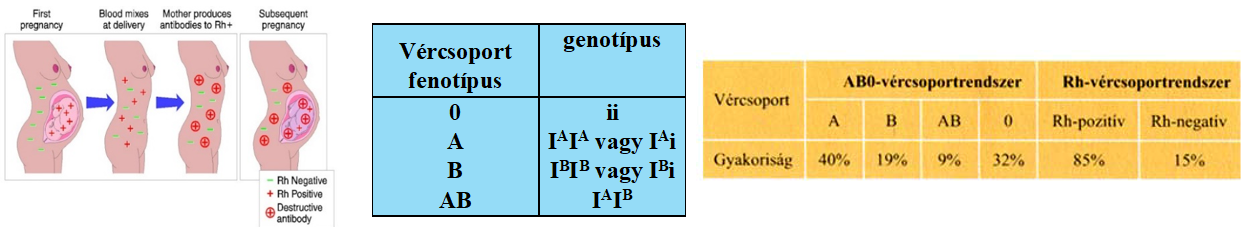 |
A vércsoportok öröklődése
Az ember AB0 vércsoportjának négy vércsoportját egyetlen gén háromtagú allélsora határozza meg.
Az RH vércsoport kialakításáért egyetlen autoszomális gén kétféle allélja felelős.
Az előbbiekből következően
70. tétel: Darwin evolúciós elmélete |
Az evolúció fogalma
A biológiában evolúciónak, törzsfejlődésnek nevezzük az élővilág többnyire fokozatos és szüntelen történeti fejlődését. Ugyanakkor az evolúció fogalmát más szinteken is értelmezzük. Tágabb értelemben az evolúció egy lassú, fokozatos, bizonyos irányba haladó változást jelent. Az evolúció jelensége így akár az Univerzum fejlődésében, a társadalmi változásokban vagy a tudományos elméletek alakulásában is megjelenik.
A törzsfejlődés során a fajok öröklődő jellegei nemzedékről nemzedékre változnak, ami
Az evolúció során a fajok a változó környezeti feltételekhez folyamatosan alkalmazkodnak, melynek során génállományuk, ezen keresztül tulajdonságaik szüntelenül változnak.
Az evolúció eszmerendszere ma teljes mértékben áthatja a természettudományos gondolkodást, megkérdőjelezhetetlen rendezőelvként van jelen.
Darwinizmus
Az evolúcióelmélet kidolgozója és névadója Charles Darwin (1809 –1882) angol természettudós.
Nem Darwin fogalmazta meg az evolúció elméletét, jelentősége nem ebben áll, hanem ő volt az, aki először megalkotott egy tudományosan kidolgozott mechanizmust az evolúció magyarázatára.
A fajok átalakulását a tudósok jelentős hányada elfogadta Darwin előtt is, azonban meggyőző magyarázatot Darwin adott A fajok eredete (1859) című művében a fajok kialakulására, az élőlények sokféleségére. A mű fogadtatása a várakozásoknak megfelelően elutasító volt, de később a tudományosan megfogalmazott elmélet lassan elfogadottá vált. (A mű teljes címe: A fajok eredete természetes kiválasztás útján, vagyis az előnyös változatok fennmaradása a létért folyó küzdelemben.)
A 22 éves Darwin természettudományos gondolkodására óriási hatást gyakorolt Beagle nevű hajóján tett 5 évig tartó világkörüli tudományos expedíciója, melynek során jutott el a Galápagos-szigetcsoportra is. Mire hazaért, már valóban biztos volt abban, hogy az evolúció létezik, de még nem találta meg működésének mechanizmusát.
Később, Lamarck gondolatait a természetes kiválasztódás, a természetes szelekció elméletével egészítette ki. Darwin elmélete közvetlen megfigyelésen alapult. Úgy vélte, hogy a természetben több egyed születik, mint amennyi később életben maradhat, mert a környezet erőforrásai végesek. Mivel a fajon belül az egyedek nem egyformák, Darwin szerint a természetes szelekció (kiválogatódás) során a létért való küzdelemben az alkalmas változatok – a rátermettebbek - fennmaradnak, tulajdonságaikat tovább örökíthetik, míg a kevésbé alkalmasak elpusztulnak, kiszelektálódnak.
A folyamatban az élőlények öröklődő tulajdonságai közül az előnyös jellegek gyakorisága nő, az előnytelenek gyakorisága pedig csökken az egyes nemzedékek során. Ha az előnyös ismertetőjegyek az öröklődés során átadódnak a következő nemzedéknek, akkor idővel túlsúlyra juthatnak a populáción belül, és a változások új faj létrejöttét eredményezhetik.
Az élőlények ezáltal alkalmazkodnak – adaptálódnak - az adott környezethez. A környezeti tényezők megváltozása azonban új alkalmazkodási folyamatot igényel, melynek során előtérbe kerülhetnek korábban semleges vagy akár hátrányos tulajdonságok is a populációban.
 |
Tehát az evolúció olyan folyamatos változások sorozata, melynek során a fajok egyes öröklődő jellegeinek gyakorisága nemzedékről nemzedékre változik.
Az evolúció jellemzői tehát
A Manchester környéki nyírfaaraszoló lepkéknek világos és sötét mintázatú változatai ismertek. A 18. század első felében e lepkének szinte csak a világos példányai fordultak elő, mivel a nyírfák világos kérgén a sötét mintázatú egyedeket a madarak könnyen észrevették és elfogyasztották (kiszelektálták). A sötét színű egyedek gyakoriságát a szelekció folyamatosan alacsonyan tartotta. Az ipari forradalom idején sötét koromszennyezés borította be a fák kérgét, aminek köszönhetően a világos egyedek váltak feltűnővé, ezért néhány évtized alatt a környék nyírfaaraszoló -populációiban a sötét és világos példányok aránya megfordult.
A fentiek szerint evolúciós változást eredményezhet az is, ha egy populáción belül folyamatosan változik az egyes tulajdonsággal jellemezhető egyedek aránya, gyakorisága, azaz a törzsfejlődéshez nem feltétlenül szükséges új tulajdonságok megjelenése.
Szelekció (kiválogatódás)
A szelekció során az élőlények nemzedékeinek váltakozásával az öröklődő tulajdonságok közül az előnyös jellegek gyakorisága nő, az előnytelenek gyakorisága pedig csökken. A szelekció előfeltétele a változatosság.
Formái a természetes és a mesterséges szelekció.
A természetes szelekció a létért való küzdelemben a legrátermettebb genotípusok elterjedését és a kevésbé rátermett egyedek kiküszöbölődését jelenti. A szelekcióval a populáció generációk során át - alkalmazkodik – adaptálódik – az adott környezethez. A folyamatot adaptív evolúciónak tekintjük.
A populáció egyedei – egyes genotípusai – különbözők, eltérőek a túlélési, szaporodási esélyeik. Az egyes genotípusok túlélése, szaporodási képessége, az utódnemzedékekben való megjelenése genetikai rátermettségétől függ. A rátermettség (fitness) egy adott genotípus megjelenésének a valószínűsége az utódnemzedékben. Ez a tényező képezi a természetes szelekció alapját.
A szelekció iránya, a mennyiségi jellegek szelekció típusai
1. Irányító szelekció
A mesterséges szelekció az ember tervszerű tevékenysége, amely révén a számára megfelelő tulajdonságokat nemzedékről nemzedékre gyarapítja a kiválasztott élőlények csoportjaiban.
2. Szétválasztó szelekció
3. Stabilizáló szelekció
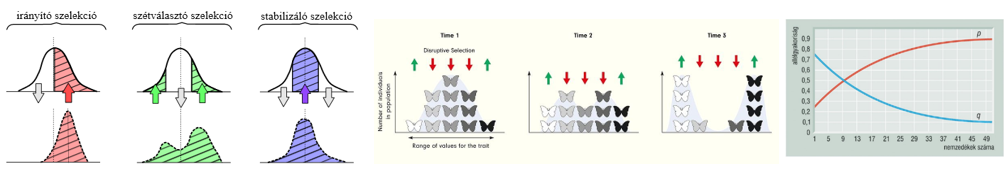 |
Teljes és részleges szelekció, mutáció-szelekció
Ha valamely genotípus fitness értéke 0, akkor teljes szelekcióról beszélünk, ekkor az egyedek az ivarérettség elérése előtt elpusztulnak.
Ugyanakkor ismertek olyan speciális esetek, amikor a recesszív allél homozigóta formában letális, azonban heterozigótákban szelekciós előnyt biztosít a hordozójának. A jeleséget heterozigóta fölénynek nevezzük. Erre példa a sarlósejtes vérszegénységre nézve heterozigóták gyakori előfordulása Afrikának azon területein, ahol a malária pusztít.
Sarlósejtes anémia hátterében egy, a hemoglobin β-lánc génjében bekövetkezett pontmutáció áll, aminek következtében a hemoglobin molekulák összecsapódnak, kikristályosodnak, oxigénkötő képességük jelentősen csökken és vörösvértestek sarló alakúvá válnak. Mivel a károsodott sejtek szétesnek, vérszegénység, vérkeringési zavar és csökkent oxigén-ellátottság jelentkezik.
A tünetek homozigóta egyéneknél súlyosak, hiszen bennük kizárólag hibás hemoglobin található. A heterozigóták esetében a hemoglobin molekulák egy része normális, így az összecsapódási hajlam is kisebb, így ők tünetmentesek. Náluk csak oxigénhiány miatt jelentkezhetnek a tünetek, pl. magas hegyi tartózkodás, vagy erősebb fizikai terhelés esetén.
Részleges szelekció esetében az utódok életképesek, de valamiért különböző mértékű szaporodási hátrányban vannak.
A szelekció – evolúció – szintjei, többszintű evolúció
A biológiai szerveződés többszintűsége egyúttal magával hozza a szelekció, illetve az evolúciós mechanizmusok többszintűségét.
1. A génközpontú evolúciófelfogás szerint a szelekció alapegységei a tulajdonságokat meghatározó gének. A szelekció a gének, ill. az allélok között zajlik, és az élettartamuk, termékenységük és másolási megbízhatóságuk szerint válogat köztük. Az evolúcióban azok a gének, allélok voltak sikeresek, azok maradhattak fent, amelyek sikeresen tudták átörökíteni magukat a múltban.
2. Az egyed szintű szelekció a populáción belül a legfontosabb, eddig alapvetően ezzel foglalkoztunk, az eltérő fitnesz hátterében genotípus, illetve öröklődő fenotípus különbségek állnak.
3. Populáció szintű szelekció során a populációk alkalmazkodnak a környezetükhöz. Egy nagyobb genetikai változatossággal rendelkező populáció – ahol egy-egy génnek igen sok alléja fordulhat elő - sokkal nagyobb eséllyel képes alkalmazkodni az állandóan változó környezeti viszonyokhoz. Ki kell emelni, hogy a populáció alkalmazkodóképességének a javulása nem az egyedek valamiféle genetikai megváltozása életük során, hanem a populáció genetikai összetételének megváltozása révén következik be, aminek mozgató rugója az egyedek közötti természetes szelekció. A kiválogatódás során a populáció gyengébb fitneszű egyedei kisebb-nagyobb számban elpusztulnak, amely a populáció genetikai összetételének, általában allélgyakoriságának változását eredményezi (mikroevolúció).
 |
A populáció szintű szelekció kapcsán említést kell tennünk a koevolúció jelenségéről. A koevolúció a biológiában két faj egymással kölcsönhatásban történő evolúcióját jelenti. A koevolúció során egy populáció génállománya a természetes szelekciónak köszönhetően egy másik populáció hatására megváltozik. A koevolúcióra számos példa ismert:
Az alkalmazkodóképesség kulcsa a diverzitásban, a szervezetek, fajok, gének sokféleségében keresendő.
A szelekció tehát több szinten zajlik egyszerre: gének, egyedek, populáción belüli csoportok, populációk, fajok szintjén egyszerre.
Fajok keletkezése
A biológiai evolúció során a populációk, ill. a fajok öröklődő tulajdonságai maradandóan megváltoznak és ennek eredményeképpen új fajok jöhetnek létre.
A fajképződés folyamata során egy faj különböző populációi olyan mértékben megváltoznak, hogy az őket alkotó egyedek már nem tudnak egymással szaporodni.
A genetikai összetételt megváltoztató mechanizmusok lehetnek adaptív jellegűek, a populációk a szelekciós folyamatok következtében lassan, fokozatosan alkalmazkodnak a környezetükhöz, melynek során az arra alkalmas genotípusok kiválogatódnak.
Fokozatos evolúció
Többnyire lassú, sok generáción keresztül valósul meg.
Izoláció
A fajok térben többé-kevésbé elhatárolódó populációkból állnak. Az izoláció során egy faj különböző populációi, ill. egy populáció egyes részei elkülönülnek egymástól, aminek eredményeképpen az elkülönülő részek között lehetetlenné válik az egyedek vándorlása. Az elkülönülő populációk a genetikai sodródás, ill. amennyiben a szétváló csoportok környezete megváltozik, az adaptáció révén sajátos – eltérő - úton fejlődhetnek.
Ökológiai izoláció (Szimpatrikus fajképződés)
A fajok populációi ugyanazon földrajzi területen különböző élőhelyeket foglalnak el, és így általában nem találkoznak egymással. Az eltérő ökológiai környezethez való alkalmazkodás eredményeképpen az elkülönülő populációrészek egyedei sajátos úton fejlődhetnek.
 |
Az ökológiai fajképződés egy jellemző példája az adaptív szétterjedés (radiáció)
Előfordul, hogy a populációk egyedei a túlnépesedés miatt kiszorulnak eredeti élőhelyükről. Az új élőhelyen a megváltozott körülményekhez a populációk alkalmazkodnak (adaptálódtak), melynek során az arra alkalmas genotípusok kiválogatódnak.
Ilyen a Galápagos-szigeteken élő pintyek példája.
Adaptív radiációval magyarázhatjuk pl. a szárazföldi gerinces állatok különböző végtagtípusait. Minden végtagtípus az ötujjú típusra vezethető vissza, azonban az egyes végtagok felépítése az evolúció során alkalmazkodott a különféle környezeti körülményekhez. Az ilyen hasonló eredetű, alapszabásában megegyező szerkezetű, azonban az eltérő környezethez való alkalmazkodás miatt különböző megjelenésű szerveket homológ szerveknek nevezzük. Az eltérő fejlődés a divergencia, melynek során tehát az élőlények egyes szerveinek a felépítése alkalmazkodik az eltérő környezeti feltételekhez.
A divergenciával ellentétes fejlődés a konvergencia, melynek során a törzsfejlődéstanilag eltérő eredetű és szerkezetű szervek a hasonló környezethez való alkalmazkodás miatt hasonlóvá válhatnak.
A konvergens fejlődés eredményeként létrejövő hasonló szerveket analóg szerveknek nevezzük. Ilyen pl.
71. tétel: Adaptív és nem adaptív evolúció |
A biológiában evolúciónak, törzsfejlődésnek nevezzük az élővilág többnyire fokozatos és szüntelen történeti fejlődését.
A törzsfejlődés során a fajok öröklődő jellegei nemzedékről nemzedékre változnak, ami
Az evolúció során a fajok a változó környezeti feltételekhez folyamatosan alkalmazkodnak, melynek során génállományuk, ezen keresztül tulajdonságaik szüntelenül változnak.
A törzsfejlődés hátterében tehát olyan folyamatos változások sorozata áll, melynek során a fajok öröklődő jellegei nemzedékről nemzedékre változnak. Az evolúció során egy faj különböző populációiban bizonyos tulajdonságok megritkulnak, mások gyakoribbá válnak, ill. új tulajdonságok jelenhetnek meg. Ezáltal a faj populációi az idők során kisebb-nagyobb mértékben különbözőkké válnak. A változások halmozódásával a populációk egyedei oly mértékben eltérővé válhatnak elődeiktől, hogy már nem is tekinthetők egy fajba tartozóknak. A klasszikus populációgenetika a génváltozatok (allélok) populációbeli arányának, ill. egyes allélok gyakoriságának megváltozását elemi evolúciós változásnak tekinti.
Változatosság, sokféleség, diverzitás
A biodiverzitás biológiai sokféleség. Az élet megjelenési formáinak gazdagságát, az élővilág változatosságát és változékonyságát kifejező fogalom. Összetettségénél fogva a biológiai szerveződés több szintjén értelmezhető.
Genetikai diverzitás
A genetikai diverzitásnak négy szintjét különböztetjük meg:
A genetikai diverzitás jelentősége abban rejlik, hogy egy nagyobb változatossággal rendelkező populáció sokkal nagyobb eséllyel képes alkalmazkodni az állandóan változó környezeti viszonyokhoz, hiszen ha megváltozik a környezet, a populáción belül mindig lesznek olyan egyedek, melyek az új környezetben rátermettebbek lesznek a többieknél. A rátermettebb egyedek nagyobb eséllyel szaporodnak, nagyobb eséllyel örökítik át genetikai állományukat, így a következő generációkban tulajdonságaik egyre elterjedtebbek lesznek, aminek eredményeképpen a populáció lassan megváltozik (lásd Manchester környéki nyírfaaraszoló lepkék). Amennyiben a változást okozó tényezők hosszabb ideig egy irányba mutatnak, új faj kialakulására van lehetőség.
 |
A populációk genetikai összetételét módosító hatások
A populációk genetika összetételének változása átalakítja a populációt genetikailag, amely tehát evolúciós átalakulást eredményezhet.
Nem adaptív evolúció
Véletlenszerű, előre nem látható, genetikai diverzitást növelő tényezők, nem adaptív evolúciós folyamatok
1. Mutáció
Az evolúció során az egyes allélváltozatok mutációval jöttek létre. Ma az élőlények az evolúció során felhalmozódott kedvező mutációkat hordozzák.
2. Rekombináció
A rekombináció során a tulajdonságokat kialakító genetikai tényezők újrarendeződnek, így új génkombinációk a szülőktől eltérő fenotípusokat eredményeznek.
3. Génáramlás-bevándorlás
Az egymás mellett élő populációk között az egyedek vándorolhatnak. A bevándorlás megváltoztatja a populáció genetikai összetételét, hiszen a jövevény egyedek új allélokat hozhatnak magukkal, így megváltoztatják az eredeti allélgyakoriságokat.
Például a B vércsoport elterjedése Európában a tatárjárásnak „köszönhető”.
Véletlenszerű, genetikai diverzitást csökkentő tényezők, nem adaptív evolúciós folyamatok
1. Genetikai sodródás (drift)
Általában kis létszámú, elszigetelt populációkra jellemző. Ha egy populációban az egyedek száma lecsökken, megváltoznak az allélgyakoriságok, elveszhetnek hasznos, fontos allélek, ezért többnyire csökken a populáció élet- és szaporodóképessége. Az elméletileg várt hasadási arányok nem tapasztalhatók, egyes genotípusok kiveszhetnek a populációból. Általában lerontja a populáció tulajdonságait, ugyanakkor a fajképződés egyik lehetséges útja (pl. endemikus fajok kialakulása).
A kihalási küszöbaz a kritikus populációnagyság, ami alatt a populáció már nem szaporodóképes.
A populációk optimális nagyságának óriási a természetvédelmi – fajmegőrzési - jelentősége, ugyanis minél kisebb az egyedszám, annál nagyobb a beltenyésztés, ill. a genetikai sodródás veszélye, melyek a populációk genetikai leromlását eredményezik.
A genetikai sodródás szélsőséges példája az alapítóelv. Ilyenkor egy nagy létszámú populáció kis egyedszámú része elkülönül, például egy szigetre való betelepülés során vagy valamilyen katasztrófa révén. Az alapító egyedek genetikai állománya eltérhet az eredeti populáció átlagától.
Az észak-amerikai indiánok többsége 0 vércsoportú, míg B vércsoport nem fordul elő közöttük, ami azért meglepő, mert Amerika benépesítése Ázsia felől történt meg, ahol pedig a B vércsoport a leggyakoribb vércsoport. Korábbi elképzelés szerint egy apró, alig száz fős csoport kivált az ázsiai őslakók közül és a befagyott Bering-szoroson keresztül átvándorolt Észak-Amerikába. (Újabban kétségbe vonják ezt, több hullámban történő benépesítést valószínűsítenek.)
2. Beltenyészet
A rokonok közötti szaporodást beltenyésztésnek nevezzük, melynek során a homozigóták aránya egyre nő. A rokon egyedek nagyobb mennyiségben hordoznak hasonló allélokat, emiatt a recesszív allélok találkozásának esélyei is megnövekednek. A recesszív allélok leggyakrabban hátrányos jellegeket – betegségeket - alakítanak ki homozigóta formában.
Szélsőséges esete az önmegtermékenyítés, ekkor minden generációban felére csökken a heterozigóták aránya.
3. Elvándorlás
 |
Adaptív evolúció
1. Nem véletlenszerű, genetikai diverzitást csökkentő tényezők, adaptív evolúció
Szelekció (kiválogatódás)
A szelekció során az élőlények nemzedékeinek váltakozásával az öröklődő tulajdonságok közül az előnyös jellegek gyakorisága nő, az előnytelenek gyakorisága pedig csökken. A szelekció előfeltétele a változatosság.
Formái a természetes és a mesterséges szelekció.
A természetes szelekció a létért való küzdelemben a legrátermettebb genotípusok elterjedését és a kevésbé rátermett egyedek kiküszöbölődését jelenti. A szelekcióval a populáció generációk során át - alkalmazkodik – adaptálódik – az adott környezethez. A folyamatot adaptív evolúciónak tekintjük.
A populáció egyedei – egyes genotípusai – különbözők, eltérőek a túlélési, szaporodási esélyeik. Az egyes genotípusok túlélése, szaporodási képessége, az utódnemzedékekben való megjelenése genetikai rátermettségétől függ. A rátermettség (fitness) egy adott genotípus megjelenésének a valószínűsége az utódnemzedékben. Ez a tényező képezi a természetes szelekció alapját.
A szelekció iránya, a mennyiségi jellegek szelekció típusai
1. Irányító szelekció
A mesterséges szelekció az ember tervszerű tevékenysége, amely révén a számára megfelelő tulajdonságokat nemzedékről nemzedékre gyarapítja a kiválasztott élőlények csoportjaiban.
2. Szétválasztó szelekció
3. Stabilizáló szelekció
Teljes és részleges szelekció, mutáció-szelekció
Ha valamely genotípus fitness értéke 0, akkor teljes szelekcióról beszélünk, ekkor az egyedek az ivarérettség elérése előtt elpusztulnak.
Ugyanakkor ismertek olyan speciális esetek, amikor a recesszív allél homozigóta formában letális, azonban heterozigótákban szelekciós előnyt biztosít a hordozójának. A jeleséget heterozigóta fölénynek nevezzük. Erre példa a sarlósejtes vérszegénységre nézve heterozigóták gyakori előfordulása Afrikának azon területein, ahol a malária pusztít.
Sarlósejtes anémia hátterében egy, a hemoglobin β-lánc génjében bekövetkezett pontmutáció áll, aminek következtében a hemoglobin molekulák összecsapódnak, kikristályosodnak, oxigénkötő képességük jelentősen csökken és vörösvértestek sarló alakúvá válnak. Mivel a károsodott sejtek szétesnek, vérszegénység, vérkeringési zavar és csökkent oxigén-ellátottság jelentkezik.
A tünetek homozigóta egyéneknél súlyosak, hiszen bennük kizárólag hibás hemoglobin található. A heterozigóták esetében a hemoglobin-molekulák egy része normális, így az összecsapódási hajlam is kisebb, így ők tünetmentesek. Náluk csak oxigénhiány miatt jelentkezhetnek a tünetek, pl. magashegyi tartózkodás vagy erősebb fizikai terhelés esetén.
Részleges szelekció esetében az utódok életképesek, de valamiért különböző mértékű szaporodási hátrányban vannak.
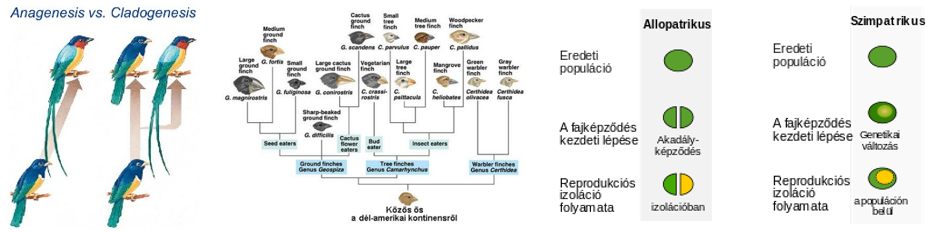 |
Fajok keletkezése
A biológiai evolúció során a populációk, ill. a fajok öröklődő tulajdonságai maradandóan megváltoznak és ennek eredményeképpen új fajok jöhetnek létre.
A fajképződés folyamata során egy faj különböző populációi olyan mértékben megváltoznak, hogy az őket alkotó egyedek már nem tudnak egymással szaporodni.
A genetikai összetételt megváltoztató mechanizmusok lehetnek
Filogenetikai szempontból a fajképződés történhet úgy, hogy
A kérdés az, hogy milyen események válthatják ki a fent említett genetikai összetételbeli változást eredményező mechanizmusokat?
1. Fokozatos evolúció
Többnyire lassú, sok generáción keresztül valósul meg.
Izoláció
A fajok térben többé-kevésbé elhatárolódó populációkból állnak. Az izoláció során egy faj különböző populációi, ill. egy populáció egyes részei elkülönülnek egymástól, aminek eredményeképpen az elkülönülő részek között lehetetlenné válik az egyedek vándorlása. Az elkülönülő populációk a genetikai sodródás, ill. amennyiben a szétváló csoportok környezete megváltozik, az adaptáció révén sajátos – eltérő - úton fejlődhetnek.
Izoláció oka lehet:
Földrajzi (Allopatrikus fajképződés)
Szaporodási
Ökológiai (Szimpatrikus fajképződés)
Az ökológiai fajképződés egy jellemző példája az adaptív szétterjedés (radiáció)
Előfordul, hogy a populációk egyedei a túlnépesedés miatt kiszorulnak eredeti élőhelyükről. Az új élőhelyen a megváltozott körülményekhez a populációk alkalmazkodnak (adaptálódtak), melynek során az arra alkalmas genotípusok kiválogatódnak.
Ilyen a Galápagos-szigeteken élő pintyek példája.
Adaptív radiációval magyarázhatjuk pl. a szárazföldi gerinces állatok különböző végtagtípusait. Minden végtagtípus az ötujjú típusra vezethető vissza, azonban az egyes végtagok felépítése az evolúció során alkalmazkodott a különféle környezeti körülményekhez. Az ilyen hasonló eredetű, alapszabásában megegyező szerkezetű, azonban az eltérő környezethez való alkalmazkodás miatt különböző megjelenésű szerveket homológ szerveknek nevezzük. Az eltérő fejlődés a divergencia, melynek során tehát az élőlények egyes szerveinek a felépítése alkalmazkodik az eltérő környezeti feltételekhez.
A divergenciával ellentétes fejlődés a konvergencia, melynek során a törzsfejlődéstanilag eltérő eredetű és szerkezetű szervek a hasonló környezethez való alkalmazkodás miatt hasonlóvá válhatnak.
A konvergens fejlődés eredményeként létrejövő hasonló szerveket analóg szerveknek nevezzük. Ilyen pl.
2. Az ugrásszerű evolúció
Rövid idő – egy-két nemzedék - alatt végbemenő evolúciós változás. Néhány egyedre terjed csak ki, amelyekből új faj keletkezhet. Módjai:
1. Génáramlással, genetikai sodródással
A genetikai sodródás kisméretű, elszigetelt populációkban akkor is megváltoztatja az allélgyakoriságokat, ha a környezet állandó, azonban, ha változik a környezet, a szelekció-adaptáció ráerősíthet a fajképződés folyamatára.
2. Génmutációval, kromoszómamutációval
Génmutációval új génváltozatok – allélok - alakulnak ki, melyek egy környezetváltozás során hasznossá válhatnak.
Kromoszómamutációval új gének, ill. allélok ugyan nem jelennek meg, de új fenotípusok jönnek létre.
A poliploidizáció (genomsokszorozódás) is hasonló eredményt hoz, ami úgy jön létre, hogy az osztódások során a kromoszómák valamilyen módon megsokszorozódnak, pl.
72. tétel: Populációgenetika |
Populációgenetika
A populációk genetikai összetételének leírása
A Hardy-Weinberg-modell
A populációgenetika tudománya
Genetikai értelemben egy populációba egy faj azon egyedei tartoznak, amelyek tényleges szaporodási közösséget alkotnak. A populáció génállományát a populációban lévő allélok összessége adja. A különböző allélok különböző gyakorisággal fordulnak elő.
Megváltoznak-e az allélgyakoriságok az egyes nemzedékekben?
A populációgenetika statisztikai módszerekkel dolgozik, ami szerint a statisztika a véletlen tömegjelenségekkel, ezek törvényeivel foglalkozik.
A relatív gyakoriság fogalma
A relatív gyakoriság tapasztalati véletlen mennyiség. Egy érték relatív gyakoriságát úgy kapjuk meg, hogy abszolút gyakoriságának értékét elosztjuk a sokaság elemszámával. A relatív gyakoriság azt mutatja meg, hogy egy adott érték az összes elem hányad részét alkotja.
A relatív gyakoriság 0 és 1, illetve 0% és 100% közötti értékeket vehet fel. A relatív gyakoriságok összege mindig 1, illetve 100%.
Genotípus- és allélgyakoriságok, a populációk genetikai összetételének leírása
Egy populáció genetikai összetételét a populációban előforduló
Egy két alléllel rendelkező autoszómás gént vizsgálva legyen
A D, H, R genotípus-gyakoriságok megállapításához csak arra van szükség, hogy minden egyednek egyértelműen meg tudjuk állapítani a genotípusát.
A genotípus-gyakoriságok ismeretében könnyen kiszámolhatjuk az egyes allélek relatív gyakoriságait is.
Az A allélek az AA homozigótákban és a heterozigótákban fordulnak elő;
Ezért
Példa: egy populáció 100 egyedből áll,
Mennyi az A allél és az a allél relatív gyakorisága?
p = D+H/2 = 0,5 q = R+H/2 = 0,5
Felmerült kérdés: a domináns allél kiszorítja-e a recesszív allélt a populációból? Azaz, megváltoznak-e az allélgyakoriságok az egyes nemzedékekben?
Mi a helyzet a következő generációban?
Tételezzük fel, hogy a vizsgált populáció ún. ideális populáció, ahol
Egy populációban az allélok és a genotípusok gyakoriságának a megoszlását az alábbi egyenlet írja le:
p2 AA + 2pq Aa + q2 aa = 1
Az egyenletben
Ha a populációban a párosodás véletlenszerű, akkor az allélgyakoriságok alapján megadhatjuk a következő nemzedék allélgyakoriság értékeit.
Egy példa: Kiindulási populáció alléljeinek relatív gyakorisága:
Hardy-Weinberg-szabály
Az ún. ideális vagy zárt populációkra vonatkozó törvényszerűség, amely kimondja, hogy az ilyen populációkban az allélek relatív gyakorisága nemzedékről nemzedékre nem változik.
A Hardy-Weinberg-szabály gyakorlati alkalmazása igen széles körű; a fenotípusok megoszlásának ismeretében bármely természetes populációban meghatározható az allélek gyakorisága, valamint a genotípus megoszlás is.
A domináns-recesszív öröklésmenetet mutató jellegeknél nem lehet közvetlen leszámolással megállapítani a három genotípus gyakoriságát, hiszen a domináns homozigótákat fenotípusuk alapján nem lehet megkülönböztetni a heterozigótáktól:
Ilyenkor feltételezzük, hogy a populáció Hardy-Weinberg-eloszlású, és az R = q2 összefüggésből határozzuk meg a q (illetve p = 1-q) allélgyakoriságot.
A népesség 85%-a Rh +. Milyenek a tulajdonságot meghatározó, egyes allél gyakoriságok? Hány % a heterozigóták és a homozigóta dominánsok aránya?
A mendeli öröklődés (meiózis és a gaméták véletlenszerű kombinációja) az allélgyakoriságot nem változtatja meg (Hardy-Weinberg-törvény). Az allélgyakoriság akkor változhat meg, ha a fenti feltételek valamelyike nem áll fenn.
A domináns allél kiszorítja-e a recesszív allélt a populációból? Nem.
Egy nagy létszámú populációban, ha nincs ki- és bevándorlás, szelekció, mutáció stb. (ideális populáció) az allélgyakoriság állandó.
A természetes – reális – populációkra ez nem feltétlenül érvényes (ha mégis, akkor az ellentétes hatások kiegyenlítik egymást, a populáció ekkor is genetikai egyensúlyban van):
Emelt szintű biológia érettségi számítási feladatainak gyűjteménye
2025 október
XI. A Mechanikai érzékelés
A hallás érzékenysége
6. Számítsa ki, hogy mennyi időbe telik, mire az ingerület a szőrsejtektől a hallókéregig eljut! Az ingerületvezetés sebességét tekintsük 100 m/s-nak, a szinaptikus késést 0,5 ms-nak, a hallóideg és a hallópálya együttes hosszát pedig 10 cm-nek. A számítás során 4 szinapszist vegyen figyelembe! Rögzítse a számítás menetét is! (2 pont)
Megoldás
2025 május
II. Éhség és jóllakottság
6. A leptint kódoló génen belül létrejöhet egy olyan pontmutáció, amely teljesen megakadályozza az egyedben a leptintermelést. A DNS-ben egy arginint kódoló bázishármas egyetlen bázisa módosul úgy, hogy a bázishármas STOP kodonná alakul át, ezért hamarabb leáll a fehérjeszintézis. Ha a DNS-ben megtalálható bázisokból véletlenszerűen egy olyan bázishármast rakunk össze, amely egy aminosavat kódol, mekkora az esélye annak, hogy az arginint fog kódolni? Használja a kodonszótárat! Ha a választ tizedestörtben fejezi ki, három tizedesjegyre kerekítve 0–1 közötti valószínűségi értéket adjon meg!
 |
Megoldás
0,098 (6/61), mivel 6 bázishármas kódol arginint, és 61 bázishármas kódol aminosavat (a 3 stop kodon nem kódol aminósavat) 1 pont
IX.A. Idegjelenségek
A nyugalmi potenciál és a kémiai szinapszis
5. Számítsa ki, hogy a központi idegrendszerben elhelyezkedő neuronon megjelenő ingerület keletkezésétől számolva mennyi időre van szükség ahhoz, hogy a ziccert kivitelező vázizomroston kialakuljon az akciós potenciál! A kosárlabdázó ugrása során az akciós potenciálnak 1,8 métert kellett megtennie, részben a piramis pályán, részben a lábba vezető gerincvelői idegen. Feltételezzük, hogy az idegrostokon végig 100 m/s sebeséggel halad az ingerület, a szinapszisokban lejátszódó folyamatok 1,3 ms-ot vesznek igénybe.
Számításának menetét írja le, a végeredményt ms mértékegységben adja meg! (2 pont)
Megoldás
2024 május
IV. Piros a szemed!
Öröklött oka is lehet annak, hogy a szem piros. Ezt az ember esetében egy testi kromoszómán található gén recesszív allélja okozza (albinizmus). Ennek következménye a melanin hiánya, az illető albínó.
4. Az albinizmus egy populációban 1:20000 gyakorisággal fordul elő. Adja meg a mutáns (albinó) allél gyakoriságát! A végeredményt három tizedes pontossággal adja meg, a számolás menetét is tüntesse fel! Feltételezzük, hogy a populáció ideális.
5. Amennyiben az 1,7 millió lakosú Budapest az albinizmus szempontjából ideális opuláció, úgy hány fő lehet hordozó (heterozigóta) a fővárosban? A végeredményt egész számra kerekítve adja meg, és a számolás menetét is írja le! (3 pont)
Megoldás
4.
Ideális populációban: p2+2pq+q2=1, ahol p és q= allélgyakoriság,
q2 a homozigóta recesszívek, azaz az albinók aránya: 1/20000=0,00005
tehát q=0,007 1 pont
5.
A 23630 és 23800 közötti értékek elfogadhatók a kerekítés miatt. Ha a 4. feladatban rossz végeredményt kapott, de azzal ebben a feladatban helyesen számolt, akkor ezen pontok megadandók.
2023 október
IV. Izomdisztrófia
A Duchenne-féle izomdisztrófia fokozatosan súlyosbodó, izomsorvadással járó betegség. Az X kromoszómához kötve, recesszív módon öröklődik.
6. Mekkora a valószínűsége annak, hogy egy, a hibás allélt hordozó anyának születendő fiú-, illetve leánygyermeke izomdisztrófiában fog szenvedni? Az apa egészséges fenotípusú. A mutáció lehetőségétől ebben az esetben tekintsünk el. (2 pont)
7. Adja meg a mutáns allél gyakoriságát, ideális populációt feltételezve egy olyan népességben, melyben a betegség átlagosan 1 esetben fordul elő 3500 fiúgyermek között!
Megoldás
6.
7.
2023 május
III. Teszt-statisztika 10 pont
Olvassa el az alábbi internetes forrás szövegét, és ennek alapján oldja meg a feladatokat!
„Az interneten rengeteg tanács kering. Néha a legnagyobb jóindulattal összeállított listába is bekerülhet egy-egy tévhit. Az egyik ilyen: “Érdemes akár félévente Lyme-tesztet végeztetni, akkor is, ha nem vettünk észre kullancscsípést.” […] A Lyme-kór a kullancsok által terjesztett bakteriális betegség, mely sokszor észrevétlen, de kezelés nélkül néha súlyos következményekkel járhat. A kezelés hosszú antibiotikum-kúrát jelent. Becslések szerint 1000 emberből átlagosan egy a fertőzött. Hiba azonban összekeverni a Lyme-betegséget a pozitív teszteredményt adó laboratóriumi lelettel. A fertőzöttséget kimutató általánosan használt, ún. szerológiai (immunválaszon alapuló) tesztek soha nem tökéletesek. Egy teszt szenzitivitásának azt szokás nevezni, hogy a beteg embert mekkora valószínűséggel minősíti tényleg betegnek a teszt. A teszt specificitásának pedig azt nevezzük, hogy a valóban egészségest mekkora valószínűséggel minősíti egészségesnek. Egy 80%-os szenzitivitású teszt például 100 betegből 80 esetben ad pozitív eredményt (jelzi a fertőzöttséget), a 80%-os specifitású teszt pedig 100 egészséges személy vizsgálata során 20 esetben pozitív eredményt ad (tehát tévesen azt jelzi, hogy a személy fertőzött). […] A Lyme kimutatására szolgáló tesztekkel nagyjából 90%-os szenzitivitás és 99%-os specificitás érhető el. […]”
A leírás alapján egészítse ki a hiányos szöveget!
Tegyük fel, hogy a tesztet véletlenszerűen kiválasztott százezer emberrel végzik el (akár vannak tüneteik, akár nincsenek). Közülük az előzetes becslés adatát elfogadva ………………… (1.) fő az, aki valóban fertőzött. A fertőzöttek közül a teszt 90%-os szenzitivitása miatt csak …………………. (2.) személyt fog betegnek minősíteni, a többi fertőzött immunrendszere nem termelt elegendő ………………………… (3.) ahhoz, hogy azt a teszt kimutassa. A szűrésbe bevontak közül a valóban egészséges személyek száma eszerint ………………….. (4.) fő, a teszt 99%-os specificitása miatt azonban közülük tévesen ………………………. (5.) személyt minősít fertőzöttnek a teszt. (Ezekben a személyekben valószínűleg egy tünetmentesen lezajlott és gyógyult fertőzés emléke ad pozitív eredményt.) Összességében a teszt által betegnek minősített személyek száma ………………………… (6.) lesz, tehát a teszt által betegnek minősített embereknek mindössze ………………… (7.) %-a az, aki ténylegesen beteg. (Egy tizedesjegyre kerekítsen!)
Megoldás
1. 100 1 pont
2. 90 1 pont
3. antitestet / ellenanyagot / immunglobulint 1 pont
4. 99900 1 pont
5. 999 (= 99900 ∙ 0,01) 1 pont
6. 1089 (= 999 + 90) 1 pont
7. [90/(90+999) = 0,0826, azaz] 8,3 (%) 1 pont
Elfogadható a [100/(90+999) = 0,0918, azaz] 9,2 (%) eredmény is. A görgetett hibák halmozódásának elkerülése miatt az 5. válaszban a hibás 4. válasz eredményének 0,01-szerese is elfogadható. A 6. válaszban a hibás 2. és 5. válasz eredményének összege is elfogadható. Így a helyes elvért/összefüggésért kapja az 1-1 pontot.
2022 október
VII. A sarlósejtes vérszegénység elterjedése
A sarlósejtes vérszegénység homozigóta recesszív formája súlyos, heterozigóta változata enyhébb tüneteket okoz. A sarlósejtes vértestek molekulái nagyobb megterhelés esetén elégtelenül kötik meg az oxigént. A sarlósejtes vérszegénységben szenvedő emberek vértestjeit nem képes megfertőzni a malária a kórokozója.
3. Mi a valószínűsége annak, hogy két, a sarlósejtes vérszegénység szempontjából heterozigóta ember házasságából egészséges gyermek szülessen?
5. Számítsa ki az ábra alapján, hogy hány heterozigóta sarlósejtes vérszegény ember él az 5 milliós lakosságú angolai Luandában! A térképen szereplő HbS allélgyakoriságok átlagával számoljon! Luandában a feltüntetett skála legsötétebb árnyalatát vegye figyelembe. Rögzítse a számítás menetét is! (2 pont)
Megoldás
3. 0,25 / 25% 1 pont
5.
A Normál Hb allél gyakorisága: p = 1 – q = 0,8358 1 pont
Heterozigóták száma: 1372384 vagy kerekített értékkel számolva: 1 372 500. 1 pont
2022 május
II. Védendő kártevők
Kedvező körülmények esetén a hörcsög évente akár háromszor is szaporodhat. A szaporodási időszak áprilistól júliusig tart. Egy-egy nőstény 4-16 kölyköt hoz a világra. A kicsik csupaszon és vakon születnek, de a szoptatás hatására elég gyorsan fejlődnek, testüket már pár naposan selymes piheszőr borítja. Az 1990-es évekig kötelező volt a gazdáknak hörcsöggyérítést végezni abban az esetben, ha hektáronként legalább 2 lakott járatot találtak. A hörcsögök túlszaporodása, amely 5-10 évenként tapasztalható, néha napjainkban is komoly gondot okoz. 2017-ben Heves megye egyes területein is nagy egyedszámot mértek, ezért a Bükki Nemzeti Park munkatársai csapdákkal fogták be a hörcsögöket, és a kezelésükben álló, nem mezőgazdasági területeken engedték szabadon az állatokat.
7. 1990-ben egy Békés megyei gazda 10 hektáros búzaföldjén hörcsögök éltek, és az év elején a járataik száma éppen elérte a kötelező gyérítés határát. Mennyi utódot hoztak volna létre a gazda földjén ebben az évben azok az egyedek, amelyek év elején a járatokban éltek, ha feltételezzük, hogy mindegyik járatban minden elléskor a lehetséges maximális számú utód született és minden járatnak egy nőstény lakója volt? Válaszát számítással indokolja!
Megoldás
(10x2x3x16 =) 960 egyed 1 pont
(termőföld területe hektárban x járatok száma x alomszám x almonkénti utódszám)
Csak az a válasz fogadható el, melyben a levezetés mind a négy eleme szerepel, a szöveg szerinti megfelelő értékekkel. Az alomszám tekintetében a maximális 3 mellett az 1 és 2 is elfogadható.
III. Orgonasípok, emberi hangok
Az 1. ábra a belső fül 30 mm-es csigájának felső és alsó járatát kitekert formában ábrázolja, feltüntetve a különböző frekvenciákra érzékeny szakaszok helyét.
A hang terjedési sebessége (c) adott körülmények között állandó, 340 m/s. A rezgés frekvenciájának (f, mértékegysége 1/s = Hz) és hullámhosszának (λ, mértékegysége m) szorzataként számítható ki. 𝑐 = 𝜆 ∙ 𝑓
A Zirci Ciszterci Apátság bazilikájában található orgona sípjai között a legkisebb 1 cm, míg a legnagyobb óriás, 5 méteres. Az orgonában található úgynevezett nyitott sípokban keletkező alaphang hullámhossza kétszerese a síp hosszának (2. ábra).
6. A csiga felső járatának kezdetétől hány mm-re vált ki depolarizációt a csigában egy 4,25 cm hosszú sípban keletkező alaphang? Számításának menetét rögzítse! (2 pont)
 |
Megoldás
V. Szívciklus
A mellékelt diagram egyetlen szívciklus alatt lejátszódó nyomás és térfogatváltozásokat ábrázolja az aortában, a bal kamrában és a bal pitvarban. A szisztolé alatt a pitvarok, majd a kamrák izomzata húzódik össze (a diagramon ez a bal kamra). A diasztolé alatt a kamra izomzata elernyed. Az A, B, C és D betűk szívbillentyűk nyitódásának, illetve záródásának pillanatait jelölik.
 |
5. Számolja ki és adja meg a keringési perctérfogatot, feltételezve, hogy a szív összehúzódásainak ritmusa és erőssége folyamatosan a grafikonon jelzett marad! Rögzítse a számítás menetét!
Megoldás
120–50 = 70 cm3 vér egy összehúzódás során (70-75 cm3 közti értékkel számolhat),
szívfrekvencia: 100/perc, ezért: 7000 cm3/perc (7 dm3/perc) vér a keringési perctérfogata 1 pont
X. A A hemoglobin és a vörösvértestek
A béta-thalassemia a vörösvértestek testi kromoszómán öröklődő betegsége. A betegség súlyos formájában a homozigóta recesszív csecsemők féléves korukra súlyos vérszegénység tüneteit mutatják, mert a hemoglobin béta-láncainak szintézise zavart szenved, emiatt a vörösvértestek feloldódnak. A szervezet fokozott vérképzéssel reagál. A gyermekek koponyacsontjai megvastagszanak, a gyermekek növekedése, fejlődése elmarad.
5. Delhiben felmérték a béta-thalasssemiában szenvedő betegek előfordulási gyakoriságát. 5408 személyből 296 bizonyult betegnek. Tekintsük az öröklést egygénesnek, a vizsgált populációt egyensúlyinak e betegség szempontjából! Határozza meg a betegséget okozó allél gyakoriságát! Írja le a számolás menetét! Az eredményt százalékban adja meg, egy tizedesjegy pontossággal!
6. Adja meg, hogy két, a jelleg szempontjából egészséges fenotípusú ember házasságából mekkora valószínűséggel születik béta-thalassémiás gyerek Delhiben! Írja le a számolás menetét! Az eredményt százalékban adja meg, egy tizedesjegy pontossággal! (3 pont)
Megoldás
5.
296/5408 = 0,0547 = q2 q = 0,234 23,4% 1 pont
6.
Helyesen felírt egyenlet esetén mindhárom pont jár.
2021 október
I. Növényi meiózis
A lúdfű egy testi sejtjében 5 pár kromoszóma van.
8. Adja meg, hányféle különböző sejt jöhet létre a lúdfű egy sejtjének meiózisa eredményeképpen, ha feltételezzük, hogy a gének kapcsoltsága nem változik! Rögzítse a számítás menetét is!
Megoldás
25 = 2·2·2·2·2 = 32 1 pont
VI. Sportélettani vizsgálat
Egy kísérleti személytől nyugalmi állapotban vért vettek, megmérték a pulzusát, valamint szív UH (ultrahang) vizsgálattal meghatározták a bal kamra által kilökött vér verőtérfogatát. Megállapították, hogy a verőtérfogat: 70 cm3. A vizsgált személy pulzusszáma átlagos érték volt, 72/perc.
1. Hány dm3 a kísérleti alany keringési perctérfogata? Rögzítse a számítás menetét! Az eredményt 2 tizedesjegy pontossággal adja meg!
Egy szívhez közeli nagyartériából vett vérmintában az oxigén mennyisége: 200 ml O2/liter vér érték volt, egy, a szívhez közeli vénából vett vérben: 150 ml O2/liter vér.
2. Számolja ki, hogy jelen esetben mennyi a nagy vérkör által ellátott szervek percenkénti O2-felvétele! A számolás menetét is írja le!
Ezután következett az ún. terheléses vizsgálat – intenzív fizikai munka után ismét elvégezték az előző méréseket. A kísérleti személy pulzusszáma 2,1-szeresére nőtt. A szívultrahang szerint a verőtérfogat: 100 ml-re (100 cm3) nőtt.
3. Hány dm3 most a keringési perctérfogat? A végeredményt 2 tizedesjegy pontossággal adja meg! A számolás menetét is írja le!
A második vizsgálat során egy nagyartériából vett vérben az oxigén mennyisége: 200 ml O2/liter vér érték volt, egy, a szívhez közeli vénából vett vérben: 100 ml O2/liter vér.
4. Számítsa ki, hogy hányszorosára nőtt az oxigénfelhasználás a nyugalmi állapothoz képest! A számolás menetét is írja le!
Megoldás
1.
70 ∙ 72 = 5040 cm3 = 5,04 dm3 1 pont
2.
1 dm3 vérből: 50 cm3
5,04 dm3 vérből: 50 cm3 ∙ 5, 04 = 252 cm3 (ml) O2 1 pont
Ha az előző feladatban rosszul számolt a vizsgázó, s azzal a számadattal egyébként jó lenne a megoldás, jár a pont.
3.
pulzusszám: 72 ∙ 2,1 = 151,2 / perc
100 ∙ 151,2 = 15120 cm3 (ml) 15, 12 dm3 1 pont
Ha a pulzusszámot rosszul számolta ki a vizsgázó, de az adattal jól számolt tovább, jár a pont.
4.
1 dm3 vérből: 100 cm3
15, 12 dm3 vérből: 100 ∙ 15,12 = 1512 cm3 (ml).
1512/252 = 6,0 Hatszorosára nőtt. 1 pont
Ha az előző feladatból származó rossz adattal számol, s azzal a számadattal egyébként jó lenne a megoldás, jár a pont.
VII. Öröklődő izomgyengeség
Az ún. perifériás neuropátiák az idegek károsodása miatt alakulnak ki, ami genetikai okból is bekövetkezhet. Ezen betegségek előfordulási gyakorisága 1/2500, ezek közül is leggyakoribb (az esetek 80%-a) a Charcot-Marie-Tooth 1A típusú betegség (CMT1A), amelyet a 17-es kromoszómán elhelyezkedő PMP22 gén megkettőződésével kialakult allél okoz.
A mellékelt családfa egy olyan családról készült, amelyben előfordult a CMT1A betegség (sötét színnel jelölve).
 |
A vizsgált esetben a hibás allélt tekinthetjük egy gén egyetlen domináns hatású alléljének.
7. Adja meg, hogy mekkora volt a valószínűsége, hogy az E-F szülőpárnak mindkét gyereke beteg legyen! Válasza indoklásához adja meg a szülők genotípusát! A hibás (duplikálódott) allélt jelölje „A” és az egészségest „a” betűvel! A 7-8. feladatoknál tekintsünk el a mutáció lehetőségétől! (3 pont)
8. Hazánk lakossága jelenleg kb. 9800000 fő. A valószínűségi adatokat figyelembe véve hány olyan személy él Magyarországon, aki a CMT1A betegségben szenved?
Megoldás
7.
8.
2021 május
V. Sziszüphoszi feladat
9. Számítsa ki, hogy a 80 kg-ot nyomó Sziszüphosz egyik vádlijában mekkora erő ébred abban a pillanatban, amikor talpa felemelkedik a földről! (Ekkor az erővonalak a vízszintes talpra merőlegesen hatnak.) Az ábrán a T betű a lábujjhegyre álláskor a talp leghátsó alátámasztási pontját jelöli, U Sziszüphosz lábujjhegyét, B a bokán áthaladó szaggatott vonal a Sziszüphoszra ható gravitációs erő hatásvonalát jelöli, a V pedig a vádli erőhatásának vonalát. A T, U és B esetében tekintsen el az egyéb szövetek jelenlététől. A 3. ábra bal oldalán látható számadatok centiméterben megadott távolságok. Számításának eredményét két tizedesjegy pontossággal adja meg! (2 pont)
A feladat megoldása során felhasználható további információk:
 |
Megoldás
A Sziszüphosz egy vádlija által emelt súly: G = m·g/2 = 80·9,81/2 = 392,4 N
A súlyból fakadó forgatónyomaték: 392,4 N ·14 cm 1 pont
A vádli által kifejtett forgatónyomaték megegyezik a fenti értékkel: F·20 = G·14
A vádliban ébredő erő (F) értéke: F = 392,4·14/ (14+6) = 274,68 N 1 pont
Amennyiben a vizsgázó a vádlierővel belső erőként számolt, ezért figyelembe vette a súlyhoz hozzáadódó ellenerőt is és a (G + Fvádli) ∙ k1 = Fvádli ∙ k2 (800 + Fvádli) ∙ 14 = Fvádli ∙ 20 Fvádli = 1866 N összefüggéssel számolva helyes eredményre jutott, akkor válaszát teljes értékűként kell elfogadni.
Egyéb helyes levezetés is elfogadható. A második pont csak akkor adható meg bármilyen levezetés esetén, ha a pontosság megfelelő.
VIII. A Fabry-kór
A Fabry-kór ritka öröklődő betegség, amit egy enzim, az alfa-galaktozidáz hibája okoz. A Fabry-kór recesszíven, az X kromoszómához kapcsoltan öröklődik.
A Fabry-kórt okozó mutáns allél gyakorisága Magyarországon 8·10-6. Ha a populációt ideálisnak tekintjük, adja meg, hogy mekkora a betegek gyakorisága férfiak és nők között Magyarországon!
Mekkora
8. A Fabry-kóros betegek gyakorisága a férfiak között?
9. A Fabry-kóros betegek gyakorisága a nők között?
Megoldás
8. q = 8 ·10-6 1 pont
9. q2 = 64 ·10-12 = 6,4 · 10-11 / 1 pont
Az enyhe tünetek nem okoznak valódi megbetegedést, de elfogadható a q2 + 2pq = 1,6 ∙ 10-5 válasz is.
2020 május
VII. Hajhullás
Az emberi haj az emlősök szőrének speciális megjelenési formája. Az ember hajas fejbőrének átlagos területe 770 cm2, ami nagyjából 100.000 hajhagymát tartalmaz. Fejlődésük jellegzetes stádiumokon megy végbe, az alábbi fázisok ciklikus váltakozásával:
 |
6. A megadott adatok alapján számítsa ki, hogy egy nőnek naponta átlagosan hány szál haja hullik ki (egész számra kerekített értéket adjon meg)! A számítás menetét is tüntesse fel! (Egy hónap hosszát tekintse 30 napnak; ha az átlagos értékekkel kell számolnia, akkor a szövegben szereplő határértékek átlagát vegye alapul! Egy hajhagymából egy hajszál nő ki.)
Megoldás
100000·0,125/(3·30) = 138,9 ~ 139 szál 1 pont
Az alábbi számítás is elfogadható:
2019 október
V. Élesztőkísérlet
A tanulók az élesztő lebontó anyagcseréjét vizsgálták. Élesztőt tettek cukoroldatba és a lombikhoz egy kalibrált fecskendőt rögzítettek az ábrán látható módon. Ezt követően egy órán keresztül minden 10 percben megmérték a fejlődő gáz teljes mennyiségét. A fejlődő gáz a fecskendő dugattyúját elmozdítja. A rajz a kísérlet elvi vázlatát mutatja, nem méretarányos.
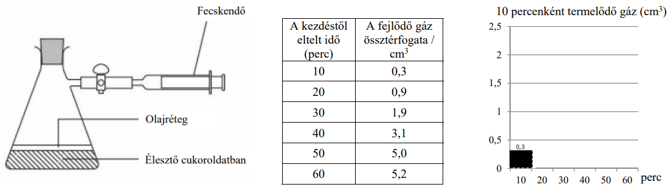 |
Ábrázolja az időegységenként (10 perc alatt) fejlődő gáz mennyiségét a mellékelt oszlopdiagram folytatásával! Az oszlopok fölött tüntesse fel az értékeket is!
Megoldás
A táblázatban a fejlődő gáz össztérfogata szerepel, ezért a 10 percenként fejlődő gázt úgy tudjuk kiszámolni, hogy az adott időpontban mért össztérfogatból kivonjuk az előző mérés össztérfogatának értékét.
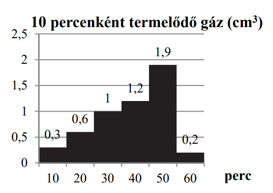 |
VII. Valószínűségek
1. Júlia és Péter gyermeket várnak. Az ultrahang vizsgálat során kiderült, hogy fiú. Genetikai tanácsadóhoz fordulnak, mert szeretnék tudni, hogy mekkora az esélye annak, hogy színtévesztő lesz. Mindketten normál színlátásúak, de Júlia édesapja színtévesztő. A genetikai tanácsadó válaszát számításokkal indokolja! A helyes színlátás allélját jelölje „A”, a színtévesztés allélját „a” betűvel! (2 pont)
2. Írja le, hogy hogyan módosulna az előző ponthoz képest a válasza, ha Júlia szüleiről nem állna rendelkezésére információ, tudja viszont, hogy ebben az elképzelt, és ideálisnak tekintett népességben a nők 0,5%-a színtévesztő. Számítását írásban rögzítse!
Az allélgyakoriságokat négy tizedes, a végeredményt százalékos formában, egy tizedesjegy pontossággal adja meg! (3 pont)
Megoldás
1.
A genotípusok
A gyermekek lehetséges genotípusai és ezek gyakoriságai:
 |
Másképpen is levezethető.
2.
Elfogadható az alábbi gondolatmenet is:
2018 október
III. Csillósok egymás közt 11 pont
F. Gause orosz kutató híres kísérletsorozatában két, közös élettérben élő papucsállatka faj egyedszámának változását követte nyomon 1934-ben. Más papucsállatkákhoz hasonlóan e fajok is főként baktériumokkal táplálkoznak. Gause minden nap azonos mennyiségű, baktériumokban gazdag oldat adagolásával biztosította ellátásukat.
4. Gause kísérletét Violle és csoportja folytatta, több mint 60 évvel később. Ők 10 különböző csillós egysejtű együttélésének lehetőségét próbálták ki, páronként vizsgálva minden lehetőséget. Adja meg, hogy összesen hányféle kombinációt vizsgáltak!
 |
A 3. ábra fekete pontjai azt mutatják, hogy a kihalás mennyi idő múlva következett be. Minden pont különböző tenyészetet jelöl.
8. Adott körülmények között a lehetséges kombinációk hány százaléka eredményezett tartós (kihalással nem járó) együttélést a két faj között? Egész számra kerekítve adja meg az eredményt! A számolás menetét is írja le!
Megoldás
4. 45 (45 = 9+8+7+6+5+4+3+2+1. A számítást nem kell feltüntetni.) 1 pont
8. 23/45 = 0,5111, kerekítve 51%. (45 helyett a 4. részfeladat eltérő megoldásával végzett számolás eredménye is elfogadható.) 1 pont
2018 május
III. Női dolgok
Egy egészséges, 38 éves nőtől 28 napon keresztül átlagosan négynaponta vért vettek és megállapították benne négy, a nemi működést befolyásoló hormon koncentrációját. A mért adatokat a következő táblázat tartalmazza. (A hormonkoncentráció ng/ml dimenzióban szerepel a táblázatban.) A petefészek hormonjainak koncentrációi sohasem érik el a 10 ng/ml-t.
 |
1. Számítsa ki, hogy a következő hormonok maximális szintje hány százalékos eltérést mutat a minimálishoz képest a ciklus során! A legnagyobb értéket tekintse 100 %-nak, eredményét egy tizedesjegy pontossággal adja meg! (2 pont)
a) I. hormon:
b) IV. hormon:
Megoldás
1.
a) 77,8 % (77,7 % is elfogadható.) 1 pont
b) 98,7 % (98,6 % is elfogadható.) 1 pont
(A megfelelő pontossággal megadott eredményekre adható pont.)
IX. B. Választható feladat – „Kis-Kunságnak száz kövér gulyája”
Legelők energiahasznosítása
Hogy hány „gulyát” lehet legeltetni az Alföld rónáin, az többek közt attól függ, hogy a Napból érkező sugárzás mi módon hasznosul a legelő ökoszisztémájában. Mért adatok alapján a legeltetés időszakában a hazai legelők irányába naponta átlagosan 3,35 kJ/cm2 napenergia érkezik. Ennek azonban csak 23%-a éri el a fotoszintézisre képes növények levélzetét.
A pigmentek által elnyelt energia egy része a fotoszintézis biokémiai folyamataiban nem hasznosul, hanem hővé alakul, s csak egy kisebb hányada alakul a növényben létrehozott vegyületben tárolt kémiai energiává. Mindazonáltal a cukor formájában megkötött energia sem kerülhet be teljes egészében a táplálékláncba, mivel annak jelentős részét lebontja maga a növény. A bemutatott veszteségek miatt a levélfelületre érkező fényenergiának mindössze 5,4 %-a kerül tartósan a termelőkbe, azaz alakul a legelő táplálékláncába belépő elsődleges produkcióvá.
Az ökoszisztémában zajló energiaáramlás néhány számadatának birtokában akár a gulyások is kiszámíthatják, hogy mekkora legelő tartja el gulyájukat.
6. Számítsa ki, hogy elvben (pusztán az energiaforgalom megadott számai alapján) hány hektáros az a kiskunsági legelő, amelyik képes eltartani a versben szereplő „száz kövér gulyát” (a számításban: egy 100 marhából álló gulyát), ha tudjuk, hogy egy szarvasmarha napi átlagos energiaigénye 150000 kJ, és a növények elsődleges produkciójának átlagosan 12 %-át hasznosítják a legelésző állatok! (1 hektár = 10000 m2) A számításokat három értékes jegy megtartásával végezze, s az eredményt is ennek megfelelően adja meg. (3 pont)
Megoldás
6.
A gulya napi energia-igénye: 150000 ∙ 100 = 1,500 ∙ 107 kJ
ehhez igényelt napi elsődleges produkció: 1,5 ∙ 107 / 0,12 = 1,25 ∙ 108 kJ 1 pont
A levélfelületre naponta érkező energia: 1,25 ∙ 108 /0,054 = 2,315 ∙ 109 kJ 1 pont
A naponta érkező napenergia mennyisége: 2,315 ∙ 109 / 0,23 = 1,006 ∙ 1010 kJ
A szükséges legelőméret:
1,006 ∙ 1010 / 3,35 = 3,00 ∙ 109 cm2 = 30,0 ha / 30 ha 1 pont
Egyéb helyes levezetésért is megadható a pontszám, részpontszámok – számítási hiba esetén az egyes sorokban szereplő összefüggések helyes alkalmazására adhatók. Elvileg helyes részlépésekre részpontok adhatók, de elvileg hibás levezetésre nem adható pont.
2017 október
VII. Karizom
A súlyzós gyakorlatok során a kar anatómiai egységei egyoldalú (egykarú) emelőként működnek. Az ábra az „emelő” azon állapotát mutatja be, amikor az alkar kb. 90 fokos szöget zár be a felkarcsonttal (a talaj az ábra alja felé helyezkedik el). A vastag nyilak erőket, a vékonyak távolságokat jelölnek.
 |
5. Számítsa ki, hogy a 15 kg-os súlyzó adott helyzetben tartásához mekkora erőt kell kifejtenie az alkarhajlító izomnak, ha a sportoló alkarjának tömege 3 kg! A számítás menetét is tüntesse fel, az értékeket két tizedesjegy pontosságig, newton mértékegységben adja meg! (2 pont)
A számítás során a következőket vegye figyelembe:
Megoldás
5.
Egyéb helyes levezetésért is megadható a teljes pontszám.
VIII. Hemoglobin és malária
A sarlósejtes vérszegénység testi kromoszómához kötött recesszív tulajdonság. A betegség élettani tünetei feltűnő mértékben csak homozigóta állapotban jelennek meg. A heterozigóták hétköznapi körülmények között nem mutatják a betegség tüneteit és feladatunkban egészségesnek tekinthetők. A mutáns gén szokatlanul nagy arányban fordul elő az afroamerikai emberek körében, ahol minden 655-dik emberre jut egy beteg.
3. Adja meg, mekkora a sarlósejtesség alléljának gyakorisága ebben a populációban! (A számolás során csak a végeredmény megadásakor kerekítsen négy tizedesjegy pontosságra!)
4. Milyen valószínűséggel születik beteg gyermeke annak a párnak, akikről tudjuk, hogy mindketten heterozigóták a sarlósejtes vérszegénység jellegre?
5. Adja meg, milyen valószínűséggel születik beteg gyermeke két egészséges fenotípusú szülőnek a fent leírt közösségben, ha nem ismerjük egyik szülő genotípusát sem! Írja le a számítás gondolatmenetét is! (2 pont)
Megoldás
3. A homozigóták gyakorisága q2 = 1/655= 0,001527, ezért q = 0,0391 (=3,9 %) 1 pont
4. 0,25 / 25% 1 pont
5.
Egészségesek között a heterozigóták aránya:
Beteg gyermek születésének valószínűsége:
Elvileg helyes megoldás és számolási hiba estén: 1 pont.
Ha a 3. kérdésre téves választ adott, de a kapott eredménnyel az 5. feladatban elvileg helyesen számolt a vizsgázó, akkor az 5. feladatra adott válaszát el kell fogadni.
X. B Választható feladat - Védett termőhelyek
Cifra kankalin
Különböző fajok együttes megjelenését úgy jellemezhetjük számszerűen, ha külön-külön vett gyakoriságuk (p és q) szorzatát (pq) összehasonlítjuk azon vizsgálati négyzetek gyakoriságával, melyekben együtt fordultak elő. Ha ez utóbbi szám lényegesen nagyobb, mint pq, akkor a két faj kötődik egymáshoz, ha közel azonos, előfordulásuk egymástól független, ha pedig kisebb annál, kerülik a közös termőhelyeket. (Az 1-25. oszlopokban a tömegességet jelző számokat e számítás során tekintsük egyszerűen + jelnek.) Így a vizsgált esetben a cifra kankalin 10 négyzetben fordult elő (p=10/25=0,4), a tavaszi kankalin 18-ban (q=0,72), szorzatuk (pq = 0,288) nem tér el lényegesen a valóságos együttes értéktől (7/25 = 0,28), azaz a két faj előfordulása egymástól független.
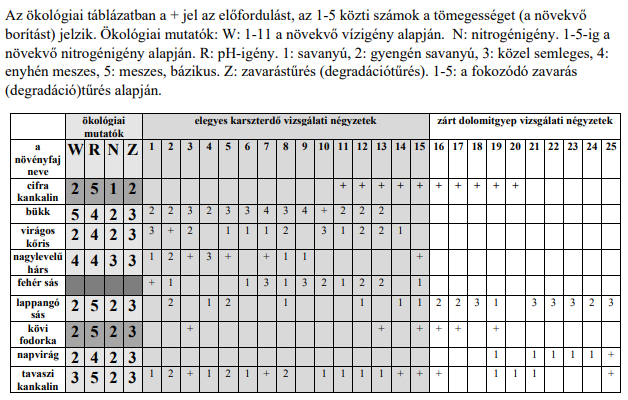 |
5. Hasonlítsa össze a leírthoz hasonló módon a bükk és a cifra kankalin előfordulását! Vonja le a következtetést, és adjon rá valószínű magyarázatot eltérő környezeti igényeik, illetve egy lehetséges populációs kölcsönhatás alapján! (3 pont)
Számítás és következtetés:
Lehetséges kölcsönhatás:
Eltérő környezeti igények:
6. Végezze el a számítást az előzőhöz hasonló módon a kövi fodorka és a cifra kankalin előfordulásai között! Adjon egy lehetséges magyarázatot az összefüggésre! (2 pont)
Magyarázat:
Megoldás
5.
A valóban mért érték: 3/25=0,12, ami jóval kevesebb a számítottnál. A bükk és a cifra kankalin közös előfordulása a véletlenszerűnél ritkább / „kerülik egymást”. 1 pont
A felsoroltak közül bármelyik különbség megfogalmazása. 1 pont
6.
együtt 5/25 = 0,2, ami több mint kétszerese a véletlenszerűnek.
A fajok kötődnek. 1 pont
Más jó javaslat, pl. szimbiózis feltételezése is elfogadható.
2017 május
VI. Vértestvérek
3. Egy szülőpár mindkét tagja A Rh-pozitív vércsoportú. Első gyermekük 0, Rh-negatív vércsoportú lett. Mekkora a valószínűsége, hogy a második gyermekük A és Rh-pozitív vércsoportú lesz? Válaszát levezetéssel indokolja és két tizedesjegy pontossággal adja meg! (2 pont)
4. Örökletes tulajdonságainkban sokszor nem szüleinkre, hanem nagyszüleinkre hasonlítunk. Indokolja, milyen esetben fordulhat elő, hogy az utód valamely tulajdonsága valóban a nagyszülőével egyezik meg, de a szüleivel nem! (A legegyszerűbb, egy gén, kétallélos, domináns-recesszív öröklésmenetet feltételezzen!)
5. Adja meg, hogy az unoka génkészlete legalább hány százalékban azonos valamely nagyszülőével!
8. Nagyjából hány db vörösvértest található egy egészséges felnőtt férfi szervezetében? Számolását rögzítse! (2 pont)
Megoldás
3.
Ha a levezetésből egyértelmű, hogy a szülők heterozigóták, az első pont akkor is megadható, ha ezt nem írta le.
4.
(másképp, jelöléssel illetve konkrét példával is megfogalmazható)
5. 25% 1 pont
8.
(Ha szerepel a helyes vértérfogat vagy vörösvértest-szám adat, 1 pont adható.
Csak a számítás gondolatmentét tükröző megoldásra adható 2 pont.)
2016 október
V. Összeférhetetlen növények
A növényfajok jelentős része esetében kialakultak olyan mechanizmusok, amelyek megakadályozzák az önmegtermékenyítést. Ezek többsége vagy az önmegporzást vagy a megporzást követő eseményeket teszi lehetetlenné.
Az önmegtermékenyítés elkerülésének egyik lehetősége, amikor genetikai hasonlóság esetén a bibe szövetei elutasítják a pollenszemeket (többnyire a pollentömlő pusztulását okozva). Az összeférhetetlenséget egy gén alléljei okozzák. Abban az esetben alakul ki az elutasító reakció, ha a bibe szövetei és a pollenszem ezen gén azonos allélját tartalmazzák. A folyamat következtében egyes egyedek nem képesek egymás megtermékenyítésére. Mivel számos termesztett gyümölcsfajunk (pl. cseresznye, mandula, kajszi) esetében létezik az önmegtermékenyítést megakadályozó allélrendszer, a jelenségnek kertészeti jelentősége is van.
Egy növényfajban a diploid növényi szövetekben az összeférhetetlenséget egyetlen gén alléljei határozzák meg. Egy szaporítási kísérletben kétféle genotípusú egyedet vizsgáltak. Az összeférhetetlenséget meghatározó allélpár egyik tagja mindkét típusban azonos volt (A1), másik tagja viszont különbözött egymástól és az A1-től is. (Jelöljük genotípusaikat A1A2 -nek és A1A3-nak.) A kutatók ebből a kétféle, eltérő genotípusú egyedből egyet-egyet közös növénynevelő kamrában helyeztek el. Virágzáskor biztosították a tetszőleges irányú megporzást a kamrákon belül. A termésérést követően begyűjtötték és elvetették a magokat, majd a kihajtott csíranövényeket genetikai vizsgálatnak vetették alá.
6. Feltételezve, hogy a két növény azonos mennyiségű pollenszemet és magot érlelt, és a csíranövények azonos életképességűek voltak, milyen genotípusúak lettek a csíranövények az A génre nézve?
Számítása során a következő kérdésekre adjon választ: (3 pont)
a) Milyen genotípusú pollenszemeket hozott létre a kétféle növény?
b) Létrejöhetett-e önmegporzással sikeres termésérlelés a leírt kísérlet során? Indokolja állítását!
c) Milyen genotípusú csíranövények milyen arányban jöttek létre? Megoldását vezesse le!
Megoldás
6.
a) a termelődő pollenszemek genotípusai:
b)
c) (Ezért csak kölcsönös megporzás jöhet létre.) A kialakuló genotípusok :
 |
Azaz a kapott genotípusok aránya:
VII. A cisztás fibrózis
A cisztás fibrózis örökletes genetikai betegség. Hazánkban négyezer emberből 1–2 beteg, míg 4–5 százalékunk hordozza a betegséget. A betegség hátterében álló kóros gént 1986-ban találták meg a 7. kromoszóma hosszú karján.
A betegség testi kromoszómához kötött, recesszív módon öröklődik. A betegség "lényege" az, hogy a génkárosodás miatt a kloridion és a víz a mirigyekbe nem vagy alig jut át, így a képződött váladék besűrűsödik, ezáltal a szervekből nem tud rendesen kiürülni, mintegy dugót képez.…
…Erre a betegségre a csecsemőkor második felétől egyre gyakoribb felső és alsó légúti fertőzések (hörghurut, tüdőgyulladás) hívják fel az orvos figyelmét. A légutakban termelődő nyák sűrű, nehezen tud távozni, a nyákon a baktériumok könnyen elszaporodnak, így állandó gyulladást hozva így létre… A másik gond a hasnyálmirigyben a váladék besűrűsödése.
2. Egy egyensúlyinak tekinthető populációban 1:1700 a betegség előfordulásának gyakorisága.
a) Mekkora itt az egészséges és a hibás allél gyakorisága?
b) Hány % valószínűséggel születik ebben a populációban két egészséges embernek beteg gyermeke? Válaszai csak a levezetéssel, számolással együtt fogadhatók el! (2 pont)
3. A tapasztalat szerint a cisztás fibrózist hordozók nagyobb eséllyel élnek túl súlyos hasmenésekkel járó járványokat. Ha a populációban hosszabb időn keresztül tartó kolera – súlyos hasmenéssel járó betegség – járvány lépne fel, hogyan változna meg a recesszív allél gyakorisága?
Megoldás
2.
a) a betegség előfordulásának gyakorisága: q2= 1/1700
a recesszív allél gyakorisága: q=0,024
a domináns allél gyakorisága: p= 1-q = 0,976 1 pont
b) Egészséges szülők esetén beteg gyermek csak heterozigóta pártól születhet.
Az egészséges szülőpár heterozigóta genotípusának valószínűsége: (2pq / p2+ 2pq)2 = 0,0022 1 pont
Két heterozigóta egyén gyermeke ¼ valószínűséggel lesz beteg. Ennek alapján
0,0022*0,25=0,00055. Vagyis 0,055%-os valószínűséggel születik ebben a populációban két egészséges embernek cisztás fibrózisban szenvedő gyermeke. 1 pont
3. Nőne a recesszív allél gyakorisága 1 pont
2016 május
V. Szívciklus
A mellékelt diagram egyetlen szívciklus során lezajló nyomás- és térfogatváltozásokat ábrázolja az aortában, a bal kamrában és a bal pitvarban. A szisztolé alatt a pitvarok, majd – a szaggatott vonalakkal jelölt időszakban – a kamra izomzata húzódik össze (a diagramon ez a bal kamra). A diasztolé alatt a szívizom elernyed. Az A, B, C és D betűk a szívbillentyűk nyitódásának, illetve záródásának pillanatait jelölik. Az első, elnyújtottabb szívhangot a vitorlás billentyűk becsapódása adja, míg a második, rövidebb szívhang a zsebes billentyűk záródásából ered. A diagram tanulmányozása után válaszoljon a következő kérdésekre!
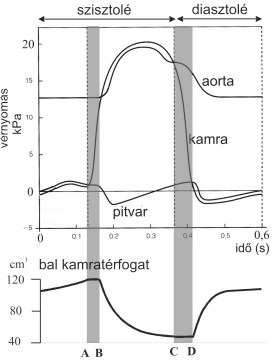 |
1. Hány cm3 vért lök ki a vizsgált személy bal kamrája egyetlen összehúzódása során?
2. Mekkora a vizsgált személy keringési perctérfogata (a bal kamrából a nagy vérkörbe juttatott vér mennyisége 1 perc alatt), feltéve, hogy az összehúzódások ritmusa végig a grafikonon látottat követi?
3. Az 1 perces időtartam alatt hány másodpercig végez munkát a kamra izomzata?
Megoldás
1. 120 - 50 = 70 cm3 (70-75 cm3 közti értékek elfogadhatók.) 1 pont
2. 70 . 100 = 7000 cm3 = 7 dm3 Az 1. pontban számolt érték százszorosa elfogadható. 1 pont
3. A kamra a 0,13 s-tól – 0,36 s-ig terjedő szakaszban húzódott össze, ami 0,23 s, a 100 ciklus alatt tehát 100 . 0,23 = 23 másodpercig dolgozott. (A 18 és 28 s közti értékek elfogadhatók). 1 pont
2015 október
IV. AIDS és pestis
A vérzékenységben szenvedők a betegség kezelésére rendszeresen „VIII-as véralvadási faktor”-injekciókat kaptak. Ezt több ember véréből nyerik. Az AIDS felismerése előtti időben az egyik készítménybe egy AIDS-es véradóból származó vér is belekerült. Emiatt a készítmény AIDS-fertőzést okozott. Mégis voltak olyan egyének, akik nem fertőződtek meg, noha ugyanazt a fertőzött vérkészítményt kapták, mint betegtársaik. A kutatók megállapították, hogy az AIDS-fertőzéssel szemben ellenálló egyének egy mutációt hordoztak a T-sejtek felszínén levő receptort meghatározó génben, a CCR5-ben. Ez a mutáció, a CCRT32 homozigóta formában megakadályozza a kórokozó HIV vírus immunsejtbe jutását, és ezáltal védettséget nyújt. (…)
Az európai populáció 1%-a homozigóta erre a mutációra, ázsiaiaknál és afrikaiaknál nem tudták kimutatni. A CCRT32 európai jelenlétét a gyanú szerint az Európában egykor gyakori pestisjárványok segítették: a mutáns homozigóták a pestissel szemben is védettek voltak. De ezt a sejtést be is kellett bizonyítani. (…)
Az 1665-ös angliai pestisjárvány egy Eyaem nevű kisvárosban is pusztított. A kórt itt egy Londonból érkezett szabó a londoni bolhákkal telített ruhaszöveteivel terjesztette el. George Vicars maga is áldozata lett mesterségének, de nemcsak ő, hanem az akkor nyolcszáz fős faluból 250-en. A falu azért nevezetes, mert a helyi lelkész javaslatára a járvány kitörése után a faluba senkit nem engedtek be, és onnan senkit sem engedtek ki. A járványban elhunytakról pedig gondos listát készítettek. A kutatók a listák alapján megtalálták a túlélők utódait, akikben a CCRT32 mutáns allél előfordulási gyakorisága az átlag többszöröse volt, a homozigóták aránya pedig az átlag kétszerese. Bizonyítást nyert tehát, hogy valóban ez a mutáció okozta a túlélést, a pestis volt az a szelektáló tényező, amely az AIDS-rezisztensek* szerencséjét eredményezte.
2. Számítsa ki, hogy egyensúlyi (Hardy-Weinberg egyensúlyban levő) populációt feltételezve az európai népesség hány százaléka hordozza ma a CCRT32 mutáns allélt (anélkül, hogy maga védett lenne a HIV-fertőzéssel szemben)! (2 pont)
Megoldás
VII. A nádszálak belsejében
A nád szárának belsejében mérték a levegőáramlást. Az eredményeket két egymást követő napon a grafikon mutatja. A „fotonáramsűrűség” a fotoszintézis intenzitásának mértéke.
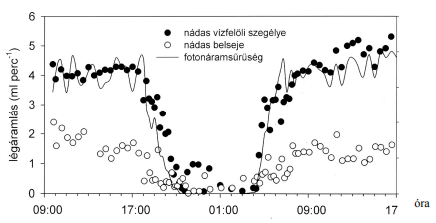 |
5. A Balaton északi partján átlagosan 67 nádszál nő négyzetméterenként. Számítsa ki, hány liter levegőt áramoltat át a nádas négyzetméterenként a szegélyeken, teljes megvilágítás mellett 1 óra alatt!
Megoldás
Mivel percenként kb. 4,5 ml levegő áramlik át egy nádszálon, ezért 1 óra alatt 67 · 4,5 · 60 = 18 090 ml = kb. 18 liter (dm3) levegő áramlik át a nádszálakon. (16 és 21 liter közti értékek elfogadhatók).
IX. A Választható feladatok
Szemtől szembe
A szemben a külvilág kicsinyített, fordított állású képe vetül a retinára. A leképezésben egy lencserendszer (a szaruhártya és a szemlencse) vesz részt, de egyszerűsítve tekinthetjük úgy, mintha egyetlen domború lencse látná el ezt a feladatot. A leképezést (közelítőleg) a lencsetörvény fejezi ki: 1/f = 1/k + 1/t; ahol f = a fókusztávolság, k = a képtávolság (a kép távolsága a lencserendszertől), t = a tárgytávolság. Az optikában a dioptriával fejezik ki a lencse(rendszer) törőképességét, mely a méterben megadott fókusztávolság reciproka (D=1/f).
7. Számítsa ki, hány dioptriásnak adódik a szem lencserendszere, ha “végtelenre” (igen távoli tárgyakra) nézünk! A vizsgált személyben a lencserendszertől a sárgafoltig a távolság 24 mm. (Az eredményt 1 tizedesjegy pontossággal, a kerekítés szabályainak megfelelően adja meg.)
8. Közelre nézéskor a lencse domborúbbá válik. Hány dioptriásra nő a törőképessége, ha egy tőlünk 25 cm-re levő szöveget olvasunk?
9. Hány dioptriás szemüveget (vagy kontaktlencsét) használjon egy 25 cm-re levő szöveg nézéséhez az előző feladatban szereplő ember, ha idős korára távollátó lett: a távoli tárgyakat élesen látja, de szemlencséjének domborúsága már nem változtatható?
Megoldás
7.
D = 1/f = 1/k + 1/t,
mivel t igen nagy, 1/t értéke elhanyagolható, így
D = 1/k = 1/ 0,024 m = 41,7 1 pont
8.
D = 1/f = 1/k + 1/t,
D = 1/ 0,024 + 1/0,25 = 41,7 + 4 = 45,7
A törőképesség 45,7 dioptriásra nőtt. 1 pont
9.
Dszemüveg = 45,7 – 41,7 = 4 dioptriás szemüveget. 1 pont
2015 május
II. Miért pusztul a nád?
A Balatonban az 1970-es évek végén kezdett pusztulni a nád (Phragmites communis). Először a vízfelőli részen jelentek meg csatornák, majd a korábban egyenletes állomány különálló zsombékcsomókra esett szét, melyeket aztán kidöntöttek a hullámok. A nádpusztulás több európai tóban is jelentkezett. (…) Az egyik elmélet a genetikai diverzitás csökkenésének tulajdonított szerepet a pusztulásban. Azt, hogy a nád annyira különböző helyeken él meg, okozhatja az, hogy a klónoknak szűk a tűrőképessége, viszont nagyon sok, különbözőkörnyezethez alkalmazkodott klón van. (Egy klónnak ebben az esetben az egy magból származó egyedek összességét nevezzük.) Az elmélet szerint a magoncokból a víz partján kialakult tarka állományból a gyöktörzsek a víz alatt elindulnak a mélyebb víz felé, de ha összeérnek a klónok, erős versengés indul meg közöttük, és a környezeti viszonyoknak legmegfelelőbb végül kiszorítja az összes többit. Ha ezután megváltoznak a körülmények, és az új helyzetnek nem felel meg ez a genotípus, nincs ami a helyére lépjen, és pusztulásnak indul a nádas. A klónokat a belőlük kivont DNS molekulák összehasonlítása alapján különböztethetjük meg egymástól. Korábban külföldön ezzel a technikával több száz méteres, egy klónból álló állományokat találtak. A kutatók arra voltak kíváncsiak, így van-e ez hazai nádasainkban is.
Egy alsóörsi nádasban ellenőrizték az elmélet érvényességét. Az ábrán látható területről 4*4m sűrűségű rács szerint vettek DNS-mintákat (5 szélső négyzetben nem volt nád). Az azonos klónhoz tartozó egyedeket azonos számmal jelölték. Az ábrán nagyobb és vastagabb számokkal emelték ki azokat a négyzeteket, melyekben azonos klónhoz tartozó egyedek voltak, és a klónok több vizsgálati négyzetre is kiterjedtek.
 |
6. A náddal borított terület hány százalékát borították olyan klónok, amelyek csak egy vizsgálati négyzetre terjedtek ki? (Egy tizedesjegyre kerekítve számoljon!)
7. Átlagosan mekkora területre terjedt ki ezen nádas náddal borított területén egy-egy klón (a vizsgált módszer alapján)?
Megoldás
6.
60 ilyen négyzet volt, tehát
a náddal borított terület (60/95)100 = 63,2%-ára terjedtek ki az ilyen kisméretű klónok. 1 pont
7.
Összesen 75 különböző klón volt,
95/75 = 1,3 mintavételi egység, azaz 1,3 · 16 = 20,8 m2 volt egy-egy klón átlagos kiterjedése. 1 pont
Más számítási elv alapján kapott eredmény is elfogadható a következőintervallumban: 20,1-20,8 m2.
VI. A DNS összetétele és működése
1. A DNS-molekula egy szakasza 850 nukleotidpárból áll. Ezen a szakaszon a citozin a bázisok összmennyiségének 22%-át teszi ki. Határozza meg, hogy az adott DNSmolekula-szakaszban hány db citozin, guanin, timin és adenin található! A számolás menetét is írja le! (4 pont)
Megoldás
Számolási hibáért – ha a gondolatmenet helyes – 1 pont vonható le.
Más gondolatmenet szerinti megoldás is elfogadható.
X.B Erdők
Erdők szerkezete és fajösszetétele
A tanulók az egyes erdőtípusok növényfajait vízigényük alapján csoportosították. Eredményeiket a grafikon mutatja. A nagyobb W-érték a faj nagyobb vízigényét jelzi.
 |
6. Számolja ki ennek alapján az egyes erdőtípusok jellemző vízellátottság-értékeit az eredmények átlagolásával. Az eredményeket írja a táblázat üres celláiba! Az „A” erdőtípusét megadtuk. (2 pont)
 |
Megoldás
 |
2014 október
VII. A bőr
Egy emberi populációban minden tizenötezer ember közül egynél nyilvánul meg e endellenesség. (Tekintsük ezt a populációt erre a jellegre nézve egyensúlyinak!)
5. A populáció hány százaléka egészséges, de egyben a rendellenes allél hordozója? Írja le a számítás menetét is! (4 pont)
Megoldás
2014 május
III. Veseműködés
Egy klinikán a vizsgált személy veséjének vérellátottságát szeretnék vizsgálni, ezért meg kívánják határozni a szerv vérátáramlását. Ehhez a gyakorlatban paraaminohippursavat (PAH) használnak segédanyagként, egy nagyvéna vérébe beadva. A vese ezt az anyagot meghatározott arányban választja ki, a vérplazma PAH-tartalmának átlagosan 90%-a ürül a vizelettel. A mérés során a vizsgált személy veseartériájának vérplazmájában a PAH koncentrációja 1,2 mg/cm3, vizeletében pedig 672 mg/cm3 volt.
1. Számítsa ki a vesén átáramló vér mennyiségét cm3/perc egységben, ismerve, hogy a vérplazma a vér térfogatának 55%-át teszi ki, a napi vizeletmennyiség pedig 1500 cm3!
A vizeletképződés sebességét a nap során tekintsük állandónak! A megoldás során egy tizedesjegyre kerekítve számoljon. (5 pont)
Megoldás
1.
A feladat megoldását egy tizedesjegy pontossággal kellett megadni, egyéb esetben ez pontlevonással jár. Más sorrendű levezetést is el kell fogadni! Nem jelent pontlevonást, ha a részeredmények eltérése nem elvi hiba, hanem a tizedesjegyek eltérő kerekítéséből adódik. Számolási hibáért csak 1 pont vonható le, de a logikailag helyes részlépésekért a pontok megadhatók.
Másik megoldás:
A két megoldás eredménye közti eltérés a kerekített értékkel történő számolásból adódik.
Ha a vizsgázó a PAH-koncentráció csökkenését feltételezte, a következő levezetés is pontozható:
azaz ellentmondáshoz jutottunk.
Az utolsó pont csak abban az esetben adható meg, ha a vizsgázó egyértelműen jelezte számítási eredményének ellentmondásosságát.
VI. Szívverés
Az ábrán egy emberi szívet, és az azon végigfutó ingerületi hullámok következtében az izomösszehúzódás kezdőpillanatait ábrázoltuk. (A pont utáni számok tized-, ill. századmásodperceket jelentenek).
 |
3. Mennyit pihen az ábrán látható szív egy perc alatt, ha percenként 75-szer húzódik össze? Tételezzük fel, hogy a kamrafalak elernyedése 0,08 másodperccel azt követően indul meg, hogy az ingerületi hullám a teljes kamrán végigfutott. (2 pont)
Megoldás
3.
Más sorrendű levezetés is elfogadható.
2013 október
III. A növényvilág hálózatai
8. Számítsa ki, hogy az első elrendezés esetében (a papírnyír megvilágításakor) hány mg glükóz keletkezett a vizsgálat ideje alatt a papírnyír által megkötött 13C-mal jelölt széndioxidból! Számítását a fotoszintézis egyenletével támassza alá!
Mr(13CO2)=45; Mr(13Ctartalmú glükóz)=186.
Megoldás
8.
6 CO2 + 6 H2O = C6H12O6 + 6 O2 1 pont
Mind a bruttó, mind a nettó egyenlet elfogadható.
Ha (6 · 45) mg = 270 mg 13CO2-ból keletkezik 186 mg glükóz, akkor (288 · 0,86) mg = 247,68 mg 13CO2-ból keletkezik 170,62 mg glükóz 1 pont
IV. Agancsképzés és csontritkulás
A szarvas képes arra, hogy az agancsfejlődés időszakában saját magának ideiglenesen csontritkulást okozzon, majd azt visszafordítsa, és újból megerősítse csontjait. A szarvas évente megújítja fejdíszét. Mindazok a tápanyag-összetevők szükségesek az agancstermeléshez, amelyek a csontnövekedésben is szerepelnek. Az új agancs növesztéséhez a szarvasoknak rengeteg tápanyagra, köztük kalciumra van szükségük. Mivel azonban az állat táplálék útján nem képes annyi kalciumot felvenni, amennyi elegendő az agancs kifejlődéséhez, ezért a csontszerkezetéből vonja ki a hiányzó mennyiséget, amivel saját csontritkulását okozza. Ilyenkor 25–28 kg-os csontszerkezetéből 4–5 kg anyagot „csoportosít át” fejdísze fejlődésére. Ez bonyolult élettani, hormonális folyamatok eredménye. Egy ekkora bika 12–15 kg-os, de akár súlyosabb agancsot is fejleszthet, majd a fejdísz kialakulása után az állat a csontvázába 2–3 hónap leforgása alatt pótolja az onnan "elvont" kalciumot, így megszűnik az ideiglenesen kialakult csontritkulása.
A csontok – sokan nem is sejtik – „élő szervek”, amelyeknek keménységét, szilárdságát nagy részben a kalciumnak köszönhetik. Az ember csontjaiban hétévenként kicserélődik minden sejt. A mai kor orvostudománya még nem képes a csontritkulás folyamatainak visszafordítására, csak a folyamatot tudja lassítani. Azt kellene kideríteni, hogy az embernél milyen mechanizmus tudná visszaállítani az „elveszett” csontot. Ebben az irányban nagy erővel folynak a további kutatások.
6. Számítsa ki és adja meg, hogy az újonnan képződő agancs tömegének hányad része származik a bika csontvázából! Intervallumok esetén a megadott szélsőértékek számtani átlagával számoljon!
7. Naponta hány gramm kalciumtartalmú anyagot kell beépítenie csontjaiba a szarvasnak a csontritkulás legyőzéséhez? Tegyük fel, hogy minden hónap 30 napos, intervallumok esetén pedig a megadott szélsőértékek számtani átlagával számoljon! Jól követhető módon írja le a számítás menetét is!
Megoldás
6.
4,5kg / 13,5kg = 1/3 = 0,33 1 pont
7.
4,5 kg kalciumsót veszít a csontjaiból; 2,5 hónap (75 nap) alatt pótolja
4500 g kalciumsó / 75 nap = 60 g/nap 1 pont
VII. Paradicsom-genetika
A paradicsomnövény magasságát genetikailag egy gén két allélja határozza meg. Az eredeti paradicsomnövény kb. 30–40 cm magasra nő. Egyik mutáns változata óriásnövésű. Ha a mutáns paradicsomnövényeket az eredeti alacsonyakkal keresztezzük, akkor az első utódnemzedék minden tagja alacsony növésű lesz. A második utódnemzedékben az alacsony növésű növények egyedszáma háromszorosa a magas növésűekének.
A genetikai elemzés során kimutatták, hogy a magasságot meghatározó M jelű gén 860 bázispár (bp) hosszúságú. Az óriásnövést a paradicsom esetében az M gén 383. bázisában bekövetkező pontmutáció (génmutáció) okozza.
A kétféle (magas illetve alacsony növést okozó) allél vizsgálatához a kutatók egy speciális enzimet, az Eco RI-et használják. Ez a bakteriális eredetű enzim a restrikciós endonukleázok csoportjába tartozik. Ezek közös sajátossága, hogy képesek a kétszálú DNS-t megadott bázissorrendű részeinél elhasítani. Az Eco RI hasító helye: 5’-GAATTC-3’. A paradicsomnövényben bekövetkező mutáció miatt ez a hasítóhely „elromlik” és a 383. bázis a nem kódoló „néma” szálban adeninről guaninra cserélődik.
Az Eco RI enzimmel kezelt M gén eredeti formájában két részre esik szét: egy 382 és egy 478 bázispár hosszúságúra. A mutációt elszenvedett génben ilyen változás nem következik be, tehát a vizsgált gén az eredeti 860 bázispár hosszúságú marad. A keresztezési kísérletek alapján háromféle genotípus létezik: homozigóta recesszív (mm), heterozigóta (Mm), homozigóta domináns (MM). A háromféle genotípusú növényből kivont gént Eco RI
Egy mesterséges, de nagy egyedszámú populációban a magas növésű növények a populáció 16%-át teszik ki. Feltételezve, hogy a populáció genetikai egyensúlyban van, adjon választ a következő kérdésekre!
8. Mekkora a recesszív allél gyakorisága? ……………… %
9. Az alacsony növésű növények között hány százalékban fordulnak elő olyanok, amelyekből a magasságért felelős gént kivonva, majd EcoRI-enzimmel kezelve mindhárom méretű DNS-darab előfordul? Válaszát százalékban kifejezve, két tizedesjegy pontossággal adja meg! Eredményét számítással is támassza alá! (2 pont)
Megoldás
8. (A magasnövésűek homozigóta recesszívek, ezért:) q2=0,16 q=0,4 40% 1 pont
9. Az alacsony növésűek 84%-ban vannak jelen. Közöttük van homozigóta domináns és heterozigóta is. Mindhárom DNS-darab a heterozigótákban van jelen, ezek aránya a teljes populációban 2pq = 48%. 1 pont
A keresett arány tehát: 0,48/0,84 = 57,14%. 1 pont
A második pont csak két tizedesjeggyel történő felírással és százalékban kifejezett eredmény esetén adható meg.
VIII. Az idegsejtek nyelve
Az axonon terjedő potenciálváltozás sebességét elsőként német tudós, Hermann von Helmholtz mérte meg oly módon, hogy egy békacomb mozgató idegrostjait ingerelte különböző pontokon, és rögzítette, hogy a válasz mennyi idő múlva észlelhető. Azt figyelte meg, hogy ha az idegrostot az izomhoz közelebbi ponton ingerli, a válasz hamarabb következik be, mintha távolabb ingerelte volna. Feltételezve a potenciálhullám egyenes vonalú egyenletes terjedését, a sebességet ezután már egyszerű volt kiszámolni.
11. Tételezzük föl, hogy az izomtól 70 mm távolságra levő pontot ingerelve a válasz 1,4 ms (0,0014 s) múlva következett be, míg ha az izomtól 20 mm távolságra fekvő pontot ingerelt, akkor az izom már 0,4 ms múlva összerándult. Számolja ki, milyen gyorsan terjedt az ingerület ebben a békaideg-szakaszban!
Megoldás
11. Az ingerületi hullám a 70-20 = 50 mm-es távolságot 1,4-0,4 = 1 ms alatt tette meg, sebessége tehát a v = s/t összefüggés alapján: 50 m/s. 1 pont
IX. B Fenntarthatatlan fejlődés
Energiamérleg
Az energia-arányszám (E) azt mutatja meg, hogy egységnyi tömegű mezőgazdasági termék elfogyasztásából nyerhető és annak előállításához szükséges energia mennyisége hogyan aránylik egymáshoz.
 |
4. Egy 20 éves férfi napi energiaigénye 10 MJ. Tételezzük fel, hogy energiaigényének átlagosan 25%-át teszik ki a gabonából, 25%-át a tejtermékekből, 25%-át a nagyüzemi állattenyésztésből és 25%-át az üvegházi zöldségtermesztésből származó ételek. Számolja ki, legalább hányszor annyi energiát kellett befektetni életmódjának fenntartásához, mint amennyit maga az elfogyasztott táplálék tartalmazott!
Megoldás
 |
Központi biológia emelt szintű írásbeli feladatsorok, javítási-értékelési útmutatók
Feladatok megjelenítése / eltüntetése
- 1. A biológia érettségi gyakorlati feladatai
- 2. Számítási feldatok
- 3. Központi biológia emelt szintű írásbeli feladatsorok, javítási-értékelési útmutatók (Forrás: Oktatási Hivatal)
- 2005. május-június feladatlap
- 2005. május-június javítási-értékelési útmutató
- 2005. október-november feladatlap
- 2005. október-november javítási-értékelési útmutató
- 2006. február-március feladatlap
- 2006. február-március javítási-értékelési útmutató
- 2006. május-június feladatlap
- 2006. május-június javítási-értékelési útmutató
- 2006. május-június feladatlap 2.
- 2006. május-június javítási-értékelési útmutató 2.
- 2006. október-november feladatlap
- 2006. október-november javítási-értékelési útmutató
- 2007. május-június feladatlap
- 2007. május-június javítási-értékelési útmutató
- 2007. május-június feladatlap 2.
- 2007. május-június javítási-értékelési útmutató 2.
- 2007. október-november feladatlap
- 2007. október-november javítási-értékelési útmutató
- 2008. május-június feladatlap
- 2008. május-június javítási-értékelési útmutató
- 2008. május-június feladatlap 2.
- 2008. május-június javítási-értékelési útmutató 2.
- 2008. október-november feladatlap
- 2008. október-november javítási-értékelési útmutató
- 2009. május-június feladatlap
- 2009. május-június javítási-értékelési útmutató
- 2009. május-június feladatlap 2.
- 2009. május-június javítási-értékelési útmutató 2.
- 2009. október-november feladatlap
- 2009. október-november javítási-értékelési útmutató
- 2010. május-június feladatlap
- 2010. május-június javítási-értékelési útmutató
- 2010. május-június feladatlap 2.
- 2010. május-június javítási-értékelési útmutató 2.
- 2010. október-november feladatlap
- 2010. október-november javítási-értékelési útmutató
- 2011. május-június feladatlap
- 2011. május-június javítási-értékelési útmutató
- 2011. május-június feladatlap 2.
- 2011. május-június javítási-értékelési útmutató 2.
- 2011. október-november feladatlap
- 2011. október-november javítási-értékelési útmutató
- 2012. május-június feladatlap
- 2012. május-június javítási-értékelési útmutató
- 2012. május-június feladatlap 2.
- 2012. május-június javítási-értékelési útmutató 2.
- 2012. október-november feladatlap
- 2012. október-november javítási-értékelési útmutató
- 2013. május-június feladatlap
- 2013. május-június javítási-értékelési útmutató
- 2013. május-június feladatlap 2.
- 2013. május-június javítási-értékelési útmutató 2.
- 2013. október-november feladatlap
- 2013. október-november javítási-értékelési útmutató
- 2014. május-június feladatlap
- 2014. május-június javítási-értékelési útmutató
- 2014. május-június feladatlap 2.
- 2014. május-június javítási-értékelési útmutató 2.
- 2014. október-november feladatlap
- 2014. október-november javítási-értékelési útmutató
- 2015. május-június feladatlap
- 2015. május-június javítási-értékelési útmutató
- 2015. május-június feladatlap 2.
- 2015. május-június javítási-értékelési útmutató 2.
- 2015. október-november feladatlap
- 2015. október-november javítási-értékelési útmutató
- 2016. május-június feladatlap
- 2016. május-június javítási-értékelési útmutató
- 2016. május-június feladatlap 2.
- 2016. május-június javítási-értékelési útmutató 2.
- 2016. október-november feladatlap
- 2016. október-november javítási-értékelési útmutató
- 2017. május-június feladatlap
- 2017. május-június javítási-értékelési útmutató
- 2017. május-június feladatlap 2.
- 2017. május-június javítási-értékelési útmutató 2.
- 2017. október-november feladatlap
- 2017. október-november javítási-értékelési útmutató
- 2018. május-június feladatlap
- 2018. május-június javítási-értékelési útmutató
- 2018. május-június feladatlap 2.
- 2018. május-június javítási-értékelési útmutató 2.
- 2018. október-november feladatlap
- 2018. október-november javítási-értékelési útmutató
- 2019. május-június feladatlap
- 2019. május-június javítási-értékelési útmutató
- 2019. május-június feladatlap 2.
- 2019. május-június javítási-értékelési útmutató 2.
- 2019. október-november feladatlap
- 2019. október-november javítási-értékelési útmutató
- 2020. május-június feladatlap
- 2020. május-június javítási-értékelési útmutató
- 2020. május-június feladatlap 2.
- 2020. május-június javítási-értékelési útmutató 2.
- 2020. október-november feladatlap
- 2020. október-november javítási-értékelési útmutató
- 2021. május-június feladatlap
- 2021. május-június javítási-értékelési útmutató
- 2021. május-június feladatlap 2.
- 2021. május-június javítási-értékelési útmutató 2.
- 2021. október-november feladatlap
- 2021. október-november javítási-értékelési útmutató
- 2022. május-június feladatlap
- 2022. május-június javítási-értékelési útmutató
- 2022. május-június feladatlap 2.
- 2022. május-június javítási-értékelési útmutató 2.
- 2022. október-november feladatlap
- 2022. október-november javítási-értékelési útmutató
- 2023. május-június feladatlap
- 2023. május-június javítási-értékelési útmutató
- 2023. május-június feladatlap 2.
- 2023. május-június javítási-értékelési útmutató 2.
- 2023. október-november feladatlap
- 2023. október-november javítási-értékelési útmutató
- 2024. május-június feladatlap
- 2024. május-június javítási-értékelési útmutató
- 2024. május-június feladatlap 2.
- 2024. május-június javítási-értékelési útmutató 2.
- 2024. október-november feladatlap
- 2024. október-november javítási-értékelési útmutató
- 2025. május-június feladatlap
- 2025. május-június javítási-értékelési útmutató
- 2025. május-június feladatlap 2.
- 2025. május-június javítási-értékelési útmutató 2.
- 2025. október-november feladatlap
- 2025. október-november javítási-értékelési útmutató
Alapszintű jegyzetek
Jegyzetek megjelenítése / eltüntetése
- Alapszintű jegyzetek
- 1. A biológiai kutatás céljai és módszerei
- 2. Az élet eredete és szerveződése
- 3. Öröklődés és evolúció, a biotechnológia módszerei és alkalmazása
- 4. Az ember szervezete és egészsége
- A vázrendszer, vázizomrendszer felépítése és működése
- Az emésztőrendszer felépítése és működése
- Az emésztőrendszer rendellenességei
- A légzőrendszer felépítése és működése
- A légzőrendszer egészségtana
- Az emberi vér és az immunrendszer
- A daganatokról általában
- A szív és az érrendszer
- A nyirokrendszer, a keringés egészségtana
- A kiválasztás
- Testkép, testalkat, mozgásképesség
- A hormonális szabályozás
- Az emberi nemek és a szaporodás biológiai alapjai
- Az ember egyedfejlődése
- Az idegszövet, elemi idegjelenségek, szinapszisok felépítése és működése
- Az idegrendszer általános jellemzése, a gerincvelő
- Az agy felépítése és működése
- Testmozgató rendszerek, vegetatív működés
- A szem felépítése és működése
- Hallás és egyensúlyérzés
- A viselkedés biológiai alapjai, a lelki egyensúly és a testi állapot összefüggése
- Egyedszintű öröklődés
Beszámolók
Mit gondolnak a diákok
Kedves Vizkievicz Tanár Úr! Mi tiszteljük Önt, mert:
Tanár Úr kiemelkedő szakértelemmel tartja az óráit mind alap, mind emelt szinten, a fakton átadott ismereteknek hála versenyelőnnyel indultam az egyetemen. Az órák középpontjában a tananyag lényegi összefüggéseinek megértése volt, emellett számos olyan plusz ismeretet kaptunk, ami meghaladta az emelt érettségi követelményeit, viszont hozzájárult a tantárgy iránti érdeklődésünk és szeretetünk növekedéséhez. Mindezt kiváló jegyzetek segítették. Az órák mindig barátságos hangulatban teltek, Tanár úr partnerként kezelt minket, így mindannyian motiváltan vettünk rajtuk részt. Gondolkodásra és kérdezésre ösztönzött minket, bonyolultabb kérdéseinknek lelkiismeretesen utánanézett. A fakultáció rengeteg pozitív hatást gyakorolt a gondolkodásomra és a tanulási folyamataimra, ezekért nagyon hálás vagyok!
"Az iskolának az a feladata, hogy a kérdezést természetes és leküzdhetetlen szokásunkká tegye."(Örkény István) Számomra Vizkievicz Tanár Úr volt az, aki erre igazán rászoktatott, és ezért azóta is nagyon hálás vagyok neki. Sokat bátorított minket, hogy kérdezzünk óra közben, így mindenkinek sokkal személyesebbé és érdekesebbé váltak az órái. Ami külön figyelmesség volt tőle, hogy amikor néha nem tudott válaszolni a részletekben kutató kérdéseinkre akkor mindig utánajárt a következő órára. Válaszaiban és az óra menetében is mindig arra törekedett, hogy mindenki megértse az adott kérdést. Ha kellett több oldalról is körül jártuk a bonyolultabb témákat, de mindig figyelt arra, hogy addig ne menjünk tovább, amíg valami nem világos. Orvosi tanulmányaim során mindig is azt érzem, hogy hatalmas előnyből indultam a többiekhez képest, amiatt a rendszerezett és megértés központú tudás miatt, amit ő átadott nekünk. Hatalmas motiváció volt, hogy őt is ugyanúgy érdekli a téma mint minket. Ami miatt még külön hálás vagyok neki az az embersége és lelkiismeretessége, hiszem, hogy a személyiségünk is sokat formálódott a kezei között.
Vizkievicz tanár úr óráiról csak pozitívat tudok mondani. Amellett, hogy az órák nagyon jó hangulatban teltek, nagyon hatékonyak is voltak. Valahogy olyan érthetővé és érdekessé tudta tenni a legunalmasabb részeit is a tananyagnak, hogy nem is volt annyira nehéz megtanulni. Nem csak a száraz anyagot adta le, hanem mindig valami pluszt is hozzátett, és ettől voltak olyan színesek az órák. Minden kérdésünkre kaptunk választ, tulajdonképpen sose volt buta kérdés, olyan természetességgel válaszolt a legegyszerűbb kérdésre is. Ha kellett többször elmagyarázta a nehezebb részeket, bármikor kérhettük. A jegyzetek, amiket készít, rengeteget jelentenek a felkészülésben. Nagy segítség, hogy nem kell átnyálazni egy könyvet, ráadásul a fontos dolgok mellett még kiegészítés is van szürke betűvel, ráadásul mindig frissül, nem az öt éve kiadott tankönyvekből tanulunk. Szóbelire való felkészítés során mindig tudja, mit kell mondani ahhoz, hogy a lehető legjobban szerepeljünk, óriási tapasztalattal rendelkezik a tanár úr e téren is. Nekem rengeteget segített az, hogy sokat gyakoroltuk ezeket a szituációkat, elképesztő türelemmel kezelve mindezt. A későbbi tanulmányaim alatt vettem észre, hogy olyan biztos alapokat kaptam, amire bátran építkezhettem a későbbiekben, és ha néha elő is kellett vennem a gimis jegyzeteket, ezen az oldalon mindig megtaláltam őket. Gimnazista éveim legjobb és leghatékonyabb órái voltak a biológia órák, ehhez természetesen a tanár úr személyisége és tanítási módszerei is nagyban hozzájárultak.
Vizkievicz Tanár Úrral nem feladat volt a biosz, hanem hobby. Az a tárgy volt ahol csak be kell ülni az órára és hallgatni a mesét. Bízhattam benne, hogy a kérdéseim után nem marad homályos folt az anyagban. Nem csak az egyetemre való felkészülésben segített, de a további egyetemi tanulmányaimban is. Két egyetemi év után is visszakutatok néha pár elfelejtett alapot az érthető, tartalmas, de jól összegzett jegyzeteiből. Nélküle talán nem tartanék ott ahol most tartok. Olyan stabil alapokat kaptam, ami életem végéig segíteni fog. Köszönöm Tanár úr!
Vizkievicz tanár úr négy éven át tanított nekem biológiát emelt szinten. Ezen túl azzal is hozzájárult biológiai irányú érdeklődésem kibontakoztatásához, hogy gimnáziumi éveim alatt számos orvos előadót hívott meg az iskolába. A szigorú, de korrekt számonkérései nagyszerűen felkészítettek az érettségire, a jegyzetei és a ppt előadásai pedig egyedülálló módon adtak ízelőt az egyetemen alkalmazott oktatási módszerekből, így megkönnyítve számomra az egyetemi tanulást. A tanórákon megszerzett tudás ráadásul biztos alapokat nyújtott számomra többek között az anatómia, a genetika, a sejtbiológia, az élettan, a biokémia és a biotechnológia tárgyakhoz. Tanár úr kiváló szakmaiságán túl emberi értékeivel is példát mutatott nekünk. Nyugodt légkörű, jó hangulatú órái maximálisan elősegítik a tanulást, ezért máig hálás vagyok neki az érettségire történő felkészüléshez nyújtott jó tanácsaiért és az elmúlt egyetemi éveim alatt – kiváló felkészítésének köszönhetően – megspórolt tanulási időért. Mindig is érdekelt ugyan a biológia, de Tanár úr volt az, akinek a hatására annyira megszerettem, hogy végül doktori kutatómunkámat is ezen a tudományterületen fogom végezni. Mindenkinek jó szívvel ajánlom. Őszintén hálás vagyok és szívből kívánom a legjobbakat!
Ha az ember egy barátságos, modern, profi és végtelenül hatékony környezetre vágyik, hogy belevesse magát a bioszba, szívből ajánlom a Bioszféra oldalt, és a Tanár úr óráit, melyek maximálisan produktív, nyugodt, ámde baráti hangulatban teltek. A jegyzetek befogadhatóvá, emberközelivé teszik a tananyagot, és bővelkednek érdekességekben, melyek megkönnyítik még az kemény diónak számító témákkal való ismerkedést. A szürke betűs részek pedig tökéletesen helytálltak, mikor az OKTV-re készültem. A témákra bontás, a képek, a rendszerezés és szerkesztése az anyagnak lehetővé teszi, hogy az órán régen átvett anyagokat, vagy akár teljesen ismeretlen témákat vágjon be az ember komolyabb megerőltetés nélkül. Az érettségi előtti para idején mindenki hálás lehet, amikor nem kell egy vastag könyvet átlapoznia, itt derül ki főleg hatékonysága a modern, emberközeli tálalása a biológiának. Visszanézve, az idő távlatából is lenyűgöz, hogy sikerült ilyen könnyen és élvezhetően ennyit tanulni e két év alatt. Köszönöm szépen!
A Tanár Úr azzal, hogy jegyzeteket csinált nekünk, és így az órán annyi volt a feladatunk, hogy figyeljünk, komolyan vett minket, és felnőttként kezelt. Ez megteremtett egy nagyon jó hangulatot az óráin, és szerettünk bejárni. Én megtanultam "tanulni", amit annyiszor tudtam hasznosítani az egyetemen, ebben nagyon segített a biosz óra. Élvezhető, modern, naprakész és vicces volt! Annyira nagyon hálás vagyok ezért a szilárd alapért, amit itt kaptam.
Jó emlékekkel gondolok vissza a biosz faktos órákra! Nekem a faktos órákban az tetszett a legjobban, hogy az egész tananyag úgy volt megszerkesztve, hogy szisztematikusan felkészítsen az érettségire. A jegyzetek könnyen követhetőek voltak, egyértelműen volt a tananyag felosztva témák szerint és a lényeg meg a mellek információ másképpen volt kiemelve a jegyzetekben ezzel egyértelműsítve mi az, ami elengedhetetlen információ és mi az, ami csak érdekességképpen van benne. A jegyzetekről meg annyit, hogy mind két tesóm és meg sok baratom használta őket az emelt színtű vizsga felkészülésére, és nagyon szerettek és egyértelműnek találták őket.
A Tanár Úr órái tulajdonképpen már a kezdetektől nagyon tetszettek nekem, mások voltak, mint a megszokott „tanórajelleg”. Izgalmas volt nézni a saját készítésű ppt-ket, majd otthon szintén a Tanár Úr által készített jegyzetekből tanulni. A Tanár Úr tanításának, érettségire való felkészítésének köszönhető az, hogy az írásbeli, és a szóbeli előtti utolsó pár napban elég volt csak egyszer átolvasni mindent, és miközben ismételtem tudtam, hogy tulajdonképpen mindenre emlékszem az elmúlt két évből. Az, hogy a 12-et úgy meghúztuk, hogy szinte minden héten írtunk, nem csak az érettségire készített fel minket tökéletesen, hanem megtanított minket rendesen, nagy mennyiségben tanulni, ami a jövőre nézve is rendkívül hasznos lehet majd. Sokszor belegondoltam, hogy mennyire jó dolgunk volt a jegyzetekkel is. Számomra sokkal átláthatóbbak, mint bármelyik bioszkönyv, amibe bármikor is belepillantottam, illetve elég jó megoldásnak találtam, hogy bárhol voltam elérhető volt a bioszféráról. Összességében azt találom a legcsodálatosabbnak, hogy hiába kellett a legtöbbet biológiát tanulnom az elmúlt években, mégis nagyon pozitívan gondolok vissza az órák minden pillanatára, és ez a légkörnek köszönhető. Amikor fáradtak voltunk, vagy épp nem annyira voltunk lelkesek a 8. órában, mindig jött egy jófajta kis poén a Tanár Úrtól, vagy egy pár biztató szó, ami engem mindig nagyon feldobott. Sosem volt szükség szigorkodásra, padracsapkodásra (mint más órákon, néha-néha), mert a Tanár Úr máshogy nyerte el rögtön a diákok tiszteletét: higgadtságával, korrektségével, és mértéktelen tudásával.
Az órákon elvárt volt a figyelem, de a Tanár Úr mindig igyekezett a diákokhoz igazítani az óra menetét, figyelte, hogy tudjuk-e követni. Teljes mértékben át tudta adni a tantárgy iránti szeretetét, lelkiismeretesen készültünk, és örömmel mentünk be minden órára. Rendszeresen vegyült bele a humor az órák hangulatába, de mindezt úgy, hogy közben a tárgynál maradtunk, és hatékonyan tudtunk dolgozni. A rendszeres számonkérés is hozzátartozott az órákhoz, de mindig pontosan tudtuk előre, hogy milyen anyagrészekből számíthatunk kérdésekre, a Tanár Úr rendkívül kedvesen feleltetett, a dolgozatidőpontok kitűzésénél figyelembe vette az adott időszakban a többi tantárgyból való számonkérést is, és javítási lehetőséget is biztosított. Minden dolgozatot utólag részletesen átbeszéltünk, így magukból a tesztekből is sokat tanultunk. Az érettségire való felkészítés rendkívül tudatosan folyt. A hatalmas tudáson kívül, amit a Tanár Úr átadott nekünk a két év során, már „tapasztalattal” is rendekeztünk, mivel rengeteg érettségi feladatot oldottunk az órákon, amiket alaposan át is beszéltünk. Így nagyon jól elsajátíthatta mindenki azt a gondolatmenetet, amit ilyen feladatok megoldásánál követni érdemes. A jegyzetek is a követelményrendszer szerint vannak összeállítva. Minden gyakran kérdezett anyagrészre a Tanár Úr külön felhívta a figyelmünket, bármikor kérdezhettünk, ha valami nem volt világos, a sokaknak bonyolult részeket akár többször is átvettük, rendszeres ismétlésekre került sor az érettségit megelőző évben, egyszóval a felkészítés rendkívül átfogó, precíz és alapos volt!”
Biológia szóbeli érettségi tételek kidolgozott B) feladatai
A szóbeli tételeket Vizkievicz András dolgozta ki.
A kidolgozott biológia szóbeli tételek teljes mértékben magukba foglalják mindazokat az aktuális követelményrendszer által elvárt vizsgakövetelményeket - kulcsfogalmakat, gondolkodási műveleteket -, melyek az adott témakörhöz tartoznak. Az oldalon a böngészést nagyban segíti a kulcsszavakra történő keresés ctrl+f használatával.
Tekintve, hogy az oldal minden tanulni vágyó diák számára költségmentesen elérhető, a tételek anyagának bármilyen kereskedelmi célű felhasználásához semmilyen módon nem járulok hozzá. Sikeres felkészülést kívánok!
Tájékoztató az emelt szintű szóbeli vizsgához, az B) feladatok címei (pdf)
A fekete színű dőlt betűs részek középszinten a szóbeli B tételek lehetséges elvégzendő vizsgálatait, a sárga színű dőlt betűs részek emelt szinten a szóbeli B tételek lehetséges problémafeladatait tüntetik fel.
Az emelt szintű követelmények magukba foglalják a középszintű ismeretanyagot is.
1.1. A biológia tudománya |
Ismertesse a tudományos vizsgálatok menetét, műveleteit alkalmazza vizsgálat- és kísérletelemzésekben.
Hozzon döntést a mérések pontosságáról, azok főbb mutatói (tárgyszerűség, érvényesség, megbízhatóság) alapján.
Találja meg egy kísérleti leírásban a kontroll- és kísérleti csoportot, a kísérleti beavatkozást, a függő és független, valamint a rögzített változókat.
A tudományos kutatás szakaszai
1. Témaválasztás.
2. Kutatási kérdés, probléma, hipotézis (előfeltevés) megfogalmazása, amit bizonyítani vagy cáfolni kell.
A hipotézis egy igazolásra váró elképzelés, a feltett kérdésre adott egyfajta válasz, amely megfelel a tapasztalatnak és a megfigyelt tényeket jól magyarázza, de még hiányzik a bizonyítása.
3. Kutatási terv összeállítása (honnan, mit, miért, hogyan? Cél, módszer meghatározása).
4. A kutatás lefolytatása
- Kutatási elővizsgálat végzése (módszerek, eszközök kipróbálása).
- Adatgyűjtés (források feldolgozása, saját kutatómunka adatainak gyűjtése).
- Adatfeldolgozás és -elemzés (adatok rendszerezése, értékelése, összegzése).
- Megfigyelés, leírás, elemzés, hasonló jelenségekkel való összehasonlítás.
- A feltevés kísérletekkel történő bizonyítása. A kísérlet olyan kutatási eljárás, melynek során a kísérletező végez valamilyen tevékenységet, majd megfigyeli annak következményeit. A kísérletnek megismételhetőnek kell lennie és azonos eredményre kell vezetnie ahhoz, hogy az eredményt tudományos eredménynek nevezhessük.
- A kísérletek kapcsán meg kell határoznunk a kísérleti változók fogalmát:
- Független változó: az a tényező, melynek hatását vizsgáljuk (az eredményt grafikonon ábrázolva értékeit az x tengelyen tüntetjük fel).
- Függő változó: azon tényező, melyet a független változó módosít, melynek a változását mérjük (értékeit az y tengelyről olvashatjuk le).
- A rögzített változókat (egyéb körülményeket, melyek befolyásolhatják a kísérlet eredményeit) állandó értéken tartjuk, hogy egyértelmű legyen az összefüggés a független, ill. a függő változó között.
- Következtetések levonása.
5. Az eredmények nyilvánossá tétele (cikk készítése, véleménycsere kutatók között).
Kísérletezéskor a kutató szabályozza a feltételeket, tervszerűen avatkozik be a jelenségbe. A megfigyeléstől a kísérlet abban különbözik, hogy a vizsgálat során a körülményeket, változókat mi határozzuk meg. A kísérletek megtervezésének, végrehajtásának fontos feltételei;
- a változók pontos mérhetősége,
- a megismételhetőség,
- kontrollvizsgálat, ellenőrzés, melynek során a jelenséget beavatkozás nélkül figyeljük meg, majd vizsgálati eredményeinket ezzel a tapasztalattal vetjük össze.
Példa
1. Témaválasztás: a csírázás hőmérsékletfüggése.
2. Kutatási kérdés: hogyan befolyásolja a környezet hőmérséklete a növényi magok csírázásának ütemét?
3. Kutatási terv összeállítása: magokat különböző hőmérsékletű – más tényezőket tekintve teljesen azonos – környezetben csíráztatva, naponta meghatározzuk a csíranövények növekedésének az ütemét.
4. A kutatás lefolytatása
- Kutatási elővizsgálat végzése.
- Adatgyűjtés.
- Adatfeldolgozás és –elemzés.
- Megfigyelés, leírás, elemzés, hasonló jelenségekkel való összehasonlítás.
- A feltevés kísérletekkel történő bizonyítása.
- Független változó: hőmérséklet (x tengely).
- Függő változó: a magok csírázásának mértéke (y tengely).
- Rögzített változók, pl. a fényerősség, az öntözővíz mennyisége stb.
- Következtetések levonása.
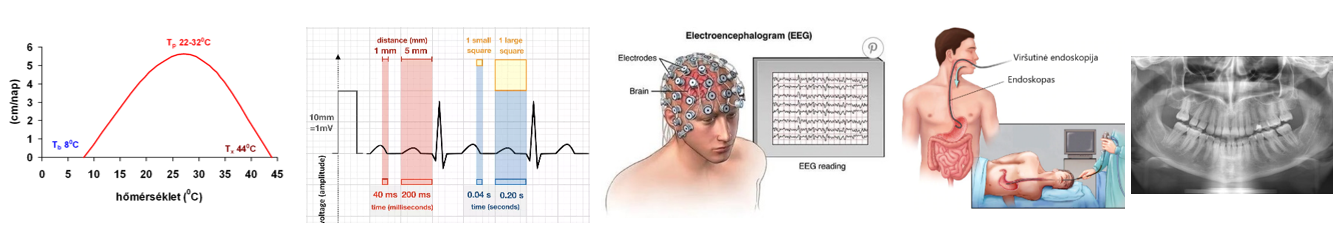 |
Értelmezzen egyszerű, (molekuláris) szűréssel kapcsolatos vizsgálatot.
Szűrés
A szűrés egy fizikai elválasztási eljárás, melynek során
- szilárd anyagokat különítünk el folyadéktól, ill.
- különböző szemcseméretű részecskéket választunk szét egymástól.
A szűrő egy olyan porózus réteg – pl. szűrőpapír -, ami bizonyos szemcseméretű anyagokat átenged, másokat visszatart.
Vázolja fel az emberi EKG, EEG, CT, MRI, UH, röntgen, endoszkópos vizsgálatok alapvető céljait, értelmezzen ezekkel kapcsolatos betegtájékoztatást.
Diagnosztikai vizsgálatok
Az orvosi eljárás első szakasza, amely
- a beteg panasza okának feltárására, állapotának tisztázására,
- valamely betegség fennállásának igazolására vagy kizárására irányul.
A diagnosztikai vizsgálatok a diagnózis meghatározását teszik lehetővé, mely megállapítás az adott betegség fennállását valószínűsíti.
A diagnosztikai vizsgálatok közé tartoznak
- fizikális vizsgálat,
- képalkotó diagnosztika (pl. röntgen, CT, ultrahang, MR),
- labordiagnosztika,
- patológia (szövettani vizsgálat),
- egyéb eszközös vizsgálat, mint pl. EKG, EEG, endoszkóp.
EKG, EEG
Az emberi test folyadékterei, mivel ionokat tartalmaznak, jól vezetik az elektromosságot, ezért a szív működésekor (EKG), ill. az agyi működés (EEG) során keletkezett elektromos jeleket mérni lehet a test felszínén is. Amennyiben a feszültségváltozásokat az idő függvényében ábrázoljuk, jellegzetes görbéket kapunk.
A különféle képalkotó diagnosztikai eszközökkel feltárás nélkül vizsgálható az emberi test.
Endoszkópos vizsgálat
Segítségével a páciens belső szerveiről, testüregeiről kapható betekintés a testnyílásokon keresztül bevezetett kamera segítségével.
Ultrahangvizsgálat
Elsősorban lágyrészek vizsgálatára használatos. Fájdalom, bódítás nélkül lehetséges a belső szerveket működés közben megfigyelni. A vizsgálófej ultrahangot bocsájt ki, mely a belső szervekben részben elnyelődik, részben visszaverődik. Az egyes szövetek, ill. a különféle szervek eltérő felületei más-más módon verik vissza a kibocsájtott ultrahangot, melyet a műszer érzékelve képpé alakít.
Röntgenvizsgálat
Alapvető vizsgálati módszer a mellkasi – tüdő - betegségek, csonttörések, ortopédiai, fogászati problémák, egyes hasi panaszok diagnosztikájában. A röntgenkészülék nagy energiájú elektromágneses sugarakat – ún. röntgensugarakat - bocsájt ki. A sugárzás bejutva a testbe, a különféle szövetekben eltérő mértékben nyelődik el, vagy másképpen az egyes szövetek különböző mértékben engedik át a sugarakat. Ezért a test mögött elhelyezett röntgenfilmen sötétebb és világosabb területek láthatók. A nagyobb sűrűségű részek, pl. csontok, jobban elnyelik a sugarakat, ezért a fotólemezen világos sávként tűnnek fel, míg a lágyrészek, melyek inkább átengedik a sugárzást, sötét foltokként jelennek meg.
CT
A computer tomográfia több síkban röntgen-rétegfelvételeket készít a vizsgált területről, amiből számítógépes eljárással akár háromdimenziós képek is létrehozhatók.
MR (mágneses rezonancia)
Az MR-készülék által generált rendkívül erős mágneses térben egyes atomok, pl. a hidrogénatomok, elektromágneses rádióhullámokkal történő gerjesztés után alapállapotba visszakerülve szintén elektromágneses hullámokat bocsátanak ki, képpontokat képezve, melyeket egy nagy teljesítményű számítógép részletes felbontású, háromdimenziós képpé alakít.
Az eszköz alkalmas rándulás, ficam, törés, gyulladások, agyvérzés, sclerosis multiplex, epilepszia, gerincferdülés, gerincsérv, csontritkulás, érbetegségek és fejlődési rendellenességek diagnosztikájára.
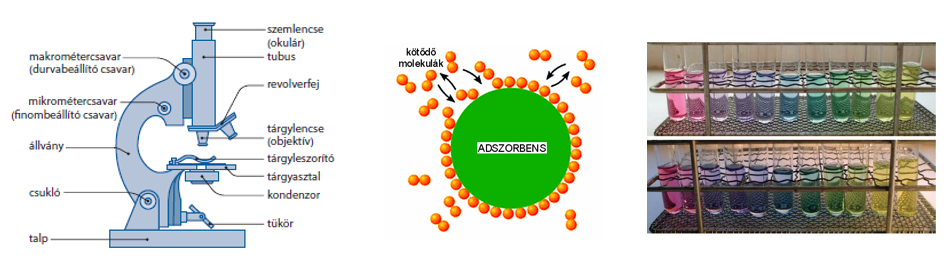 |
Ismertesse a fénymikroszkóp használatának alapelveit, tudja, hogyan kell kiszámítani a mikroszkóp nagyítását. Készítsen vázlatrajzot a megfigyelt preparátumról, mikroszkópos metszetről.
A fénymikroszkóp
Összetett optikai rendszer. Főbb részei:
- talpazat,
- a tubus,
- a revolverfoglalat,
- mozgatható tárgyasztal,
- kezelőgombok,
- megvilágító rendszer.
A tubus a szemlencse (okulár) és a tárgylencse (objektív) befoglalására szolgál. A tárgyasztal az objektív alatt található, amely a preparátum elhelyezését teszi lehetővé.
Az optikai rendszer nagyítása az objektív és az okulár nagyításának szorzatával egyenlő. A fénymikroszkóp nagyítása maximum 1500-szoros, elméleti felbontóképességük 0,2 mikrométer.
Nagyítás alatt egy kép méretének megnövelését értjük, jele N. A nagyítás a képméret (K) és a tárgyméret (T) hányadosa. N=K/T
A felbontóképesség az a legkisebb távolság, amelynél az elhelyezkedő tárgypontok még különálló képpontokként képződnek le. Az emberi szem felbontóképessége 0,2 mm.
Videó
https://drive.google.com/file/d/11jK-_VVH9STjICBRKw71Pzi6VmQtBb5V/view?usp=sharing
Mutassa ki az orvosi szén nagy felületi megkötő képességét festékoldattal. Tudja magyarázni és értelmezni a kísérletet, értse annak következményeit, alkalmazási lehetőségeit.
Orvosi szén felületi megkötőképességének vizsgálata
Vizsgálat: Öntsön lombikba kb. 200 cm3 vörösbort vagy vizet, amit 5-6 csepp fukszinoldattal festett meg. 4 szem orvosi szenet törjön össze a dörzsmozsárban! Tegye a szenet a lombikba, és jól rázza össze! Szűrje le az oldatot, és figyelje meg a színét!
Tapasztalat és magyarázat: A kiinduláskor vörös oldat az aktív szénnel történt összekeverést követően elveszítette a színét, ezért a szűrés során színtelen oldat csepeg le a tölcsérből. A jelenség oka az adszorpció, amely felületen történő anyagmegkötést jelent. Az aktív szénpor összfelülete rendkívül nagy, ezért képes a festékanyag maradéktalan megkötésére.
https://www.youtube.com/watch?v=LtnWG7eqMzo
Felülethez kötődés (adszorpció)
Gázok, ill. folyadékok részecskéinek szilárd felületen történő megkötődését adszorpciónak nevezzük. A megkötődés többnyire gyenge másodlagos kötésekkel történik. Biológiai jelentősége pl. az enzimek működésében, ill. a talajkolloidok szerepében van (lásd enzimek, ill. talaj). Az adszorpció ellentéte a deszorpció, melynek során az adszorbens felületéről távozik a megkötött anyag.
Az adszorpció mértékét befolyásolja a hőmérséklet, a megkötendő gázalkotó parciális nyomása és az adszorbens felületének nagysága.
Fajlagos felület
A fajlagos felület egységnyi tömegű anyag felülete, az anyag felületének és tömegének a hányadosa. Minél apróbb szemcséjű egy anyag annál nagyobb a fajlagos felülete. A nagy fajlagos felülettel rendelkező anyagok - aktívszén, kolloidok - kiváló adszorbensek.
Végezzen kísérletet az antociánok pH-tól függő színváltozásának vizsgálatára, ismertesse a tapasztalatokat.
Az antociánok színe pH függő. Savakban vörösek, lúgokban kékek.
https://www.youtube.com/watch?v=ZqNDGLUUBlw
https://www.youtube.com/watch?v=EcHIPyh1W60
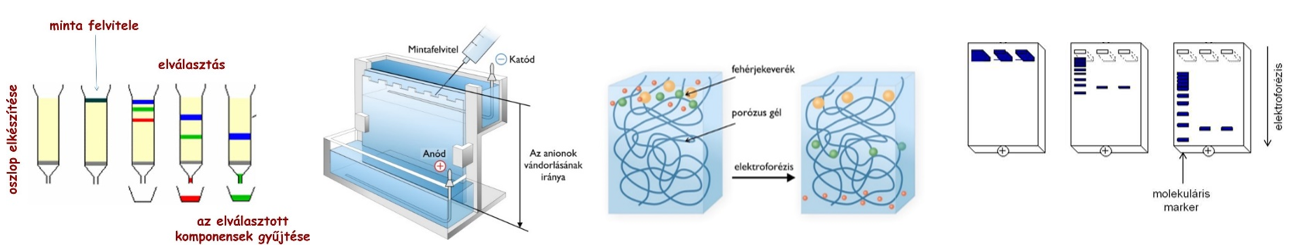 |
Értelmezze különböző elválasztástechnikai eljárások (kromatográfia, centrifugálás, gélelektroforézis) eredményét biológiai vizsgálatokban.
Kromatográfia
Gáz- vagy folyadékelegyek szétválasztására szolgáló vegyi eljárás. Az eljárás során az elválasztandó alkotórészek két fázis között oszlanak meg;
- az egyik fázis álló,
- a másik mozgó fázis (minta), tulajdonképpen a szétválasztandó keverék, amely átáramlik az álló fázison.
Az álló fázis többnyire szilárd anyag.
A mozgó fázis lehet
- folyadék vagy
- gáz, ami egyben folyadék- vagy gázkromatográfiát jelöl.
Az álló fázisként szolgáló finomszemcsés adszorbens - mely fontos sajátsága a nagy fajlagos felület -, lehet például Al2O3, aktív szén, szűrőpapír (cellulóz). Ez alkotja a kromatográfiás oszlopot. Erre viszik fel a szétválasztandó elegyet, ami mint mozgó fázis mozog az álló fázison keresztül.
Az álló fázis felületén a mozgó fázis komponensei különböző erősséggel megkötődnek, adszorbeálódnak. Az egyes alkatrészek aszerint választódnak el, hogy mennyire erősen kötődnek az álló fázisra. Azok a komponensek, amelyek erősebben adszorbeálódnak, kisebb sebességgel vándorolnak, mint azok, amelyek kevésbé adszorbeálódnak, ennek megfelelően a folyadékmozgás irányában az alkotórészek elválasztódnak egymástól, és gyűrű formájú mozgó zónákba összpontosulnak.
https://drive.google.com/file/d/0B15KJwYa2uMFbFkzXzU1Uk84Ylk/view?usp=sharing
Gélelektroforézis
A gélelektroforézist leggyakrabban nukleinsavak és fehérjék méret szerinti elválasztására használják. Az eljárás segítségével a makromolekulákat három fizikai tulajdonságuk, a méretük, alakjuk és a töltésük alapján tudjuk elválasztani.
A gélelektroforézis alapelve, hogy a töltéssel rendelkező molekulák elektromos térben, össztöltésüknek megfelelően, az ellentétes töltésű elektróda felé vándorolnak. A vándorlás sebessége többek között függ a molekula töltésétől, tömegétől és alakjától.
A gél egy térhálós szerkezetű anyag, mely a molekulák méretétől, alakjától függően lassítja azok mozgását. Ha a molekula mérete kicsi a pórusokhoz képest, gyorsan mozog, ha nagy, a molekula szinte mozdulatlan marad a gélben.
Méret szerinti elválasztás esetén az ismeretlen mintáink mellett mindig kell futtatnunk egy ismert tömegű/hosszúságú fehérjéket/nukleinsavakat tartalmazó ún. molekulamarkert.
A megfelelő ideig történt futtatás után a feszültséget kikapcsoljuk és a gélt megfestjük, pl. a nukleinsavak esetében valamilyen, a nukleinsavakhoz kötődni képes fluoreszcens festék oldatával, majd az elválasztott nukleinsavakat a fluoreszcens festéket gerjeszteni képes fénnyel (pl. UV-fénnyel) láthatóvá tehetjük.
https://www.youtube.com/watch?v=czpuYnrU4E8
Centrifugálás
Elegyek komponenseit szétválaszthatjuk centrifugálással is, melynek során a keverék alkotórészei a centrifugális erő hatására tömegük, ill. sűrűségük szerint elkülönülnek.
A centrifuga egy forgó rotort tartalmaz. A rotor fogadja be a mintákat tartalmazó centrifugacsöveket. A centrifugális erő - melyet a tengely körül forgó rendszerrel lehet előállítani – hatására a nagyobb tömegű, sűrűségű anyagok a centrifugacsövek alján, míg a kisebb tömegű részecskék a csövek felső részén rétegződnek.
Magyarázza a fény- és az elektronmikroszkóp felbontóképességét, a kapható információk különbségét.
Nagyítás alatt egy kép méretének megnövelését értjük, jele N. A nagyítás a képméret (K) és a tárgyméret (T) hányadosa. N=K/T
A felbontóképesség az a legkisebb távolság, amelynél az elhelyezkedő tárgypontok még különálló képpontokként képződnek le. Az emberi szem felbontóképessége 0,2 mm.
A fénymikroszkóp nagyítása maximum 1500-szoros, elméleti felbontóképességük 0,2 mikrométer, ezért fénymikroszkópban a baktériumok még láthatók, de a vírusok már nem.
A fénymikroszkópok látható fényt (400-800 nm), az elektronmikroszkópok ugyanakkor jóval kisebb hullámhosszú, mágnesekkel fókuszált elektronsugarat használnak képalkotásra.
Az elektronmikroszkóppal 100 000-szeres nagyságrendű nagyítás és 0,5 nm-es felbontóképesség érhető el, amivel már nem csak a sejtek, hanem a sejtalkotók finomszerkezete is tanulmányozható, továbbá megfigyelhetők a vírusok is.
 |
Értelmezzen és tervezzen ezüsttükör-próbán, Lugol-próbán és biuret-reakción alapuló kísérletet. Magyarázza a megfigyelhető tapasztalatokat a bekövetkező kémiai változások alapján.
A glükóz kimutatása etüsttükör-próbával
A szénhidrátokat a reakcióképes aldehid-csoporton keresztül lehet kimutatni. A reakció lényege, hogy az aldehidcsoport megfelelő körülmények között karboxil-csoporttá oxidálódik, miközben a reagens anyagok – pl. ezüstionok - színváltozás kíséretében redukálódnak.
A próba során tehát az ezüstionokat lúgos közegben az aldehidcsoport fém ezüstté redukálja, amely kiválva az oldatból bevonatot képez az edény falán.
- Kémcsőbe öntsünk 4 cm3 1 tömegszázalékos ezüst-nitrát-oldatot,
- majd adjunk hozzá annyi ammóniaoldatot, hogy a képződő csapadék éppen feloldódjék!
- Ezután öntsünk a kémcsőbe 2 cm3 10 tömegszázalékos szőlőcukor-oldatot,
- rázzuk össze, majd tegyük a kémcsövet 80 Celsius-fokos vízfürdőbe.
https://drive.google.com/file/d/0B1BZOTldajAjbWxjUGttVGhnNnc/view?usp=sharing
A keményítő kimutatása lugol-próbával
- Por keményítőből melegítéssel készítsünk keményítőoldatot.
- Híg oldatba cseppentsünk 1-2 csepp kálium-jodidos jódoldatot – Lugol-oldatot.
- + próba esetén az oldat megkékül.
- Melegítés hatására elszíntelenedik, visszahűtéskor újra kék lesz.
https://drive.google.com/file/d/1TZoSfkICnmJyjfYMpoduMgbMlHSJGS0R/view?usp=sharing
Magyarázat
Az apoláris jódmolekulák a poláris vízben barna színűek. A jódmolekulák mérete olyan, hogy pont beleférnek az amilóz spiráljába, onnan kiszorítva a vízmolekulákat, apoláris közeget hozva létre. Az apoláris közegben a jód színe kékre változik.
Melegítés hatására a hőmozgás miatt a jódmolekulák kibújnak a spirálból a kék szín elhalványodása kíséretében. Hűtéskor a folyamat ellenkező irányú.
A fehérjék kimutatása biuret-próbával
A biuret-próbával a fehérjékben jelenlévő amidcsoportot lehet kimutatni. Pozitív próba esetén ibolyaszíneződést tapasztalunk, mivel a reagensben található rézionok komplexet képeznek az amidcsoporttal.
Egy kevés fehérjeoldathoz adjunk néhány csepp 40%-os nátrium-hidroxid-oldatot, majd 1-2 csepp réz(II)-szulfát reagenst.
https://drive.google.com/file/d/1FeMFJxsNbFNSiFUHMECStgkHkSBipG8G/view?usp=sharing
Állapítsa meg egydimenziós, idő- és helyfüggő változásokban a változás gyorsaságát, értelmezze a változást bemutató grafikonok meredekségét.
Egydimenziós változás: egyetlen irány mentén (pl. idő vagy hely szerint történő változás).
A változás gyorsasága azt mutatja meg, hogy egy adott mennyiség milyen gyorsan változik egy másik mennyiség függvényében, azaz milyen gyorsan megy végbe egy biológiai folyamat. Például:
- Sejtosztódás: hány sejttel nő a sejttömeg időegységenként?
- Enzimaktivitás: mennyi termék keletkezik időegység alatt?
A meredekség azt mutatja meg, hogy egy egységnyi változás a vízszintes tengelyen (pl. idő vagy hely) mekkora változást eredményez a függőleges tengelyen (pl. hőmérséklet, sejtszám stb.), másképp egy egységnyi idő vagy hely alatt mennyit változik a vizsgált biológiai mennyiség.
A populáció egyedszámának változása az idő függvényében tipikusan egy olyan biológiai jelenség, ahol egydimenziós időfüggő változás történik. Ennek a változás gyorsasága a növekedési ráta, amit a grafikon meredeksége mutat meg. A grafikont lásd alább.
1.2. Fizikai, kémiai alapismeretek |
 |
Vizsgálja és magyarázza növényi bőrszövet-nyúzatban lezajló plazmolízis jelenséget.
Növényi szövetek vizsgálata, metszetkészítés
Bőrszöveti metszetek:
- hagymalevél epidermisz nyúzata,
- tulipánlevél epidermisz nyúzata.
Nyúzat készítése
Ilyenkor az epidermiszt csipesszel letépjük az alatta levő szövetekről, ami azért lehetséges, mert az epidermiszsejtek egymáshoz jóval erősebben kapcsolódnak, mint az alatta elhelyezkedő sejtekhez.
Ozmózis az a jelenség, amikor az oldószer – a víz – féligáteresztő hártyán keresztül a hígabb oldatrész felől a töményebb oldatrész felé áramlik diffúzióval.
Az ozmózis törvényei szerint a növényi sejt citoplazmája és a környezete közötti koncentrációkülönbség határozza meg a víz mozgásának irányát.
- Ha a külső közeg hígabb (hipotóniás), a víz a sejtbe áramlik, a citoplazma – a vakuólum - vízzel telítődik, térfogata megnő, a sejt megduzzad. A citoplazma nekifeszül a sejtfalnak, ami ellenáll a sejt térfogatnövekedésének. Ekkor citoplazma sejtfalra gyakorolt nyomását turgornyomásnak nevezünk.
- Ha a külső közeg töményebb (hipertóniás), mint a sejt vizet veszít, a citoplazma – vakuólum - összezsugorodik, plazmolitikus állapot jön létre, a jelenség a plazmolízis.
Plazmolízis vizsgálata
- Készítsünk két vizsgálható nyúzatot a lilahagyma húsos alleveléből!
- Az egyik nyúzatot helyezzük a tárgylemezre előzőleg cseppentett KCl-oldatba, a másikat egy másik tárgylemezre, amelyre CaCl2-oldatot cseppentett! Az inkubációt 5-7 percig végezzük!
- Helyezzünk fedőlemezt a mintáinkra, és vizsgáljuk meg őket fénymikroszkóppal!
- A sejt zsugorodását az antocián tartalmú vakuólum színe miatt lehet jól nyomon követni.
https://www.youtube.com/watch?v=eWcheUXSqMA
Kapcsolja össze a növények hervadását és az ozmózis jelenségét.
A növények hervadása és az ozmózis jelensége szorosan összefüggnek egymással, a hervadás közvetlen oka az, hogy ozmózissal víz távozik a sejtekből, így azok elveszítik turgornyomásukat.
Normál esetben a talajban elegendő víz van. A víz ozmózissal beáramlik a sejtekbe. A sejtek megduzzadnak, a turgornyomás nő. A növény feszes.
Szárazság vagy sófelhalmozódás esetén a talaj oldatának koncentrációja magasabb, mint a sejtplazmáé. A víz ozmózissal kiáramlik a sejtekből. A sejtek elveszítik víztartalmukat, turgoruk csökken. A sejtek összeesnek, a növény elhervad.
Tervezzen és értelmezzen a diffúzióval és az ozmózissal kapcsolatos kísérleteket.
Diffúzió
Koncentrációkülönbség hatására a nagyobb koncentrációjú hely felől a kisebb felé irányuló, spontán végbemenő anyagtranszportot diffúziónak nevezzük. A diffúzió eredményeként a rendszerben a diffundáló anyag koncentrációja kiegyenlítődik, a részecskék a rendelkezésre álló teret egyenletesen kitöltik. Elsődlegesen gázokban és oldatokban van jelentősége.
A diffúziót a részecskék rendezetlen hőmozgása – ún. Brown-féle mozgás – teszi lehetővé. A diffúzió sebessége a diffundáló molekula méretével és a közeg sűrűségével fordítottan, a hőmérséklettel pedig egyenesen arányos.
A diffúziónak óriási a jelentősége az élővilágban, az anyagáramlás alapvető módja, a passzív transzportfolyamatok alapja. Diffúzióval
- lép be pl. az oxigén a tüdő légteréből a hajszálerekbe, a hajszálerekből a szövetekbe, a diffúz légzésű állatoknál a kültakarón át a testbe,
- az akciós potenciálok lezajlásának hátterében szintén a különféle ionok – pl. Na+, K+ – passzív transzportfolyamatai állnak.
- Továbbá diffúzióval – pontosabban ozmózissal, lásd alább – mozog a víz a különböző határfelületeken keresztül, pl. ilyen a növények vízfelvétele a talajból, a vesében a víz visszaszívódása, a bélcsőben a víz felszívódása stb.
https://www.youtube.com/watch?v=sA222HoL760
Ozmózis
Az ozmózis folyamatában az oldószer – a víz – féligáteresztő hártyán – ún. szemipermeábilis hártyán – keresztül diffundál a hígabb oldatrész felől a töményebb oldatrész felé. A féligáteresztő hártyák olyan pórusos membránok, amelyek az oldott anyag nagyobb molekuláit nem, csak a kisebb méretű oldószer molekuláit engedik át. Amennyiben az ilyen hártyák két oldalán különböző koncentrációjú oldatok találhatók, akkor az oldószer részecskéi a hígabb oldat felől – ahol a víz koncentrációja nagy – a töményebb oldat felé – ahol a víz koncentrációja kisebb, hiszen az oldatban oldott anyag is van – diffundálnak.
Kísérlet
Egy üvegedénybe desztillált vizet öntünk, majd ebbe belehelyezünk egy üvegcső végére kötözött, cukoroldatot – szacharóz – tartalmazó féligáteresztő tulajdonságú celofánzsákot.
A kísérlet kezdetén az üvegkádban levő víz szintje és az üvegcsőben található cukoroldat vízszintje megegyezik. Néhány óra múlva azt tapasztaljuk, hogy az üvegcsőben levő oldat szintje megemelkedik.
A jelenség hátterében az ozmózis áll.
- A rendszerben koncentrációkülönbség van mind a cukorra, mind a vízre nézve.
- A cukor diffúzióját, nagymolekula mérete miatt, a féligáteresztő hártya megakadályozza.
- A pohárban tiszta víz van, ezért annak térfogategységeiben több vízmolekula van, mint a zsákban lévő cukoroldat térfogategységeiben. Emiatt a diffúzió a vízmolekulákat a zsák belseje felé hajtja.
- Kifelé is lépnek vízmolekulák, de kisebb sebességgel, mint befelé.
- Ha a vízbelépés sebessége v1, a kilépésé pedig v2, akkor a folyamat elején v1 > v2, ezért a zsák folyadékszintje emelkedni fog, de csak addig, ameddig a folyadékoszlop hidrosztatikai nyomása egyenlővé nem válik az ún. ozmózisnyomással.
- Ennek az az oka, hogy az emelkedő folyadékoszlop nyomása fokozza a víz kilépési sebességét, s így v2 végül addig nő, amíg v1-gyel egyenlő lesz.
- Amikor v1=v2, dinamikus egyensúlyi állapot áll be.
https://www.youtube.com/watch?v=370t8MSoRKg
https://www.youtube.com/watch?v=29_Q0Ll5fI0
https://www.youtube.com/watch?v=tHzkRtzVmUM
Ozmózisnyomásnak nevezzük azt a nyomást, amelyet a tiszta oldószerrel féligáteresztő hártyán át kapcsolatban lévő oldatra kell kifejteni ahhoz, hogy a dinamikus egyensúly beálljon.
Az élő szervezetben különféle határfelületek rendelkeznek féligáteresztő sajátságokkal, mint pl. a sejtmembránok vagy a kapillárisok fala. E felületek két oldalán a különféle méretű molekulák koncentrációkülönbségét az okozza, hogy az ozmotikusan aktív szerves vegyületek a kolloid mérettartományba sorolhatók, s emiatt ezekre nézve e határfelületek féligáteresztő tulajdonságúak. A kolloid részecskék mérete 1-1000 nm-ig (IUPAC) terjed.
Élettani példák az ozmózisra
A növények életműködéseiben fontos szerepe van az ozmózisnak, a jelenség szerepet játszik
- a talajból történő vízfelvételben,
- a gázcserenyílások nyitásában-záródásában,
- a szerves anyagok áramlásában a háncs rostaelemeiben.
Az állati szervezetekben, és így az emberi szervezetben zajló vízmozgás irányának meghatározásában is óriási szerepe van az ozmotikusan aktív anyagoknak.
A vese nefronjaiban a szűrletből jelentős vízvisszaszívás történik, ami annak köszönhető, hogy az elvezetőcsatornák, ill. a gyűjtőcsövek olyan szöveti környezetben futnak, amelynek az ozmotikus nyomása jelentősen meghaladja a szűrlet ozmotikus nyomását.
A tápcsatornában is a víz passzív transzporttal szívódik fel,annak köszönhetően, hogy a béltartalom koncentrációja többnyire kisebb a bélhámsejtek citoplazmájának koncentrációjánál.
Tervezze meg és magyarázza az enzimműködéshez szükséges optimális kémhatást és hőmérsékletet bemutató kísérletet, értékelje annak eredményeit.
A biokémiai reakciók hőmérséklet-, ill. kémhatás-érzékenysége is az aktív centrum bonyolult szerkezetével függ össze, mivel már a hőmérséklet, ill. a kémhatás kismértékű megváltozásának hatására a kötőzseb elveszti a szubsztrátéval komplementer szerkezetét. Az enzimek optimális működéséhez tehát meghatározott környezeti feltételek – hőmérséklet, kémhatás, ionkoncentráció – szükségesek.
Egy enzimműködéshez szükséges optimális kémhatás, ill. hőmérséklet bemutatására tervezett kísérlet segít megérteni, milyen pH-, ill. hőmérséklettartományban működik a legjobban egy enzim, és hol csökken a hatékonysága.
Kísérlet célja megállapítani, hogy egy adott enzim (pl. amiláz) mely pH-értéken, ill. hőmérsékleten működik a leghatékonyabban.
A pH-függés vizsgálata
Szükséges eszközök és anyagok:
- Keményítőoldat (mint szubsztrát)
- Amilázoldat (mint enzim)
- Különböző pH-jú pufferoldatok (pl. pH 3, 5, 7, 9, 11)
- Jódoldat (keményítő kimutatására)
- Kémcsövek, pipetták
- Vízfürdő (37 °C, a testhőmérséklet biztosítására)
- Időmérő (stopper)
Külön kémcsövekbe keverjünk össze:
- egyenlő mennyiségű keményítőoldatot
- amiláz enzimet
- adott pH-jú pufferoldatot
A kísérlet menete
- Tegyük a kémcsöveket vízfürdőbe (37 °C-on).
- Indítsuk az időmérést.
- 30 másodpercenként cseppentsünk egy kis mennyiséget fehér csempén jódoldatra.
- Amíg kékes szín jelenik meg, van még keményítő, az enzim még dolgozik.
- Ha nincs elszíneződés, a keményítő lebomlott, a reakció befejeződött.
- Mérjük meg, mennyi idő alatt bomlott le teljesen a keményítő minden egyes pH-nál.
Eredmények kiértékelése:
Készítsünk grafikon:
- X tengely: pH-érték
- Y tengely: enzimaktivitás (az enzimek működésének hatékonyságát az enzimaktivitással jellemezzük. Az enzimaktivitás megmutatja, hogy adott idő alatt, adott mennyiségű enzim, adott feltételek mellett mennyi szubsztrátot képes átalakítani)
Az enzimaktivitás változása egy haranggörbével jellemezhető.
- Lassú reakció szélsőséges pH-knál (pl. pH 3 vagy 11)
- Leggyorsabb reakció az optimális pH-nál (pl. pH 7)
Értelmezés:
- Az enzimek térszerkezete pH-függő.
- Az optimális pH-n a fehérje térszerkezete ideális az aktív centrum működéséhez.
- Túl savas vagy túl lúgos közegben az enzim denaturálódik, és elveszíti az aktivitását.
https://www.youtube.com/watch?v=yFGt0ct2mrU
A hőmérsékletfüggés vizsgálata
Kísérlet célja megállapítani az amiláz enzim optimális működési hőmérsékletét, azaz azt a hőmérsékletet, ahol a keményítő lebontása a leggyorsabban történik.
Szükséges eszközök és anyagok
- Amilázoldat (enzim)
- Keményítőoldat (szubsztrát)
- Jódoldat (a keményítő kimutatására)
- Kémcsövek, pipetták
- Vízfürdők különböző hőmérsékleteken (pl. 0 °C, 20 °C, 37 °C, 50 °C, 70 °C)
- Időmérő (stopper)
- Fehér csempe vagy tálca
Minden hőmérséklethez külön kémcsőbe öntsünk:
- azonos mennyiségű keményítőoldatot
- azonos mennyiségű amilázoldatot
A kísérlet menete
- Állítsunk be 5 különböző hőmérsékletű vízfürdőt (pl. 0 °C, 20 °C, 37 °C, 50 °C, 70 °C)
- Helyezzük a kémcsöveket ezekbe, és hagyjuk néhány percig akklimatizálódni.
- Minden kémcsőben keverjük össze az amilázt és a keményítőt, majd indítsuk az időmérést.
- Minden 30 másodpercben vegyünk ki mintát, és cseppentsünk jóddal kezelt fehér csempére.
- Amikor nincs elszíneződés (sárga marad), a keményítő lebomlott.
- Jegyezzük fel, mennyi idő alatt bontotta le az enzim a keményítőt minden hőmérsékleten.
Eredmények értékelése
Készítsünk grafikon:
- X tengely: hőmérséklet (°C)
- Y tengely: enzimaktivitás
Az enzimaktivitás változása egy haranggörbével jellemezhető.
Értelmezés
- Az enzimek működéséhez van egy optimális hőmérséklet, amelyen a legaktívabbak (pl. emberi amiláz esetén ~37 °C).
- Túl alacsony hőmérsékleten az enzim lassú, nem hatékony.
- Túl magas hőmérsékleten az enzim denaturálódik, így elveszíti működőképességét.
https://www.youtube.com/watch?v=jOBUDzI5XTk&t=71s
https://www.youtube.com/watch?v=eAUOV4vzhbs
2.1. Szervetlen és szerves alkotóelemek |
Végezzen el vizsgálatot a C, N, Ca2+ kimutatására.
Értelmezzen biogén elemek (a szén, a hidrogén, a nitrogén, a kén és a vas) kimutatására irányuló kísérletet.
A biogén elemek kimutatása
A szerves vegyületek minőségi analízise – elemi összetételének meghatározása - során a minta kis részletének elégetésekor keletkezett égéstermékeket – gázokat, hamut - vizsgáljuk.
A szén és a hidrogén kimutatása
Bármely szerves vegyület elégetésekor az anyag széntartalma szén-dioxiddá, hidrogéntartalma vízzé oxidálódik.
A szén kimutatása során a fejlődő szén-dioxid gázt meszes vízbe vezetjük, mely a keletkező, rosszul oldódó kalcium-karbonáttól megzavarosodik.
CO2 + Ca(OH)2 = CaCO3 + H2O
https://drive.google.com/file/d/1QHnGBDcSzEZu6P78zppni4tkko1unvNE/view?usp=sharing
A hidrogén kimutatásakor a kémcső hidegebb részein lecsapódó páracseppek a szerves vegyületek hidrogénjének oxidációját jelzik.
A nitrogén kimutatása során a szerves vegyületek nitrogéntartalma nátrium-hidroxiddal ammóniává alakítható. A lúgos kémhatású ammónia keletkezését a lakmusz indikátor kékre változása jelzi.
https://drive.google.com/file/d/1Tu_10LRPIIrH5aAnLEMyQv-U1kgPea7p/view?usp=sharing
A kén kimutatása során a szerves vegyületek kéntartalmát nátrium-hidroxiddal melegítve szulfidionokká alakítjuk. A keletkezett szulfidionok ólom-nitráttal fekete ólom-szulfid csapadékká alakulnak.
S2- + Pb(NO3)2 = PbS +2 NO3-
https://drive.google.com/file/d/1PN9zMt5mqTprJfRZg-EKuLMbxvaMbA2q/view?usp=sharing
A vas kimutatásakor a hamuból a vasat salétromsavval kioldjuk, majd kálium-rodaniddal reagáltatjuk, melynek során vörös színű vas-rodanid keletkezik.
Fe(NO3)3 + 3 KSCN = Fe(SCN)3 + 3 KNO3
- Kémcsőben egy ujjnyi növényi hamut kb. 2 ujjnyi híg salétromsavval feloldunk.
- Enyhén melegítjük.
- Leszűrjük.
- A szűrlethez hozzáöntjük a kb. 1 ujjnyi reagenst.
https://drive.google.com/file/d/1VluZsl2Di-nalPYSGPwk5EHsl-t2xuqL/view?usp=sharing
Magyarázza és az élettani folyamatok elemzésén keresztül igazolja a víz alapvető biológiai funkcióinak jelentőségét.
A víz biológiai szerepe
Kitűnő poláris oldószer
-
- Poláris vegyületeket hidratációval,
- ionrácsos vegyületeket elektrolitos disszociációval,
- amfipatikus vegyületeket micellaképződéssel oldja.
Általában mindazon vegyületek jól oldódnak vízben, amelyek H-kötésre képesek.
Reakciópartner
- Hidrolízis során egy nagyobb molekula víz belépésével kisebb molekulákra bomlik. Makromolekulák monomerekre bomlásában van jelentősége, pl. az emésztésben.
- Kondenzáció során kisebb molekulák víz kilépésével nagyobb molekulákká egyesülnek. Monomerek makromolekulákká összekapcsolódásában jelentős, pl. fehérjeszintézisben.
Reakcióközeg, mivel az anyagcsere-folyamtok vizes oldatokban játszódnak le.
Szállítóközeg, vízben oldott anyagokat szállítja (pl. vér, növényi anyagszállítás).
Szerkezetmeghatározó
- A sejtek alakjának,
- növényi sejtek turgorának,
- lágyszárú növények, puhatestűek testalakjának meghatározója.
Hőszabályozás (hőkapacitás és párolgás)
A víznek nagy a hőkapacitása, ezért nagyon sok hőt képes elnyelni, tárolni és szállítani. Ennek jelentősége abban áll, hogy az élőlények testhőmérséklete a magas víztartalmuk miatt kevésbé ingadozik, akár a külső hőmérsékletváltozás okán, akár a hőtermelő (exoterm) anyagcsere folyamatok végett.
A párolgáshő egységnyi tömegű folyadék állandó hőmérsékleten történő elpárologtatásához szükséges energia. Nagy párolgáshőjének köszönhetően a víz sok hőt von el a környezetétől párolgása során, aminek többek között az élőlények – növények, állatok egyaránt – hőszabályozásában van jelentősége.
Tápanyag
A fotoszintézis egyik kiindulási vegyülete.
6CO2 + 6H2O + fényenergia = C6H12O6 (glükóz) + 6O2
Végezzen el a zsírok oldódásával kapcsolatos kísérletet, kapcsolja össze a zsírok biológiai szerepével.
A lipidek olyan szerves vegyületek gyűjtőcsoportja, amelyek igen különböző szerkezetűek, azonban közös sajátságuk, hogy apoláris oldószerekben jól oldódnak.
https://drive.google.com/file/d/1gt4E-3GP17LM6N5I4vrzkblL38bbFSVZ/view?usp=sharing
Neutrális zsírok (trigliceridek)
Ide tartoznak a természetes zsírok és a nem illó olajok. Apolárisak, vízben nem, alkoholban jól oldódnak.
Szobahőmérsékleten azokat a neutrális zsírokat, amelyek
- szilárdak, zsíroknak,
- folyékony halmazállapotúak, olajoknak nevezzük.
A neutrális zsírok kémiailag a glicerin zsírsavakkal alkotott észterei (triacil-gliceridek).
Glicerinből és zsírsavakból kondenzációval jönnek létre, melynek eredményeképpen a kapcsolódó csoportok között észterkötés jön létre.
A neutrális zsírok biológiai szerepe
1. Raktározott tápanyagok, kétszer annyi energiát tartalmaznak, mint az azonos tömegű szénhidrátok, mivel
- egyrészt vízmentesen tárolhatók, tekintve, hogy apolárisak,
- másrészt több hidrogén-szén kötést tartalmaznak, azaz redukáltabbak, mint a szénhidrátok, ezért oxidációjukkal több energia nyerhető.
Heterotróf szervezetekben elsődleges energiaraktárak. Állati szervezetekben a zsírszövetekben halmozódnak fel, pl. bőr aljában, szervek körül.
Növényekben pl. olajos magvakban találhatóak meg.
2. Helyzetüknél fogva - a bőr alatt - hőszigetelő szerepet látnak el, ill.
3. mechanikai szerepük van, pl. szervek helyét rögzítik, talpon puha párnákat képeznek.
4. Zsírban oldódó vitaminok (D, E, K, A) oldószerei és raktározó helyei.
Az állati szervezetekben minden feleslegben felvett tápanyag zsírrá alakul.
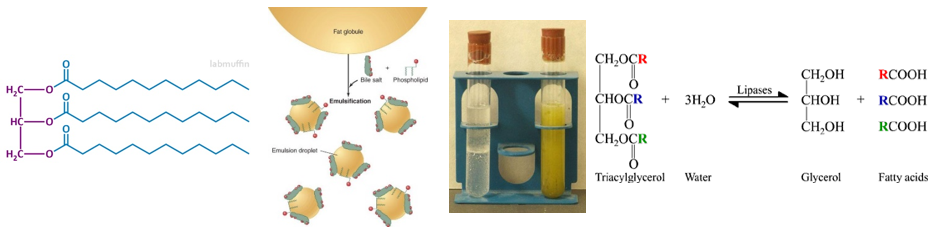 |
Magyarázza a kapcsolatot az epesav polaritása és az epesavas sók emulziót stabilizáló szerepe között, értelmezzen ezzel kapcsolatos kísérleteket.
Epesavak olyan szteroidok, amelyek az epében találhatók meg, sók formájában fordulnak elő, ezért az epe nem savas, hanem lúgos.
A középbélben segítik – nem végzik - a zsírok emésztését, úgy,
- egyrészt aktiválják a zsírbontó lipázokat,
- másfelől a bélmozgások miatt felaprózódott zsírcseppek felületén elhelyezkedve csökkentik az emulgeált zsírgolyócskák felületi feszültségét, stabilizálják azokat, és fajlagos felületüket nagyobbítva kedvezőbb feltételeket teremtenek a lipáz emésztés számára (hatásuk a szappanokéhoz hasonló).
Az epesavak amfipatikus vegyületek, apoláris részükkel (szteránváz) körbeveszik a zsírcseppeket, poláris részükkel (OH csoportok) pedig a vizes fázis felé fordulnak. Hatásuk a szappanokéhoz hasonlítható.
Az epe zsírokat szétoszlató szerepét bemutató kísérlet
- Két kémcsőbe öntsünk 3-3 ujjnyi vizet, mindkettőre rétegezzünk 1-1 ujjnyi olajat! Az egyikbe öntsünk egy ujjnyi epét!
- Gumidugóval zárjuk le a két kémcsövet, rázzuk össze azokat és várjunk néhány percig!
A víz poláris, az olaj apoláris, vízben nem oldódik, az epe kettős oldódási tulajdonságú. Az epét nem tartalmazó rendszer két fázisra vált szét, az epés kémcső zavaros maradt, mivel az epe apró cseppek formájában oldatban tartja a vízben nem oldódó olajat.
Értelmezzen a zsírok emésztésével kapcsolatos kísérletet.
A zsírok bomlása hidrolízissel történik, pl. az emésztés során lipáz enzim hatására a vékonybélben. A zsírok hatékony lebontásához nemcsak az enzimre (lipázra), hanem az epe mechanikai előkészítő munkájára is szükség van. A zsírok emésztése tehát egy összetett, kooperatív folyamat.
A zsírok bontásának vizsgálata lipáz és epe jelenlétében
Eszközök és anyagok
- Emulgeált zsírt tartalmazó oldat (pl. tej vagy krém)
- Lipáz enzim (pl. hasnyálmirigy-kivonat)
- Epeoldat
- pH-indikátor (pl. fenolftalein vagy univerzális indikátor)
- Kémcsövek, kémcsőtartó, vízfürdő (kb. 37 °C)
Kísérlet menete
Kémcső Összetevők
- 1. Zsíros oldat + víz (kontroll)
- 2. Zsíros oldat + lipáz
- 3. Zsíros oldat + epe
- 4. Zsíros oldat + epe + lipáz
Minden kémcsőhöz indikátort adunk, majd 37 °C-on inkubáljuk őket (kb. 15-30 percig), és figyeljük a színváltozást.
Tapasztalat
- Kémcső 1: Nem történik változás – sem enzim, sem epe nincs jelen, nincs emésztés.
- Kémcső 2: Kisebb pH-csökkenés (indikátor változása), lipáz elkezd dolgozni, zsírsavakat termel.
- Kémcső 3: Csak epe, nincs jelentős pH-változás, mivel emésztéshez enzim is kell.
- Kémcső 4: Jelentős pH-csökkenés, az epe emulgeálja a zsírt, a lipáz hatékonyabban működik, zsírsavak keletkeznek, a rendszer savasodik.
Értelmezés
- A lipáz a zsírokat bontja le zsírsavakra és glicerinre.
- Az epe (máj termeli, epehólyag tárolja) emulgeálja a zsírokat, azaz kisebb cseppekre bontja, ezáltal megnő az összfelület, amin az enzimek hatni tudnak.
- A reakció során keletkező zsírsavak savas kémhatásúak, ezt mutatja a pH csökkenése.
Értelmezzen szöveges leírás alapján a fehérjék szerkezetének megváltozásával kapcsolatos tulajdonságváltozásokat a prionok, a sarlósejtes vérszegénység példáján.
A prionok (PrPSc) olyan fertőző, kórokozó fehérjék, melyek különféle neurodegeneratív betegségek, mint pl. emberben a Jakob-Creutzfeldt betegség (fertőző szivacsos agyvelősorvadás), szarvasmarhákban a szivacsos agyvelősorvadás (kergemarhakór, szivacsos agyvelőgyulladás) kialakulásában játszanak szerepet. A kórfolyamat előrehaladtával a központi idegrendszer sejtjei elhalnak, az agyban üregek jönnek létre, állománya szivacsossá válik.
A prion betegségre jellemző, hogy nincs ellenanyag termelés, nincs gyulladásos reakció, a lappangási idő évekig tarthat.
A prionoknak nincs örökítő anyaguk, semmilyen életjelenséget nem mutatnak, a vírusoknál is egyszerűbb képződmények. A prionok az egyébként normálisan is meglevő sejtes fehérjék – celluláris prion (PrPc) – szerkezetileg módosult változatai.
A celluláris prionok (PrPc) általában idegsejtek felszínén a membránban találhatók, az idegsejtek egészséges működésében játszanak szerepet. A normál sejtes prionok vízoldékonyak, érzékenyek lebontó enzimekkel szemben.
A kóros prionok (PrPSc) aminosavsorrendje nem, de a másodlagos szerkezete – konformációja - eltér a normálisan is meglevő sejtes fehérjétől, s emiatt bontó enzimekkel szemben nem érzékenyek, vízben pedig nem oldódnak. Feltételezik, hogy a prionok (PrPSc) a sejtekbe jutva elősegítik – katalizálják - a normális fehérjék prionná átalakulását, melyek felhalmozódva, a hidrofób felszínükkel összecsapódva, oldhatatlan rögöket képeznek és az idegsejtek elhalását okozzák.
 |
A hemoglobin egyik közismert rendellenessége a sarlósejtes vérszegénység. A betegség oka, hogy egy pontmutáció miatt a globin egyik láncában egy aminosav kicserélődik, az N-terminálistól számítva a hatodik helyen glutamin helyett valin található.
A rendellenes vörösvértestek citoplazmájában a hemoglobin kicsapódik, kikristályosodik, a molekulák oxigénkötő képessége jelentősen csökken. A beteg sejtek egyrészt már csak részben képesek az oxigén szállítására (kb. 50%-ban), másrészt sarlóalakúvá válnak, összecsapódnak, vérrögöket képeznek, melyek elzárhatnak kisebb ereket. A hibás sejteket a szervezet – a lép - 10-20 nap múlva kivonja a forgalomból, ill. mivel ezek a deformált sejtek törékenyek, a kisebb ereken való áthaladásuk során szétesnek, ezért a vörösvértestszám lecsökken, ami vérszegénységhez vezet. Az oxigénellátási zavarokon túl a betegség szövődményei pl. stroke, tüdőembólia, melyek többnyire az érelzáródásokkal hozhatók kapcsolatba.
Magyarázza a fehérjék kimutatását biuret-reakcióval.
A fehérjék kimutatása: biuret-próba
A biuret-próbával a fehérjékben jelenlévő amidcsoportot lehet kimutatni. Pozitív próba esetén ibolyaszíneződést tapasztalunk, mivel a reagensben található rézionok komplexet képeznek az amidcsoporttal.
Egy kevés fehérjeoldathoz adjunk néhány csepp 40%-os nátrium-hidroxid-oldatot, majd 1-2 csepp réz(II)-szulfát reagenst.
Elemezzen kísérleteket a DNS örökítő szerepének bizonyításával kapcsolatban (Griffith és Avery, Hershey és Chase kísérlete).
A DNS örökítő szerepét bizonyító kísérletek
Az öröklődés anyagi hátterének tisztázásához – az örökítőanyag felfedezéséhez - vezető út első állomása Fred Griffith katonaorvos, bakteriológus nevéhez fűződik, aki az I. világháború után 1928-ban baktériumok okozta tüdőgyulladás elleni vakcina fejlesztésén dolgozott.
Kísérleteiben egy gömb alakú baktérium - Diplococcus pneumoniae - kórokozó képességét – patogenitását – vizsgálta, amely egérben vérmérgezést, emberben tüdőgyulladást okoz.
E baktériumnak két törzse ismert.
- Patogén (sima felszínű, S-törzs), amelynek egyedei az immunrendszernek ellenálló poliszacharid tokot fejlesztenek maguk köré. A tokképzés örökletes, azaz az utódok is tokosak lesznek.
- Nem patogén (ráncos felületű, R-törzs), amelyek nem képeznek tokot, s emiatt a sejteket az immunrendszer meg tudja semmisíteni. Ez is természetesen örökletes tulajdonság.
Griffith azt vizsgálta, hogy ezek a tulajdonságok vajon megváltoztathatatlan tulajdonságai ezeknek a törzseknek, vagy esetleg a két törzs egymásba átalakulhat.
Griffith a baktériumokat egerekbe oltotta, majd a következő eredményeket kapta:
- Egér + élő tok nélküli baktérium = az egerek nem pusztultak el.
- Egér + élő tokos baktérium = az egerek 1-2 napon belül elpusztultak.
- Egér + hővel elölt tokos baktérium = az egerek nem pusztultak el.
- Egér + hővel elölt tokos + élő tok nélküli baktérium = néhány egér elpusztult!
Az elpusztult állatok hulláit megvizsgálták és meglepődve tapasztalták, hogy bennük élő tokos baktériumok voltak. Griffith tehát azt a váratlan felfedezést tette, hogy az elölt patogén sejtek az élő, nem fertőző sejtekhez keverve, azokat patogénné tették, azaz megváltoztatták.
Griffith arra a következtetésre jutott, hogy az elölt S-törzsből valamilyen hőnek ellenálló, információt hordozó anyag átjut - ún. transzformációs faktor - az élő R-törzs sejtjeibe, és a sejtek egy részét átalakítja, transzformálja.
Griffith tehát az örökítőanyag létét bizonyította, amely a hőhatás után átkerülve a nem kórokozó sejtekbe, azokat tok képzésére serkentette.
Ezt a folyamatot genetikai transzformációnak nevezzük. Griffith szerint tehát a baktérium - általa nem ismert – örökítőanyaga okozta a jelenséget, ugyanakkor maga nem próbálta mindenáron meghatározni ezt az anyagot.
 |
Az örökítőanyag meghatározását Oswald Avery végezte el 1941-ben.
A hővel elölt kórokozó S-törzs baktériumaiból sejtmentes kivonatokat készítettek, melyeket egyenként RNS-t, fehérjét, illetve DNS-t emésztő enzimekkel kezelték.
A különféle enzimekkel kezelt S kivonathoz R-variáns baktériumtenyészetet kevertek, majd megvizsgálták, hogy találnak-e köztük transzformált baktériumokat.
Azt találták, hogy a baktériumkivonatból
- fehérjék (proteáz) és
- RNS-ek (RNáz) kivonhatók anélkül, hogy a transzformáció mértéke lényegesen csökkenne.
- Azonban, amikor DNS-bontó enzimekkel – DNáz - kezelték a kivonatot a transzformáció elmaradt.
Tehát a transzformációért felelős anyag minden kétséget kizáróan a DNS, mivel a transzformáló képességet DNáz- készítménnyel való kezelés megszüntette.
 |
1952-ben Alfred Hershey és Martha Chase baktériumokat fertőző és pusztító vírusokkal, ún. bakteriofágokkal végeztek vizsgálatokat.
Mint minden vírus, a fágok is alapvetően két részből állnak:
- a központi információtároló DNS-ből,
- és az ezt körülvevő fehérjetokból, a kapszidból.
Röviden a fertőzés úgy történik, hogy
- a fág hozzátapad a baktérium felületéhez,
- egy enzim feloldja a baktériumsejt falát, s az így keletkezett nyíláson a DNS behatol a baktériumsejtbe. A fág fehérjeburka a sejten kívül marad.
- A bejutott DNS többszöröződik a sejt anyag- és energiakészletét felhasználva.
- A létrejött fágok érése után az elpusztult baktériumsejt szétszakad és mintegy 1OO-5OO új fertőzőképes fág szabadul ki belőle.
A kutatók olyan fágokkal kísérleteztek, melyek fehérjéit radioaktív kénnel, DNS-ét radioaktív foszforral jelölték meg. Az így jelölt fágokkal baktériumsejteket fertőztek.
Rövid idő után rázással a fágokat eltávolították a baktérium felületéről, majd az elegyet nagy sebességgel forgó berendezésbe helyezték, centrifugálták.
A nagyobb tömegű baktériumsejtek a centrifugacsövek aljára ülepedtek, a sejten kívül maradt könnyű vírusrészecskék a folyadék felszínén gyűltek össze.
Ezt követően megmérték az elegyek aktivitását. A fentieknek megfelelően
- a radioaktív kénnel (S) jelölt vírus esetében a centrifugálás után a folyadék felszínén mértek erős radioaktivitást,
- ugyanakkor a radioaktív foszforral (P) jelölt vírus esetében a centrifugálás után a cső alján mértek erős radioaktivitást.
Mivel a radioaktív foszfort tartalmazó vírusok által megfertőzött baktériumok a rázást követően mutatták a radioaktivitást, a radioaktív kénnel megfertőzött baktériumok a rázást követően pedig nem, egyértelmű volt, hogy a DNS jutott be a sejtekbe és szervezte át a baktériumok működését.
Kapcsolja össze a DNS duplikáció folyamatát a polimeráz láncreakció (PCR) technológiai módszerrel, magyarázza a módszer lényegét, értelmezze e módszer szerepét az orvosi diagnosztikában.
A DNS megkettőződése, a replikáció
A sejtek osztódásához szükséges a genetikai információ pontos megkettőződése és átadása az utódsejteknek, ez biztosítja ugyanis a biológiai információ lényegében változatlan megőrzését az utódnemzedékben. A DNS megkettőződés során a DNS-molekula pontos másolata jön létre.
A DNS-molekulában észlelhető szabályosság, a bázisok megfelelő kapcsolódása (komplementaritása) képezi az alapját a genetikai információ egyértelműen pontos másolásának, a replikációnak (adeninnel szemben mindig timin van, citozinnal szemben pedig guanin helyezkedik el).
A replikáció alatt a DNS-molekula két szála elválik egymástól és az így különváló láncok mintaként szolgálnak egy-egy újabb DNS-szál szintézisének.
Ennek megfelelően az utód DNS-molekulák egyik lánca a szülői DNS-ből származik, a másik komplementer lánc újonnan szintetizálódik hozzá (szemikonzervatív – félig megmaradó - jelleg).
A replikáció mechanizmusa
A replikáció a DNS-szál szigorúan meghatározott részén, az ún. kezdő – iniciációs - ponton indul meg, amit replikációs origónak nevezünk.
- A másolást megelőzően a kromoszómaszakaszhoz kicsavarófehérjék – helikázok - kapcsolódnak. A folyamatban résztvevő enzimek felszakítják a bázisok közötti H-kötéseket, aminek következtében a DNS-szálak elválnak egymástól.
- A szabaddá vált DNS-szakaszon egy RNS-polimeráz (primáz) segítségével megindul a DNS-száléval komplementer, 50-100 ribonukleotid egységből RNS-primer szintézise.
- A primer RNS 3'-OH végéhez a DNS-polimeráz III lépésenként dezoxiribonukleotid-trifoszfátokat A folyamat energiaigényét a lehasadó difoszfátok hidrolízise biztosítja.
- Az új szál tehát 5' - 3' irányba nő. Ahogy az új szál hosszabbodik, a kicsavaró fehérjék elválasztják a komplementer szálakat, kialakítják a replikációs villa újabb szakaszait.
- Mivel a DNS két szála ellentétes polaritású, úgy szintézisük is ellentétes irányba történik, hiszen a szintézis iránya mindig 5’ - 3’.
Minthogy a kicsavarófehérjék csak az egyik irányba haladnak, azaz a replikációs villa csak az egyik irányba nyílik szét, csak az egyik szál szintézise lehet folyamatos, az, amelynek növekedési iránya megegyezik a helikázok haladási irányával. Ebből következik, hogy a másik ellenkező irányba növő szál szintézise csak szakaszosan mehet végbe (Okazaki-fragmentek).
Az ellenkező irányba növekvő szálon a szintézis 1000-2000 RNS nukleotid egység összekapcsolásáig tart. Az RNS- primert a polimeráz I távolítja el, majd helyére komplementer dezoxiribonukleotidokat épít (kitölti a hézagokat).
Az egyes DNS-szakaszokat a ligáz kapcsolja össze úgy, hogy két nukleotid egység között foszfodiészter kötés kialakulását katalizálja.
https://drive.google.com/file/d/0B15KJwYa2uMFeDJsQjJQZHdJUEk/view?usp=sharing
 |
A DNS sokszorozása, a PCR technika (1993 Nobel díj)
A szekvenáláshoz a vizsgálni kívánt DNS-szakaszt sokszorozni kell. Ma már számos módszer ismert különböző méretű DNS-szakaszok sokszorosítására. Ezek közül talán a leginkább elterjedt a polimeráz láncreakció, a PCR, amely forradalmasította a géntechnológiát, a molekuláris diagnosztikát.
Az eljáráshoz szükségesek:
- DNS-minta.
- Két (DNS) primer, amely szükséges a reakció megindulásához. A primerek rövid, mesterséges DNS-szálak, amelyek komplementerek a sokszorosítandó DNS-szakasz elejével és végével. Hozzákapcsolódnak a DNS-mintához, ahová a DNS-polimeráz kötődik, ami majd megkezdi egy új DNS-szál szintetizálását.
- DNS-polimeráz enzim – amely lemásolja a DNS-t.
- Nukleotidok – amelyekből a DNS-polimeráz felépíti az új DNS-t.
- Pufferoldat, amely biztosítja az enzim számára megfelelő körülményeket.
A PCR-eljárás húsz-harminc ciklus egymásutánjából áll. Mindegyik ciklus három lépést tartalmaz.
- A minta DNS-t magas hőmérsékletre (95 oC) kell hevíteni, ekkor a DNS két szála különválik.
- A hőmérséklet csökkentését (kb. 68 oC) követően a primerek a DNS-szál elejéhez kapcsolódnak.
- Ismét magasabb hőmérsékleten (72 oC) a hőrezisztens polimeráz enzim a primerektől elindulva megkezdi a mintaszállal komplementer DNS szintézisét.
A ciklus végére a kiindulási DNS-szakasz mennyisége megkétszereződik.
A DNS sokszorosítás (PCR) felhasználási területei, szerepe az orvosi diagnosztikában
Az alábbi eljárások alkalmazásához a felhasznált DNS-mintát sokszorosítani szükséges.
- Az igazságügyi orvosszakértői munkában újabban a gyanúsított személyt genetikai ujjlenyomata alapján azonosítják, úgy, hogy a DNS-ét a bűncselekmény helyszínéről származó minta DNS-ével hasonlítják össze. Nem szükséges, hogy a minta sok DNS-t tartalmazzon, elméletileg egyetlen DNS-szál is elegendő.
- Az örökletes és szerzett mutációk vizsgálatához szükséges a DNS-minta megsokszorozása. A géndiagnosztikai eljárásokkal azonosíthatók a rosszindulatú daganatok kialakulásáért felelős gének, mint pl. a BRCA-1 és BRCA-2 gének, melyek mutációja a mell-, illetve a petefészekrák kialakulásában jelentős. A CRISPR módszerrel egyrészt tumorszupresszor gének aktiválhatók, másrészt onkogének kikapcsolhatók a CRISPR technikával bevitt pontmutációk kiváltásával.
- Egyes vírusok (pl. HPV, HIV, HCV, koronavírus stb.) és baktériumok által okozott fertőzések kimutatásához a fertőzést okozó mikroorganizmus DNS-ét sokszorosítani kell. A mikroorganizmusok ellen termelődött antitestek kimutatásán alapuló immunodiagnosztikai vizsgálatokkal szemben a PCR lehetővé teszi a fertőzés kimutatását akkor is, ha az immunválasz valamiért elmarad.
- DNS szekvenáláshoz.
Néhány példa arra, hogy a genetikai diagnosztikában mely betegségek esetén használatosak DNS-szekvenálási módszerek:
- cisztás fibrózis,
- hemofília, fenilketonúria,
- örökletes emlőrák (BRCA-1, BRCA-2) stb.
Más betegségeknél több, a betegségre hajlamosító gének vizsgálatát végzik molekuláris genetikai módszerrel. Ezen rizikófaktorok ismeretében felmérhető a betegség kialakulásának kockázata. Például: Alzheimer kór, laktózérzékenység, csontritkulás, trombózis (több gén együttes vizsgálata).
A gyakori betegségek genetikai vizsgálatának legfontosabb jelenlegi módszere a genom-asszociációs analízis. Az eljárás alapjául az a tapasztalat szolgál, hogy egy kromoszóma szakasz bizonyos variánsa nagyobb gyakorisággal fordul elő betegekben, mint az egészségesek között. Ennek az az oka, hogy a megfigyelt szakasz olyan gént vagy génműködést szabályozó DNS-t hordoz, amely hatással van a betegség kialakulására. Az elemzés során a vizsgált DNS-szakaszt összehasonlítják egészséges emberekből vett DNS-minták megfelelő régióival.
2.2. Az anyagcsere folyamatai |
Végezzen el vizsgálatot/kísérletet egy vízinövény fotoszintézisével összefüggésben, magyarázza a tapasztalatokat.
Tervezzen és értelmezzen kísérletet a fotoszintézist befolyásoló tényezők fotoszintézisre gyakorolt hatásának és a fotoszintézis végtermékeinek bemutatására.
A fotoszintézis vizsgálata vízinövényen (pl. békalencse vagy kanadai átokhínár)
6CO2 + 6H2O = C6H12O6 (glükóz) + 6O2
Kutatási kérdés
Hogyan befolyásolja a fényerősség a vízinövény fotoszintézisét, a képződő oxigénbuborékok számának mérésén keresztül.
Szükséges eszközök és anyagok
- Békalencse vagy egy szál kanadai átokhínár
- Főzőpohár vagy kémcső vízzel
- Asztali lámpa (állítható távolságú)
- Vonalzó (a lámpa távolságának méréséhez)
- Stopperóra vagy időzítő
- Szódabikarbóna (kis mennyiség, szén-dioxidforrásként)
- Jegyzetfüzet a méréshez
A kísérlet menete
- Helyezzük az átokhínárt vízzel teli főzőpohárba.
- Adjunk hozzá egy kevés szódabikarbónát (szén-dioxid biztosítása céljából).
- Világítsuk meg a növényt az asztali lámpával.
- Állítsuk be a lámpát 10 cm-re a növénytől.
- Indítsuk el a stoppert, és 1 percen keresztül számoljuk a kibocsátott oxigénbuborékokat (a fotoszintézis mellékterméke).
- Ismételjük meg a mérést más távolságokon is: 20 cm, 30 cm, 40 cm stb.
Eredmények
Lámpa távolsága (cm) Buborékok száma / perc
10 32
20 20
30 11
40 5
Tapasztalatok és értelmezés
- A fényintenzitás növekedésével nőtt a fotoszintézis sebessége (több oxigénbuborék).
- Azt tapasztaljuk, hogy a fotoszintézis sebessége egy adott határig arányos a fény intenzitásával.
- A csökkenő fényerősség (nagyobb lámpatávolság) csökkenti a fotoszintézis sebességét.
 |
Végezzen el vizsgálatot/kísérletet egy vízinövény fotoszintézisével összefüggésben, magyarázza a tapasztalatokat.
A fotoszintézis vizsgálata vízinövényen (pl. tündérhínár)
Kutatási kérdés
Hogyan befolyásolja a szén-dioxid-koncentráció a vízinövény fotoszintézisét, a képződő oxigéngáz mennyiségének mérésén keresztül.
Szükséges eszközök és anyagok
- Egy szál kanadai átokhínár
- Főzőpohár vagy kémcső vízzel
- Lámpa
- Különböző koncentrációjú nátrium-hidrogénkarbonát-oldat
- Stopperóra vagy időzítő
- Jegyzetfüzet a méréshez
A kísérlet menete
- Helyezzük az tündérhínárt vízzel teli fecskendőbe.
- Adjunk hozzá adott koncentrációjú szódabikarbónát (szén-dioxid biztosítása céljából).
- Világítsuk meg a növényt lámpával.
- Indítsuk el a stoppert, minden oldat esetén mérjük, hogy mennyi idő volt szükséges, hogy a folyadék elérje az 5 cm-es jelzést a kapilláriscsőben.
- Ismételjük meg a mérést különböző koncentrációjú nátrium-hidrogénkarbonát-oldatba helyezve a növényt.
Eredmények rögzítése
Tapasztalatok és értelmezés
- A szén-dioxid-koncentráció növekedésével nőtt a fotoszintézis sebessége (a folyadék gyorsabban érte el az 5 cm-es jelzést a kapilláris csőben).
- Azt tapasztaljuk, hogy a fotoszintézis sebessége egy adott határig arányos a szén-dioxid-koncentrációval.
A fotoszintézis intenzitását tehát befolyásolja a fényintenzitás, ill. a szén-dioxid-koncentráció. E két tényező növekedése egy adott érték felett már nem tudja fokozni a fotoszintézis mértékét, mivel a folyamatok elérték kapacitásuk felső határát.
Fotoszintézis animáció
https://www.youtube.com/watch?v=gnNDC1aOBU0
Videó
https://www.youtube.com/watch?v=sQjnne6wloA&t=10s
https://www.youtube.com/watch?v=nqBKGHN8HeE&t=15s
https://www.youtube.com/watch?v=OcH1UVg3gto
Elemezzen az erjedéssel és a biológiai oxidációval kapcsolatos kísérleteket, esettanulmányokat, tervezzen a folyamatokkal kapcsolatos kísérleteket.
A biológiai oxidáció során keletkező CO₂ kimutatása és összehasonlítása élő és elhalt magok között.
C6H12O6 + 6 O2 = 6 CO2 + 6 H2O
Eszközök
- Csírázó borsó vagy bab
- Főtt (elölt) magvak (kontrollcsoport)
- Légmentesen zárható lombik
- Meszesvíz (Ca(OH)₂-oldat)
- Gumidugó csövezéssel
- Állvány, kémcső, vízfürdő
A kísérlet menete
- Két lombikba helyezünk egy-egy adagot: az egyikbe csírázó, a másikba főtt magvakat.
- Mindkettőhöz kapcsolunk egy külön kémcsövet meszesvízzel.
- A lombikokat melegen tartjuk (~30 °C).
- Néhány óra után megfigyeljük a meszesvíz zavarosodását (kalcium-karbonát kiválása).
Tapasztalatok, értelmezés
- A csírázó magvak lombikjához kapcsolt mészvíz zavaros lesz, míg a főtt magvaké nem.
- Ez a CO₂ jelenlétére utal, tehát a csírázó magvak sejtlégzést végeznek.
https://www.youtube.com/watch?v=iHr1Kb8KND4
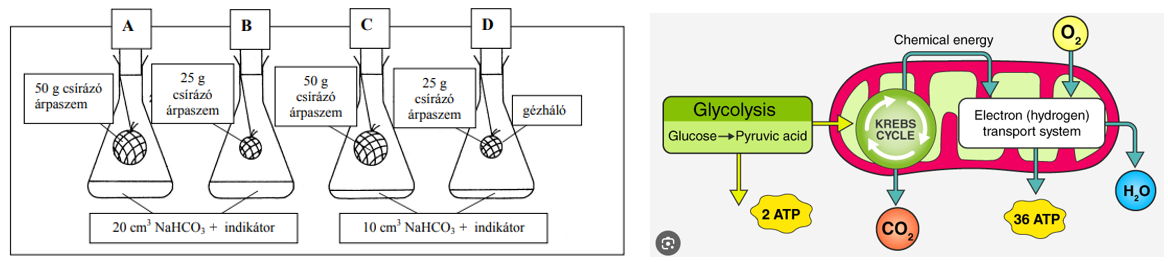 |
Kutatók csírázó magvak anyagcsere-folyamatait fény kizárása mellett vizsgálták. Árpaszemeket két napig Petri-csészében csíráztattak, majd egy részüket az ábrán látható módon nedves gézbe csomagolva felfüggesztették egy dugóval lezárt lombikban. A lombikba töltött lúgos folyadék olyan indikátort tartalmazott, mely savasodás hatására pirosból sárga színre vált. Négy kísérleti berendezést készítettek a rajzon föltüntetett anyagmennyiségekkel szobahőmérsékleten.
A biológiai oxidáció során felszabaduló szén-dioxid oldódik a NaHCO3 oldatban, közömbösítve, ill. elsavasítva azt, amit az indikátor színének a megváltozása jelez. A C jelű lombikban tapasztaljuk először a változást, hiszen ebben található a legtöbb csírázó árpaszem, itt termelődik több CO2 ill. kisebb mennyiségben van jelen a lúg, amit közömbösíteni kell.
Az erjedés vizsgálata
Különféle heterotróf mikroorganizmusokban, baktériumokban, ill. élesztőgombákban anaerob körülmények között alkoholos erjedés zajlik, melynek során a glikolízisben keletkezett piroszőlősavból szén-dioxid kiválása mellett, NADH redukciójával etil-alkohol keletkezik. Alkoholos erjedés zajlik a bor keletkezésénél és a tészta kelésénél egyaránt.
C6H12O6 + = 2 CH3CH2OH + 2 CO2
Egy függőlegesen tartott kémcsőbe 5 cm3 glükóz-oldatot öntünk. Ebbe egy darabka élesztőt teszünk, majd a kémcsövet 37 ºC-os vízfürdőbe helyezzük. A kémcsövet egy kifúrt dugóval bedugjuk, amelynek furatába egy „U”-alakban meghajlított üvegcsövet illesztünk. Az üvegcső másik végét egy telített meszes vizet tartalmazó kémcsőbe merítjük. Figyeljük a változást.
Egy idő után a folyamat során keletkezett szén-dioxidtól a meszes víz zavarossá válik.
https://www.youtube.com/watch?v=yKfpfv0ye5A&t=12s
2.3. Sejtalkotók (az eukarióta sejtben) |
Ismerje föl mikroszkópban és mikroszkópos képeken a sejtfalat, zöld színtestet, sejtmagot, zárványt.
 |
Mutassa be ábra vagy szöveg segítségével a fehérjék lehetséges transzport útvonalait a sejten belül: az elválasztott fehérjék, az intracelluláris emésztés és a membránfehérjék példáján egy radioaktívan jelölt aminosav útjának nyomon követésével.
A fehérjék sejten belüli transzportja bonyolult, de jól szervezett folyamat, amely során a fehérjék szintézisük után különböző útvonalakon keresztül jutnak el rendeltetési helyükre. Ezt a folyamatot jól lehet tanulmányozni radioaktívan jelölt aminosavak (pl. 35S-metionin) felhasználásával, amelyek beépülnek az újonnan szintetizált fehérjékbe, lehetővé téve azok útjának követését.
A durva felszínű endoplazmatikus retikulum RER (DER)
A RER-t lapos zsákok bonyolult hálózata építi fel, amely kiterjedt kapcsolatban áll a sejtmaghártyával. Felületén riboszómák találhatók, amelytől felszíne szemcséssé válik – innen az elnevezés.
A RER, a sejtből kikerülő ún. exportfehérjék, ill. a membránfehérjék keletkezésének a helye. Feladatából adódóan elsősorban intenzív fehérjeszintézist végző sejtekben található meg nagy mennyiségben, pl. mirigysejtekben, idegsejtekben.
Az exportfehérjék szintézise
Az elkészült mRNS a citoplazmába kerül, ahol szabad riboszómákkal kapcsolódik.
Az exportra kerülő fehérjék esetén a meginduló fehérjeszintézisnek köszönhetően a peptidlánc végén először egy ún. szignálpeptid jelenik meg, ami az endoplazmatikus retikulum felszínére irányítja a riboszómát.
Az endoplazmatikus hálózat riboszómáin elkészült fehérje a RER üregébe kerül, ahol további változásokon megy keresztül. A módosítások lényege, hogy egyes aminosavakhoz különféle oligoszacharidok kapcsolódnak.
A RER-ben elkészült fehérje a Golgi-készülékbe jut, ahol újabb átalakuláson megy keresztül.
 |
A Golgi-készülék
A Golgi-hálózat egymással párhuzamosan rendeződött lapos zsákokból áll, amelyek széléről folyamatosan apró hólyagok, ún. Golgi-hólyagok (vezikulák) fűződnek le.
Golgi-apparátus feladata
- a RER-ben szintetizálódott váladék- és membránfehérjék fogadása,
- a szénhidrátok fehérjékhez történő kapcsolódása.
- A membránfehérjék transzportja.
- Mirigysejtekben az elválasztásra kerülő fehérjék becsomagolása.
- Lizoszómák előállítása.
A sejten belüli emésztés
A sejt belsejébe membránba burkoltan érkező, a fagoszómákban elhelyezkedő anyag – baktériumok, makromolekulák – megemésztését hidrolázokat tartalmazó savas közegű lizoszómák végzik. A lizoszómák a Golgi-készülékből lefűződéssel keletkeznek.
- Első lépésben ún. primer lizoszómák keletkeznek. Ezek hidrolitikus, bontó enzimeket tartalmaznak.
- A fagoszóma a sejtbe kerülve összeolvad a primer lizoszómákkal és létrehozza a szekunder lizoszómát (emésztő űröcskét), amelyben megkezdődhet az emésztés. A makromolekulák addig emésztődnek, amíg a keletkezett anyagok olyan méretűvé válnak, amelyek már transzportfolyamatokkal képesek átjutni a membránon.
- Az emésztési folyamat következtében a lizoszómában már csak az emészthetetlen anyagok maradnak vissza. Ekkor már tercier lizoszómáról beszélünk. Ez előbb vagy utóbb a plazmamembránhoz vándorol és exocitózis révén tartalmát a sejten kívüli térbe juttatja.
Értelmezzen ábrán és szövegben bemutatott komplex transzportfolyamatokat a glükóz emberi vékonybélben zajló felszívódása példáján.
Glükóz-Na+ kotranszport (csatolt anyagtranszport, szimport), a bélhámsejtek glükózfelvétele
Az aktív transzportfolyamatok egyenlőtlen anyageloszlást hoznak létre a membrán két oldala között. Mivel az anyag a koncentrációgradiens által meghatározott spontán mozgási iránnyal szemben mozog, az aktív transzport mindig E-t igényel.
Az energia ATP hidrolíziséből származik. Az aktív transzport mindig szállítókkal történő transzport. Energiaigényes szállítófolyamatok pl. a különféle ionokat szállító pumpamechanizmusok.
A membránpumpák olyan transzportrendszerek, amelyek egy adott anyagot a membrán egyik oldalán felhalmoznak, míg a membrán másik oldalán a folyadék az adott anyagban elszegényedik. A ma ismert pumpamechanizmusok ionszállító transzportrendszerek, amelyek egyben ATP-bontó enzimek is.
Ismertek olyan szállítók a membránban, amelyek kétféle anyagot is képesek egyszerre átjuttatni a sejthártyán. Ebben az esetben csatolt transzportról – kotranszport - beszélünk. Ilyen módon veszik fel a tápcsatornában a bélhámsejtek a glükózt.
A glükóz felvétele aktív transzport, az alacsonyabb koncentrációjú hely felől a magasabb koncentrációjú hely felé szállítódik, amihez az energiát a Na+ koncentrációgradiense által meghatározott diffúziója biztosítja. Tehát a glükóz energetikailag kedvezőtlen irányba megy, a Na+ pedig kedvezőbe, de végső soron a kettő eredője kedvező.
A mechanizmus másodlagos aktív transzport, hiszen a folyamat feltétele a Na+ koncentrációkülönbsége a sejthártya két oldalán, melyet a Na+-K+ pumpa hoz létre ATP felhasználásával.
https://www.youtube.com/watch?v=nsggCqbsVxs
 |
Elemezze ábrán a gyökérszőrök ionfelvételét, a sejtek inzulin hatására történő glükózfelvételét.
A víz és a sók felvételének módja a hajtásos növényekben, a gyökérszőrök anyagforgalma
A gyökérszőrök sejthártyája féligáteresztő sajátosságú, azon a víz passzívan, ozmózissal beáramolhat, mivel a gyökérszőr sejtjeinek a citoplazmája töményebb, mint a talajoldat, így sejtplazma egyfajta szívóerőt gyakorol a gyökérsejtek körüli vízmolekulákra. A beáramlott víz növeli a sejten belüli nyomást, turgor állapotot létrehozva, ezért a víz átpréselődik a szomszédos alapszöveti sejtekbe, majd onnan a farész vízszállító csöveibe (gyökérnyomást kialakítva).
A szükséges ionokat a gyökérszőrök sejthártyája válogatja ki a növény szükségleteinek megfelelően, majd a koncentráció viszonyoktól függően (esetleg passzív de) főleg aktív transzporttal a sejt belsejébe juttatja. Az energiaigényes transzporttal az ionok a nagyobb koncentráció irányába is szállítódhatnak.
A bőrszövettől a szállítószövetek farészéig a sók transzportja révén egy egyre növekvő koncentrációgradiens biztosítja a víz mozgását az ozmózis szabályainak megfelelően.
Tehát a gyökérben a turgornyomás és a szomszédos sejtek egyre növekvő szívóereje a vizet bepréseli a farészbe, majd onnan a szárba, amelynek eredményeképpen egy ún. gyökérnyomás alakul ki.
A gyökérnyomás tehát az ozmózis hatására létrejövő a hajtóerő, amely a vizet a szállítónyalábokba préseli. Fákban néhány méter magasságig képes a vizet felnyomni.
https://www.youtube.com/watch?v=L5sEm56qbDc
Facilitált – könnyített - diffúzió, a sejtek inzulin hatására történő glükózfelvétele
A szállított anyag mindig a koncentrációgradiense által megszabott irányba mozog diffúzióval, passzív transzporttal, szállítók segítségével. A szállított anyag így gyorsabban jut át, mint a molekula méretéből, lipidoldékonyságából, koncentráció-gradienséből következne.
A sejtbe bejutott anyagok igen gyorsan hígulnak, ami biztosítja a folyamatok egyirányúságát. Pl. a glükóz gyorsan foszforilálódik, a glükóz-foszfát a szállítóval már nem kapcsolódik.
Segített diffúzióval veszik fel a különféle szöveti sejtek a glükózt. Többféle – 14 - glükóztranszporter ismert (GLUT).
- A GLUT-1 a legtöbb szövetben – pl. agy, vörösvértestek, vastagbél - kis mennyiségben található meg, inzulintól független biztosítja a glükóz felvételét.
- Az inzulinfüggő GLUT-4 többek között izom- és zsírszövetben fordul elő.
Inzulin hatására a GLUT-1 transzporterek mellé, az addig a sejt belsejében, vezikulákban tárolt GLUT-4 transzporterek beleolvadnak a külső membránba, s megnövelik a zsírsejtek és az izomrostok glükózfelvételét.
https://www.youtube.com/watch?v=r69MPM0_5bQ
Rakja sorrendbe a sejtosztódás szakaszait rajzolt ábrák vagy képek alapján, párosítsa a szakaszokat a bennük zajló folyamatokhoz.
A mitózis
A mitózis fonalas sejtosztódást jelent (mitosz = fonál). A mitózis, más néven számtartó sejtosztódás lényege, hogy az interfázisban megkettőződött DNS - melyek a kromoszómák egyes kromatidáit alkotják – egyenlő mértékben megoszlik a két utódsejt között, két utódsejtmag jön létre, amelyekben ugyanolyan számú kromoszóma van, mint az osztódás előtti magban volt. A számtartó sejtmagosztódás bonyolult, általában néhány óra alatt lejátszódó folyamat.
Mitózissal osztódnak:
- a növények testi sejtjei,
- állatok testi sejtjei, azaz a zigóta és az abból létrejövő sejtek.
A mitózis folyamatán belül 4 fázist különböztetünk meg.
Előszakasz, profázis
- A sejtmagvacska eltűnik, a maghártya kezd hólyagokká feldarabolódni.
- A kromatin fokozódó tömörödése miatt láthatók lesznek a kromoszómák.
- A már kettéosztódott sejtközpontok a sejt két ellentétes pólusára vándorolnak.
Középszakasz, metafázis
- Ekkorra a kromoszómák a legnagyobb mértékben rövidülnek,
- a kromoszómákat a sejt középső síkjában rendezi el,
- a kromoszómák centromérjeihez húzófonalak kapcsolódnak, ez a magorsó.
Utószakasz, anafázis
- A húzófonalak a kromatidákat a sejt két pólusára mozgatják.
Végszakasz, telofázis
- A pólusokra érkezett kromatidák fokozatosan kitekerednek,
- a magorsó eltűnik, új maghártya szerveződik.
Az örökítő anyag szétosztását a citoplazma osztódása követi, amelynek során a sejthártya befűződik, a sejt kettéválik, a sejtszervecskék kb. egyenlő mértékben elosztódnak az utódsejtek között. A folyamat a citokinézis.
Minden mitózis után két utódsejt keletkezik, melyek genetikailag azonosak egymással és az anyasejttel (mutációtól, másolási hibáktól eltekintve).
Genetikai szempontból legfontosabb eseménye az, hogy a két kromatidából álló kromoszómák kromatidái (DNS kettős hélixei) az osztódás során szétválnak, majd az utódsejtekbe kerülnek. Az utódsejtekben tehát ismét egy kromatidából (egy DNS kettős hélixből) álló, de változatlan kromoszómaszámú kromoszómakészlet található.
https://www.youtube.com/watch?v=VlN7K1-9QB0
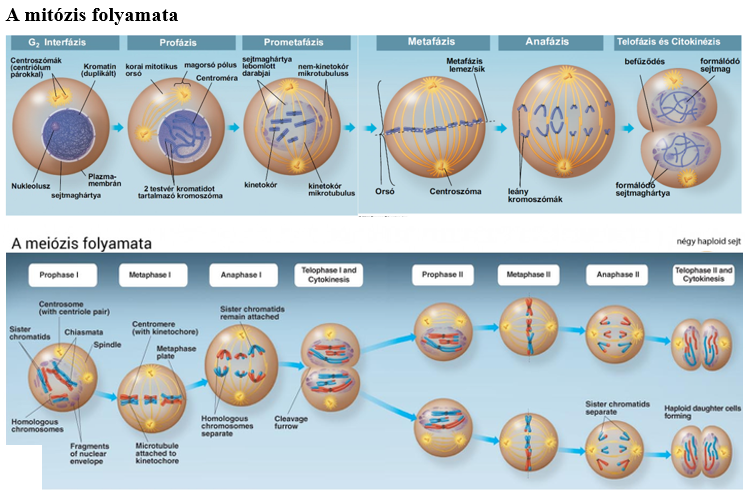 |
Meiózis
- A meiózis az állati ivarsejtek, növényi, ill. gombaspórák termelésére specializálódott diploid sejtek osztódástípusa.
- A meiózis során egyetlen diploid anyasejtből 4 haploid utódsejt keletkezik.
- Az utódsejtek kromoszómaszáma - ezáltal genetikai információtartalma - pontosan fele az anyasejtének (számfelező sejtosztódás).
- A keletkezett négy haploid utódsejt információtartalma egymástól különböző.
A meiózis két főszakaszra bontható.
- A meiózis első szakaszának profázisában a homológ kromoszómák párt alkotnak, kromatidáik között átfedések, törések, majd szakasz – gén - kicserélődések következhetnek be. Ennek a folyamatnak, az átkereszteződésnek (crossing over) az eredménye az intrakromoszómális rekombináció. A rekombináció során a tulajdonságokat kialakító genetikai tényezők újrarendeződnek, így új génkombinációk a szülőktől eltérő fenotípusokat eredményeznek.
Az átkereszteződést követően a homológ kromoszómapárok tagjai szétválnak egymástól és külön utódsejtbe kerülnek, mialatt a testvérkromatidák még együtt maradnak (számfelezés). Véletlenszerű, hogy az egyik ill. másik utódsejtbe a kromoszómapárok apai vagy anyai eredetű kromoszómája fog vándorolni.
- A második osztódási szakaszban a testvérkromatidák válnak szét egymástól, így az utódsejtek kromoszómánként egy kromatidából (egy DNS kettős hélixből) álló, de az anyasejthez képest fele kromoszómaszámú kromoszómakészlettel rendelkeznek.
Első főszakasz
Profázis I:
- A homológ kromoszómák hosszuk mentén párt alkotnak, így egy komplexet 4 db. kromatida alkot.
- Átkereszteződések jelennek meg a kromatidák között. A törések következtében a homológok egyik kromatidájában kromoszómaszakaszok cserélődhetnek ki.
Metafázis I A homológok az egyenlítői síkban rendeződnek, a homológok centromereihez az ellenkező pólushoz tartozó húzófonalak tapadnak.
Anafázis I A homológ párok tagjai az ellenkező pólusra jutnak (számfelezés!).
Telofázis I Kialakul a sejtmembrán, de DNS-szintézis nincs!
Az első osztódás redukciós, mert a kromoszómák számát a felére, a haploid számra (n) csökkenti az utódsejtekben.
Második főszakasz
- Lényegében egy mitózis.
- A kromoszómák az egyenlítői síkba rendeződnek.
- A kromatidák szétválnak.
- A húzófonalak az ellentétes pólusokra viszik a kromatidákat.
- 4 haploid utódsejt keletkezik, sejtenként 1 kromatidás kromoszómákkal.
Magyarázzon rajzolt ábra segítségével jelátviteli mechanizmust az adrenalin (glikogénbontó enzimre) és a glukokortikoidok (transzkripcióra) gyakorolt hatásának példáján.
Az állandóan változó külső környezetben az állati szervezet homeosztázisát a biológiai szabályozás teszi lehetővé. A szabályozás feltétele az egyes sejtek, szövetek, szervek működésének összehangolása. A soksejtű szervezetben a homeosztázis érdekében a sejtek között szoros együttműködés van. Valamennyi sejtnek értesülnie kell a többi sejt állapotáról, valamint a saját működésének változásairól. Ez úgy valósul meg, hogy a soksejtűekben a sejtek között egy olyan kommunikációs rendszer létezik, ami a sejtek működésének az összehangolását, így az egész szervezet normális működését szolgálja. A sejtek közötti kommunikáció az egyes sejtek által kapott és adott jelek sorozata, amely meghatározza a sejtek helyét és feladatát. A különféle feladatokat ellátó egységek összehangolását tehát a köztük működő kommunikáció teszi lehetővé, melynek során egyes sejtek különféle jeleket - kémiai, elektromos - bocsátanak ki, amelyeket más sejtek - a célsejtek - felismernek, aminek következtében működésüket megváltoztatják.
A jel megjelenése meghatározó jelentőségű, de a szabályozás szempontjából ugyanilyen jelentősége van a jel megszűnésének is. Ezért a jelmolekulák élettartama meglehetősen rövid (percek, másodpercek).
A jelforgalmi hálózatok működhetnek
- a sejtek között, sejten kívül,
- de egyaránt lehetnek sejten belül is.
A sejtek közötti kommunikációban a jelként ható vegyületek lehetnek pl. hormonok, szöveti hormonok, sejtfelszíni molekulák.
A kommunikációnak, a jelátvitelnek a szervezet szintjén több formája alakult ki, pl. az endokrin kommunikáció a hormonális szabályozás alapja, aminek során belső elválasztású mirigyek hatóanyagaikat, a hormonokat a vérbe ürítik, amely elszállítja azokat a célsejtekhez, megváltoztatva működésüket.
A jelfelfogó sejt a célsejt, mely a jeleket specifikus receptoraival köti meg. A receptor jelenléte teszi szelektívvé a hatást a különféle sejtek között.
A hormonreceptorok elhelyezkedésük szerint lehetnek:
- sejten belüliek (szteroidhormonok),
- sejten kívüliek (pl. peptidhormonok).
A receptorok elhelyezkedése meghatározza a sejten belüli jelátvitel formáját.
Sejten belüli receptorok
A szteroidhormonok – pl. ivari hormonok (tesztoszteron) vagy a mellékvesekéreg hormonjai (glükokortikoidok pl. kortizol) - receptorai a citoplazmában találhatók.
Példaként a kortizol akadálytalanul jut át a sejthártyán, bejutva a sejtbe sejten belüli citoplazmatikus receptorhoz kapcsolódik (GR). A kötődést követően a receptor-hormon komplex transzkripciós faktorként a DNS-hez kötődik, aminek következtében egyes gének aktiválódnak, megindul a transzkripció, majd a fehérjeszintézis, és így létrejönnek azok a fehérjék, melyek a hormonhatás kivitelezéséért felelősek.
A transzkripciós faktorok olyan molekulák, melyek génműködést aktiválnak vagy gátolnak az átírásra – transzkripcióra - gyakorolt hatásuk révén.
https://www.youtube.com/watch?v=m9jOXiYdMeY
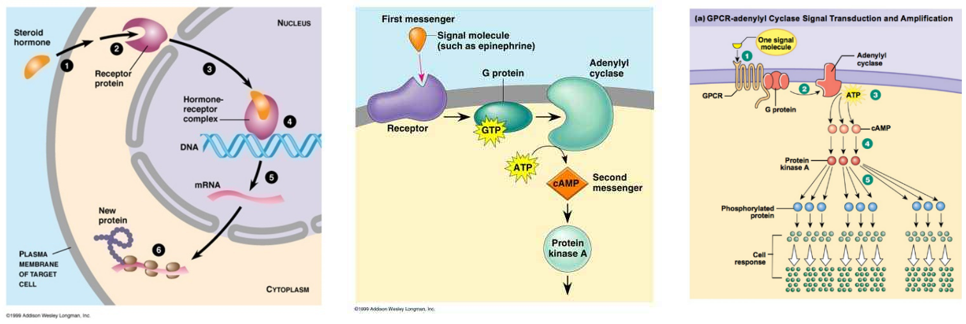 |
A sejten kívüli receptorok a sejthártya felszínén helyezkednek el. Mivel a jel – a hormon, az elsődleges hírvivő - nem jut be a sejt belsejébe - ahol azok az anyagcsere-folyamatok zajlanak, amelyekre hat - szükség van egy olyan sejten belüli jelre, ún. másodlagos hírvivőre, amely a membrán felszínén történő változásokat közvetíti a sejt belseje felé (jelátvitel). Ilyen másodlagos hírvivő pl. a cAMP, vagy a Ca-ion. A másodlagos hírvivők a sejten belül különféle enzimekre – kinázokra - hatnak – serkentik, ill. gátolják azokat -, amivel megváltoztatják a sejtek működését.
A cAMP keletkezése és működése a májsejtekben a következő:
- az adrenalin hormon megkötődik a májsejtek felszínén található receptorokon,
- a receptor konformációt vált, amelynek hatására a mellette levő G-fehérje GTP-t köt meg.
- Ennek köszönhetően a G-fehérje aktiválódik, aminek következménye az adenilát-cikláz aktiválódása.
- Az adenilát-cikláz egy enzim, amely az ATP-ből cAMP-t készít.
- A cAMP protein kinázokat aktivál, ezek olyan enzimek, amelyek foszfátcsoportokat kapcsolnak – foszforilálnak - más enzimekhez azokat működő állapotba hozva.
- Jelen esetben ezek olyan enzimek, melyek a glikogént glükózzá bontják.
Összefoglalva, adrenalin hatására a májsejtekben a glikogén glükózzá bomlik le, amelyet a sejtek a vérpályába adnak le növelve a vércukorszintet.
A jelátvitelt követően jelerősítés történik, aminek köszönhetően néhány jelátvivő molekula is jelentős változást válthat ki a sejt működésében.
https://www.youtube.com/watch?v=rbu-x5-F57w
3.1. Nem sejtes rendszerek |
Magyarázza ábra alapján a vírusfertőzés folyamatát a bakteriofágok litikus és lizogén ciklusa, valamint egy retrovírus példáján keresztül.
A vírusok szaporodása („életmódja”), a vírusfertőzés folyamata
A vírusok nem tekinthetőek sem élőnek, sem élettelennek, az élő és az élettelen rendszerek határán állnak.
A vírusnak két megjelenési formája ismert:
- virion a sejten kívüli fázis, amely élettelen, kristályos szerkezetű.
- A sejten belüli vegetatív fázis, amely a működő, szaporodó vírus.
A vírusok önmagukban semmilyen életjelenséget nem mutatnak, nincs anyagcseréjük. A vírusok kizárólag azáltal képesek szaporodni, hogy megtámadnak valamilyen sejtet, behatolnak, átszervezik a sejt működését, arra kényszerítik, hogy a saját anyagait felhasználva, a bejutott vírushoz hasonló vírusokat hozzon létre. A folyamat a sejt pusztulásához vezet. Ez a jelenség a parazitizmus vagy élősködés.
A vírus által megtámadott sejtet gazdasejtnek nevezzük.
A vírusok többsége gazdaspecifikus, azaz kizárólag a rá jellemző gazdaszervezetben képes szaporodni. Kivételek is ismertek, ilyen pl. a veszettség, száj és körömfájás, tehénhimlő stb. vírusa.
A vírus tehát
- nem élő, mert a gazdasejt nélkül nem képes szaporodni, nincs önálló anyagcseréje.
- De nem is élettelen, mivel a gazdasejtbe jutva, annak anyagait felhasználva szaporodni képes, megváltoztatja a gazdasejt működését.
A fertőzés folyamata, bakteriofágok litikus és lizogén ciklusa
- A virion a közeg áramlásával jut el a gazdasejthez, mivel önálló mozgásra képtelen.
- A virion megtapad a gazdasejt felületén. A megtapadás specifikus víruskötő helyeken (receptorokon) történik. A receptorok mindig az adott sejttípusra jellemzőek, így ezért gazdaspecifikusak a vírusok.
- Bakteriofágok esetén a megtapadás fehérjefonalakkal történik, majd a talpon található enzim (lizozim) kilyukasztja a sejtfalat. A lyukon keresztül a DNS a sejtbe injektálódik, a burok kívül marad.
A sejten belül a vírusok kétféleképpen viselkedhetnek:
- Litikus életciklus
A bejutás után azonnal megkezdődik az új vírusok képzése. A sejt elpusztul, szétszakad.
- Lizogén életciklus
A vírus DNS beépül a gazdasejt DNS-ébe. Ilyenek pl. az RNS tumorvírusok, herpesz vírusok, a HPV, HIV. A beépülés után a vírus sokáig észrevétlen maradhat, majd még nem tisztázott okok miatt – pl. immunrendszer legyengülése - aktívvá válhat.
https://www.youtube.com/watch?v=hFwA0aBX5bE
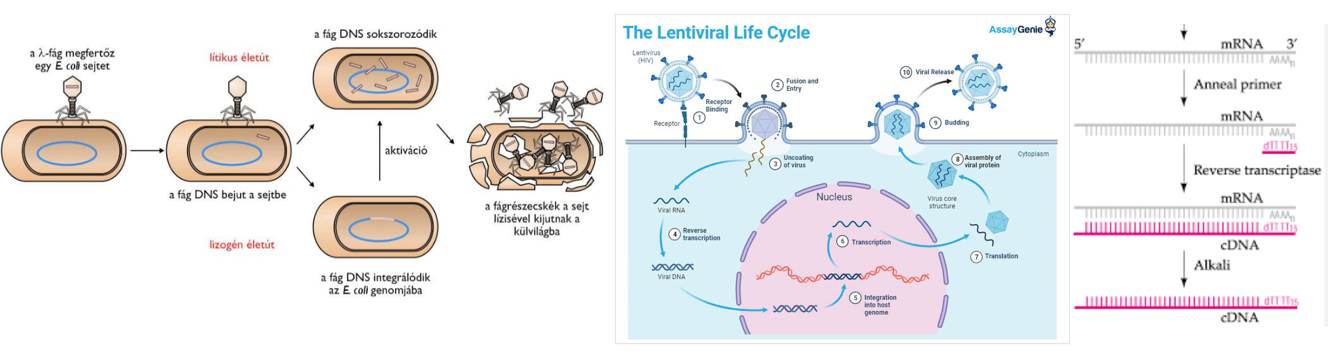 |
A retrovírusok egy speciális víruscsoportot alkotnak, amelyek RNS-alapú genetikai anyagot tartalmaznak, és különleges módon fertőzik meg a gazdasejtet. A legismertebb retrovírus talán a HIV (humán immundeficiencia vírus), ill. ilyenek a koronavírusok és az influenzavírusok.
A HIV fertőzés folyamata
Megkötődés a gazdasejthez
A retrovírus felszínén lévő fehérjék (pl. gp120 a HIV esetén) felismerik és megkötik a gazdasejt membránján lévő specifikus receptorokat (pl. CD4 receptor). Ez a kölcsönhatás lehetővé teszi a vírus bejutását a sejtbe.
Bejutás a sejtbe
A vírus burka összeolvad a gazdasejt membránjával (fúzió), és a vírus belsejében lévő RNS-genom, valamint az enzimek (például reverz transzkriptáz, integráz, proteáz) bekerülnek a sejt citoplazmájába.
Reverz transzkripció (visszairás)
Ez a retrovírusok egyik legfontosabb és megkülönböztető folyamata.
A reverz transzkriptáz enzim a vírus RNS-ét kettős szálú DNS-sé írja át.
Integráció a gazdasejt genomjába
A keletkezett vírus DNS-t az integráz enzim beépíti a gazdasejt sejtmagjában lévő DNS-be. Ettől kezdve a vírus genetikai anyaga a gazdasejt génállományának részévé válik, és együtt replikálódik a sejt DNS-ével.
Transzkripció és transzláció
A beépült vírus DNS-ről a gazdasejt átírja a vírus-RNS-t (mRNS), majd ez alapján vírusfehérjéket szintetizál (transzláció). Ez magában foglalja a vírus burkát alkotó fehérjéket, enzimeket.
Vírus összeállítása és kijutása
Az új vírusalkotó részek (RNS, fehérjék) a sejt citoplazmájában összeállnak. Az új vírusok a gazdasejt membránján keresztül távoznak (bimbózással), miközben megszerzik a vírusburokot a sejtmembrán egy részéből.
https://drive.google.com/file/d/1V7xtd6MtVYo0ZI3TSRUC4oU_v3NoXyzu/view?usp=sharing
Magyarázza, hogy a szivacsos agyvelőgyulladást okozó fehérje normális térszerkezetű változata az egészséges agyszövetben is megtalálható, értelmezzen erre vonatkozó leírásokat, esettanulmányokat.
A prionok (PrPSc) olyan fertőző, kórokozó fehérjék, melyek különféle neurodegeneratív betegségek, mint pl. emberben a Jakob-Creutzfeldt betegség (fertőző szivacsos agyvelősorvadás), szarvasmarhákban a szivacsos agyvelősorvadás (kergemarhakór, szivacsos agyvelőgyulladás) kialakulásában játszanak szerepet. A kórfolyamat előrehaladtával a központi idegrendszer sejtjei elhalnak, az agyban üregek jönnek létre, állománya szivacsossá válik.
A prion betegségre jellemző, hogy nincs ellenanyag termelés, nincs gyulladásos reakció, a lappangási idő évekig tarthat.
A prionoknak nincs örökítő anyaguk, semmilyen életjelenséget nem mutatnak, a vírusoknál is egyszerűbb képződmények. A prionok az egyébként normálisan is meglevő sejtes fehérjék – celluláris prion (PrPc) – szerkezetileg módosult változatai.
A celluláris prionok (PrPc) általában idegsejtek felszínén a membránban találhatók, az idegsejtek egészséges működésében játszanak szerepet. A normál sejtes prionok vízoldékonyak, érzékenyek lebontó enzimekkel szemben.
A kóros prionok (PrPSc) aminosavsorrendje nem, de a másodlagos szerkezete – konformációja - eltér a normálisan is meglevő sejtes fehérjétől, s emiatt bontó enzimekkel szemben nem érzékenyek, vízben pedig nem oldódnak. Feltételezik, hogy a prionok (PrPSc) a sejtekbe jutva elősegítik – katalizálják - a normális fehérjék prionná átalakulását, melyek felhalmozódva, a hidrofób felszínükkel összecsapódva, oldhatatlan rögöket képeznek és az idegsejtek elhalását okozzák.
Esettanulmány az új variáns Creutzfeldt–Jakob-kórról (vCJD), amely jól bemutatja, hogyan történt a fertőzés, hogyan zajlott le a betegség lefolyása, és milyen következtetéseket vontak le belőle az orvosi és közegészségügyi szakemberek.
Egy 19 éves brit nő esete – vCJD diagnózis (1996)
Háttér
- A beteg egy 19 éves nő volt az Egyesült Királyságban.
- Nem volt kórtörténetében korábbi neurológiai vagy pszichiátriai betegség.
- Nem utazott olyan országokba, ahol más típusú prionbetegség gyakori lett volna.
Tünetek megjelenése és kórlefolyás
- Első tünetek (pszichiátriai jellegűek):
- Szorongás, depresszió, ingerlékenység.
- Alvászavarok és szociális visszahúzódás.
- Később neurológiai tünetek:
- Mozgáskoordinációs problémák (ataxia),
- beszédzavarok,
- memóriazavar és súlyos kognitív leépülés.
- A beteg állapota rohamosan romlott.
- 8 hónappal az első tünetek után elhunyt.
Diagnózis és vizsgálatok
- A korai tünetek miatt először depressziót és pszichózist feltételeztek.
- MR-vizsgálat során a pulvinar jel (thalamusban észlelt eltérés) gyanút keltett prionbetegségre.
- EEG: nem volt klasszikus CJD-re jellemző görbe.
- Liquorvizsgálat: a fehérje jelenléte nem volt meggyőző.
- Halál után elvégzett agyszövettani vizsgálat egyértelműen igazolta a PrPˢᶜ jelenlétét és a jellegzetes prionlerakódásokat.
Fertőzés forrásának vizsgálata
- A nő gyermekkorától kezdve az Egyesült Királyságban élt.
- Korábban rendszeresen fogyasztott marhahúst, főként gyorséttermekben.
- A vizsgálatok alapján nagy valószínűséggel BSE-vel fertőzött hús fogyasztása vezetett a fertőzéshez.
Tudományos következtetések
- Ez volt az egyik első bizonyított emberi eset, amely megerősítette:
a BSE átjuthat fajhatáron, és emberi prionbetegséget okozhat. - A beteg agyában és nyirokszöveteiben (pl. mandulákban) megtalált prionok azonos típusúak voltak, mint a BSE-ben – ezt molekuláris priontípus-vizsgálatokkal igazolták.
- Az eset hozzájárult a széles körű közegészségügyi beavatkozásokhoz, pl. állati eredetű fehérje takarmányozásának betiltása, húsipari szabályozások szigorítása.
Következmény
Ez az eset kulcsfontosságú volt a vCJD mint új betegségforma azonosításában, és hozzájárult a BSE emberre való átvihetőségének felismeréséhez, amely világméretű figyelmeztetésként szolgált a prionbetegségek kezelésére, kutatására és megelőzésére.
3.2. Sejtes rendszerek |
Elemezze ábra vagy szöveg alapján a nitrifikáló baktériumok, a denitrifikáló baktériumok, a tejsavbaktériumok és a nitrogénkötő baktériumok anyagátalakítási és energianyerési lépéseit, valamint ezek kapcsolatát.
A nitrifikáló, denitrifikáló, tejsavbaktériumok, valamint a nitrogénkötő baktériumok anyagátalakítási és energianyerési folyamatai, valamint ezek ökológiai kapcsolatai a nitrogénkörforgás és más biogeokémiai ciklusok szempontjából.
Nitrogénkötő baktériumok (nitrogénfixálók)
Anyagátalakítás
- Többnyire heterotróf szervezetek.
- A légköri nitrogént (N₂) alakítják át ammóniává (NH₃). Ez az átalakítás az oxigénre érzékeny nitrogénáz enzim segítségével történik.
- N2 + 8H+ + 8e− + 16ATP → 2NH3 + H2 + 16ADP + 16Pi
Energianyerés
- A nitrogénkötés energiaigényes folyamat (sok ATP kell).
- Az energiát aerob légzéssel, illetve anaerob erjedéssel nyerik.
- Példák:
- Szabadon élő baktériumok: Azotobacter, Clostridium
- Szimbionta baktériumok: Rhizobium (pillangós virágúak gyökerében)
Nitrifikáló baktériumok
Anyagátalakítás
- Kemoautotrófok (kemoszintetizálók).
- A talaj ammóniáját (NH₃/NH₄⁺) oxidálják nitritté (NO₂⁻), majd nitrátokká (NO₃⁻).
- Két lépcsőben zajlik (két baktériumtípus):
- Nitrosomonas: NH3 + O2 → NO2− + H2O + energia
- Nitrobacter: NO2−+ O2 → NO3− + energia
Energianyerés
- Kemoszintetizálók: a fenti oxidációs reakciók során felszabaduló energiát használják szén-dioxid megkötésére, saját szerves anyag előállítására.
- Tehát autotróf szervezetek, de nem fotoszintetizálnak.
Denitrifikáló baktériumok
Anyagátalakítás
- Heterotrófok.
- A nitrátot (NO₃⁻) alakítják vissza nitrogéngázzá (N₂)
- NO3− → NO2− → N2
- Ez a folyamat anaerob környezetben megy végbe (pl. vízzel telített talajokban).
Energianyerés
- Nitrátlégzők, a nitrátot használják terminális elektronfelvevőként (mint ahogy az oxigént az aerob légzésben).
- NO3− → NO2− → NO→ N2O → N2
- Tehát anaerob denitrifikáló légzést végeznek, ezzel energiát nyernek miközben csökkentik a talaj nitráttartalmát.
- Pl.: Nitrococcus denitrificans
Tejsavbaktériumok (lebontók)
Anyagátalakítás
- Heterotrófok, pl. cukrokat (pl. glükóz) bontják le tejsavvá:
- C6H12O6→2CH3CH(OH)COOH + energia
Energianyerés
- Anaerob erjedés révén nyernek energiát (kevés ATP, kb. 2 mol/glükóz).
- Nincs nagy jelentőségük a nitrogénkörforgásban közvetlenül, de szerepük lehet a szerves anyagok lebontásában, pl. a szerves vegyületek nitrogéntartalmának ammóniává történő visszaalakításában.
- Hasznosak élelmiszeriparban (pl. joghurt, savanyúságok).
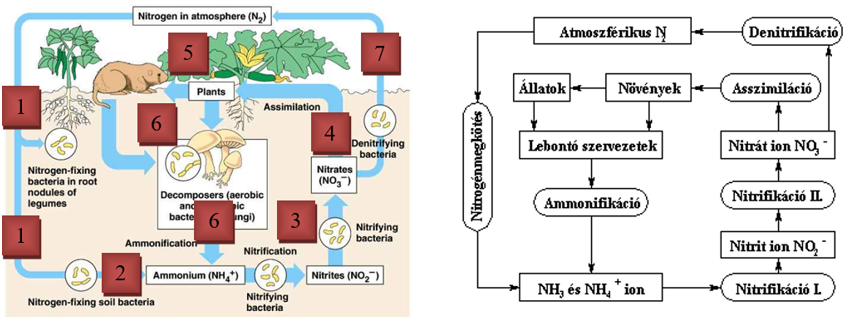 |
A nitrogénkörforgalom összefoglalása
A nitrogén nélkülözhetetlen alkotóeleme a fehérjéknek, nukleinsavaknak. Az elemi nitrogént kizárólag baktériumok és egyes gombák képesek közvetlenül a levegőből felhasználni. Az összes többi élőlény ezek tevékenységére van utalva.
1. A főleg pillangósvirágú növények gyökérgümőiben – Rhizobium - szimbiózisban, és a talajban szaprofita módon élő aerob heterotróf nitrogénkötő baktériumok a légköri nitrogént megkötik és ammóniává alakítják. A folyamat energiaigényes.
2. Az előállított ammónia
- szerves vegyületekbe – aminosavakba – épül be, melyet a gazdanövény részben felhasznál,
- másrészt a talajba kerül (2).
3. Az ammóniát kemoautotróf nitrifikáló baktériumok nitritionná, majd a nitritionokat nitrátionokká oxidálják. (3).
4. Az ammónium- és a nitrátion vízben oldódik, és így a növények könnyen felveszik, s a nitrogént beépítik szerves vegyületekbe, pl. aminosavakba (4).
5. A heterotróf szervezetek a növények által előállított szerves nitrogénvegyületek – pl. fehérjék - elfogyasztásával jutnak nitrogénhez (5).
6. Az elpusztult élőlények nitrogéntartalmú szerves anyagaiból a lebontó baktériumok és gombák ammóniát állítanak elő, amelyből ismét ammóniumion és nitrátion keletkezhet (6).
7. Oxigénmentes talajokban az anaerob vagy fakultatív anaerob denitrifikáló baktériumok hatására a nitrátionokból elemi nitrogén képződik, amely elszökik a levegőbe (nitrát-légzők). A denitrifikáló bacik többnyire anaerob heterotrófok, de ismertek kemoautotrófok is. A denitrifikáló baktériumok jelenléte káros a talaj nitrogéntartalmának szempontjából ezért a talaj szellőztetésével – kapálás – lehet ellenük védekezni (7).
Elemezzen az egysejtűek életmódjával összefüggő kísérleteket.
Ismerje fel ezeket az élőlényeket és jellemző sejtalkotóikat fénymikroszkópos képeken, rajzolt ábrákon.
Az egysejtű élőlények életmódját kutató kísérletek célja gyakran annak vizsgálata, hogy ezek a szervezetek hogyan reagálnak a környezeti ingerekre, milyen anyagcsere-folyamatokat végeznek, és hogyan táplálkoznak, mozognak vagy szaporodnak.
Fototaxis vizsgálata zöld szemesostorosnál (Euglena)
Kísérlet leírása
- Zöld szemesostorosokat tartalmazó vízcseppet világítunk meg csak az egyik oldalról.
- A sejtek a megvilágított terület felé mozognak (pozitív fototaxis).
Értelmezés
- Az Euglena autotróf (fotoszintetizáló) életmódot is folytat, ezért a fény fontos energiaforrás számára.
- A fototaxis annak jele, hogy az egysejtű felismeri és kihasználja a környezeti feltételeket a fotoszintézishez.
- A sejt szemfoltja érzékeli a fényt, az ostor segítségével irányított mozgás valósul meg.
https://www.youtube.com/watch?v=Md0PtdRxXvw
Ozmózisvizsgálat amőbánál (Amoeba proteus)
Kísérlet leírása
- Az amőbát különböző ozmolalitású oldatokba helyezzük (pl. desztillált víz, fiziológiás sóoldat, tömény cukoroldat).
- Megfigyelhető, hogy a lüktetőűröcske működésének gyakorisága a környezet ozmotikus koncentrációjának csökkenésével egyre nő.
Értelmezés
- Az édesvízi egysejtűeknél, mivel a citoplazma koncentrációja meghaladja a környezeti közeg – édesvíz – töménységét, folyamatosan víz áramlik a sejt belsejébe, amit onnan a sejt optimális működésének az érdekében el kell távolítani, ki kell választani. A jelenség hátterében az ozmózis áll. Egysejtűekben a felesleges víz kiválasztását hártyával határolt kis hólyagocskák az ún. lüktető űröcskék végzik. Működésük során a citoplazmából a felesleges vízzel és a benne oldott anyagokkal fokozatosan telítődnek, majd tartalmukat exocitózissal a környezetbe ürítik. A folyamatot fénymikroszkópban szemlélve a telítődés, kiürülés periódusai a lüktetés benyomását keltik.
- Az amőba ozmoregulációt végez, lüktetőűröcske segítségével eltávolítja a felesleges vizet.
- Ez létfontosságú az édesvízi életmódhoz, ahol a sejten belüli és kívüli koncentráció eltér.
- A vizsgálat rámutat arra, hogy még az egysejtűeknek is vannak homeosztatikus szabályozó mechanizmusaik.
https://www.youtube.com/watch?v=o89zBS9fC_c
 |
Papucsállatka táplálkozásának megfigyelése
Kísérlet menete
Szerezzünk be élő papucsállatkákat (akár szénaáztatásos módszerrel).
Készítsünk színesített táplálékoldatot, fessük meg (pl. metilénkékkel, kárminvörössel vagy más festékkel színezve), így a táplálékszemcsék jobban láthatók lesznek a sejtben.
Egy csepp papucsállatka-tenyészetet és egy csepp festett táplálékoldatot helyezünk tárgylemezre, lefedjük fedőlemezzel. A sejtek mozgásának korlátozására célszerű a tárgylemezre vattaszálakat helyezni, hogy a csillósok ne ússzanak ki a látótérből.
Megfigyelés
- Mikroszkóppal figyeljük meg, ahogy a papucsállatkák csillóikkal mozogva kavarják a vizet.
- A sejtszáj környékén lévő csillók mozgatásának hatására a táplálékszemcsék bejutnak a sejtszájba, onnan a sejtgaratba. A kialakuló emésztő űröcskék a plazmaáramlás révén körpályát írnak le a sejtben.
- Tojásfehérje-oldatot színezzük meg kongóvörössel, forraljuk az oldatot 2 percig, majd hagyjuk lehűlni.
- Figyeljük meg a keletkezett emésztő űröcskék színét a mikroszkóp alatt!
A kongóvörös indikátorfesték, amely
- semleges (pH=7) közegben vörös;
- gyengén savas (pH= 5) vörösesibolya;
- erősen savas közegben (pH=3) pedig kék színűvé válik.
Értelmezés
- A papucsállatka többnyire szerves anyagokat bontó baktériumokkal táplálkozik.
- A táplálékfelvétel - szemben az amőbával - sejtfelszín egy meghatározott pontján következhet csak be. A sejt közepén található egy tölcsérszerű bemélyedés, amit szájmezőnek nevezünk. A tölcsér alját sejtszájnak nevezzük. A sejtszájban történik a táplálék bekebelezése, melynek során kialakul egy sejthártyával körülvett hólyag az ún. fagoszóma (Phagia=evés, szoma=test lat.). A folyamatot bekebelezésnek, fagocitózisnak nevezzük.
- A fagoszómával a táplálékot lebontó emésztőenzimeket tartalmazó, hártyával körülvett hólyagocskák, ún. lizoszómák egyesülnek. A fagoszóma és a lizoszóma egybeolvadásával létrejött hólyagocskát emésztő űröcskének nevezzük. Az emésztő űröcskében történik a táplálék megölése, majd lebontása. A citoplazmában, egy időben sok emésztő űröcske található - ezek adják főleg a sejtplazma szemcsézettségét -, amelyek határozott irányú mozgást végeznek.
- A kísérlet során megfigyelhető, hogy a lefűződő emésztő űröcskék színe változik. Ennek oka, hogy az emésztő űröcskében lévő tápanyagtartalom sok enzim közreműködésével bomlik le, és az egyes enzimek különböző pH-tartalmú közegben működnek optimálisan.
- Az emésztés befejezésével az emésztő űröcskék a sejthártyához sodródnak és a lebonthatatlan salakanyagot a felvétellel ellentétes folyamattal – exocitózissal - leadják a környezetbe.
- Ahogy a táplálék felvételének, úgy a leadásának is meghatározott helye van, amit alrésnek nevezünk. Az emésztőűröcskék működése lényegében megegyezik az amőbánál leírtakéval.
https://www.youtube.com/watch?v=sn3MTYNe8mM
3.4. Szövetek, szervek, szervrendszerek, testtájak |
Vizsgáljon fénymikroszkóppal növényi szövet- preparátumot (hajszálgyökér, lágy szár, levél keresztmetszet), készítsen bőrszövet-nyúzatot (pl. hagyma allevél).
Vizsgáljon kristályzárványt. Értelmezze a látottakat, mikroszkópos képen is.
Magyarázza a fás szár kialakulását, az évgyűrűk keletkezését fatörzs keresztmetszetén.
Vizsgáljon mikroszkópban gázcserenyílást és értelmezze a látottakat, mikroszkópos képek alapján is.
Ismerje fel fénymikroszkópos képeken a növényi szöveteket hajszálgyökerek kereszt- és hosszmetszetén, egy- és kétszikű lágyszárú növények szárkeresztmetszetén, kétszikű fás szár keresztmetszetén, valamint kétszikű levél metszetén és hozza összefüggésbe a szerkezeti elemeket azok funkciójával.
Nyúzat készítés
Ilyenkor az epidermiszt csipesszel letépjük az alatta levő szövetekről, ami azért lehetséges, mert az epidermiszsejtek egymáshoz jóval erősebben kapcsolódnak, mint az alatta elhelyezkedő sejtekhez.
https://www.youtube.com/watch?v=eWcheUXSqMA
Kristályzárvány vizsgálata vöröshagymában
Vizsgálat
Vöröshagyma hártyás allevelének darabkáját tegye tárgylemezre, cseppentse le
vízzel, fedje le, és vizsgálja mikroszkópban!
Tapasztalatok és magyarázat
A kristályok anyaga kalciumoxalát. A zárványok vízben nem oldódó anyagcsere végtermékek, amelyeket a sejtek ilyen formában közömbösítenek, vagy tartalék tápanyagok, mint a burgonyagumó keményítőzárványai.
https://www.youtube.com/watch?v=Ev309y40HAU
 |
Hajszálgyökerek kereszt- és hosszmetszete
A hajszálgyökér keresztmetszete
Három szövettájat különböztethetünk meg: a csúcs mögött néhány centiméterrel, a szállítózóna alsó határán.
- Bőrszövetrendszert gyökérszőreivel.
- Az elsődleges kérget, melyet raktározó alapszövet épít fel.
- A központi hengert, amelyben az egyszerű szállítónyalábok helyezkednek el.
A gyökér bőrszövetén nincsenek gázcserenyílások és nincsen kutikularéteg sem. A gyökér bőrszövetének zárvatermőkre jellemző speciális képződményei a gyökérszőrök. A gyökérszőrök a gyökér bőrszövet sejtjeinek kesztyűujjszerű kitűrődései. A gyökérszőrökön keresztül történik a víz és az oldott ionok felvétele. A gyökérszőrök feladata a felületnagyobbítás. A gyökérszőrök élettartama néhány nap, a csúcstól távolabb pusztulnak, a csúcs irányába mindig megújulnak.
A hajszálgyökér hosszmetszete
A gyökéren függőlegesen különböző működési zónák vannak.
1. Osztódási zóna
A gyökér csúcsi része osztódószövetből áll, amit kívülről a gyökérsüveg véd, melynek sejtjei elnyálkásodva segítik a gyökér mozgását a talajban, továbbá kőzetet oldó gyökérsavat termelhetnek.
2. Megnyúlási zóna
Itt a legerőteljesebb a sejtek hosszirányú növekedése (auxin hatására).
3. Felszívási zóna
Gyökérszőrök jellemzik. Legfelső sejtjei fokozatosan elhalnak, míg a csúcshoz közel újraképződnek, így biztosítva a folytonos tápanyagfelvételt.
4. Szállítási zóna
A felszívott anyagok a gyökér egyszerű szállítónyalábjaiban továbbítódnak a szár felé.
 |
Kétszikű lágy szárú növények szárkeresztmetszete
A lágy szár, akárcsak a gyökér, keresztmetszetét vizsgálva 3 szövettájra tagolódik.
1. Bőrszövet
- Egyrétegű epidermis.
2. Elsődleges kéreg
- Az elsődleges kéreg raktározó, ill. táplálékkészítő alapszövetet tartalmaz. A fiatal szárak többnyire zöldek, a táplálékkészítő alapszövetnek köszönhetően képesek fotoszintetizálni.
3. Központi henger
- A központi henger alapszövetbe ágyazódva tartalmazza a szállítónyalábokat.
A lágy szárban a szállítónyalábok elrendeződése lehet
- körkörös, mint a kétszikűekben vagy
- szórt, mint az egyszikűekben.
Általános tendencia, hogy a faelemek a szár közepe felé, a háncselemek perifériásan helyezkednek el.
A fás szár szerkezete
A fás szár a több éves zárvatermőkre, ill. a nyitvatermőkre jellemző. Kívülről befelé haladva a következő rétegeket lehet megkülönböztetni.
- Többrétegű bőrszövet, a periderma, héjkéreg.
- Élő háncs, általában néhány mm.
- Kambiumgyűrű.
- Fatest,
- melynek külső, vizet és sókat szállító élőrésze a szijács.
- Az elhalt fatest a geszt, amely elsősorban szilárdító feladatokat lát el. Az idősebb szállítóelemek elzáródnak, a sejtek falába tartósító, ún. cseranyagok rakódnak, amelyek védik az elhalt elemeket a lebontó szervezetektől.
Ahol az év során nem kiegyenlítettek a csapadékviszonyok (mérsékelt égöv), ott a fák kambiuma
- elsősorban a csapadékosabb tavaszi időszakban működik intenzíven, ekkor nagyobb átmérőjűek a vízszállítóelemek (világosabb tavaszi pászta),
- nyár végén és ősszel szűkebb átmérőjű, vastagabb falú csövek jönnek létre (sötétebb őszi pászta), hiszen a növények már kevesebb vizet vesznek fel,
- ennek köszönhető az évgyűrűs szerkezet a mérsékelt égövi fáknál.
- Télen természetesen nem működik a kambium.
 |
A szár fejlődése
Attól függően, hogy a kambium
- különálló kötegeket képez, akkor lágy szár marad,
- vagy összefüggő hengert alkot, ekkor fás szár alakul ki (azaz a kambium a nyalábok között is kialakulva gyűrűvé záródik).
A fás szárú növények is először lágyszárú csemeteként indulnak fejlődésnek, majd a kambiumgyűrű záródásával alakul ki a fás szár.
Kétszikű levél metszete
A lomblevél felépítése
A lomblevelek a fotoszintézis, a gázcsere és a párologtatás szervei. A lomblevelekre jellemző:
- nagy felület,
- kiterjedt sejtközötti járatrendszer,
- raktározó szövetek nincsenek,
- nincs periderma és másodlagos vastagodás.
Szöveti felépítés
A levelet minden oldalról egyrétegű bőrszövet – epidermis - határolja, melyet a kiszáradás ellen védelmet nyújtó kutikula borít. A bőrszövet színtelen, átlátszó, zöld színtesteket nem tartalmaz (kivéve a sztómák zárósejtjeit).
A bőrszövet folytonosságát a gázcserenyílások (sztómák) szakítják meg, amelyek
- a kétszikűeknél a levelek fonákján,
- míg a víz felszínén úszó leveleknek csak a színén találhatók meg.
- egyszikűeknél mindkét oldalon megtalálhatók.
A sztómák elsősorban a levélen találhatók, de jelen vannak fiatal szárak epidermiszén, virágleveleken, sőt esetleg föld alatti szárakon is, de mindig hiányoznak a gyökereken.
A sztómákon belül megkülönböztetjük
- a légrést és
- a légrést körülvevő bab alakú két zárósejtet.
A sztómákat ún. kísérősejtek veszik körül. A bab alakú zárósejtekben zöld színtest található.
A légrés alatt egy nagyobb üreg, a légudvar helyezkedik el, amely rendkívül gazdag sejtközötti járatrendszerben folytatódik. A gázcserenyílások tehát a sejtközötti járatrendszer felszíni kivezetőnyílásai.
A zárósejtek lehetnek:
- az epidermiszsejtek szintjében, közepes vízellátottságú növényeknél (mezomorf),
- kiemelkedhetnek a bőséges vízellátottságú helyeken (higromorf),
- be is süllyedhetnek a sivatagi, félsivatagi növényeknél (xeromorf).
- A párhuzamos erezetű leveleken (egyszikűek és tűlevelek) a sztómák párhuzamos sorokban,
- a hálózatos erezetű leveleken (kétszikűek) a sztómák szórtan állnak.
A sztómák kettős feladatot látnak el,
- lebonyolítják a gázcserét, illetve
- végzik a párologtatást.
A zárósejtek speciális felépítésük révén a légrést tágítani vagy szűkíteni képesek,
- amennyiben a zárósejtek K+ és vizet vesznek fel és megduzzadnak, turgoruk nő, a légrés kinyílik,
- ellenkező esetben a vízvesztés következtében a zárósejtek összezáródnak.
A bőrszöveten belül található a zöld színtestekben gazdag, fotoszintetizáló táplálékkészítő alapszövet, melyre jellemző a gazdag sejtközötti járatrendszer.
A kétszikű növények leveleiben szerkezete felépülhet
- a felületre merőleges irányban megnyúló sejtekből álló, ún. oszlopos alapszövetből
- és laza szivacsos alapszövetből.
Az oszlopos réteg rendesen a levél felső oldalán helyezkedik el, a levél alsó része szivacsos alapszövetből áll. Az ilyen, kétszikűekre jellemző leveleken csak az alsó epidermiszen vannak sztómák. Az oszlopos alapszövet elsősorban a fotoszintézis helye, a szivacsos réteg pedig elsősorban a párologtatás színtere.
Az egyszikűekben csak szivacsos alapszövet fordul elő, így a levél mindkét felszínén megtalálhatók sztómák.
A levél alapszövetében futnak a szállítónyalábok, melyek a levél erezetét alkotják.
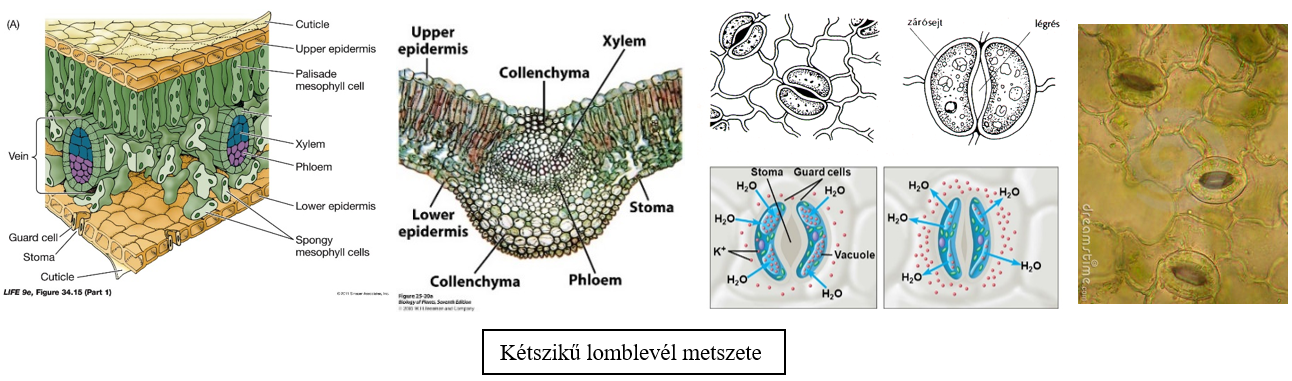 |
A levél erezete
A levél erezetét a levéllemezben futó zárt szállítónyalábok képezik. Az erekben a farész a levél színe felé, a háncsrész a levél fonákja felé néz. A tápanyagszállítás mellett, mivel szilárdító alapszöveteket is tartalmaznak, merevítik is a levelet, kifeszítve tartják a levéllemezt, s védik a szél tépő hatásával szemben.
Az erezet formái:
- Hálózatos, főérből és belőle elágazó oldalerekből áll, kétszikűekben.
- Párhuzamos, egyforma nagyságú mellékerekből áll, egyszikűekben.
Végezzen el növényi anyagszállítással kapcsolatos kísérletet, magyarázza a tapasztaltakat.
Értelmezzen növényi anyagszállítással kapcsolatos kísérletet.
A víz útja a zárvatermő növényben
Vizsgálat
Állítsunk fehér virágú növényt tintával megszínezett vízbe! Vizsgáljuk meg a szár keresztmetszetét különböző magasságokban. Vizsgáljuk meg a virágot is.
Tapasztalat és magyarázat
A tinta a szállítószövetrendszer farészét festette meg, ugyanis ez szállítja a gyökér felől a vizet és az ionokat a levelek irányába. A sziromlevelekben a festék a levélerekben jelent meg, hiszen azok a szállítószövet elemei a lomblevélben.
https://www.youtube.com/watch?v=RMT_njY1UDw
Magyarázza a csírázás külső és belső feltételeit egy csírázási kísérlet kapcsán.
A magvas növények élete különböző fejlődési szakaszokra osztható fel.
- Csíra- és magképzés (lásd fent),
- nyugalmi állapot,
- csírázás,
- vegetatív fejlődés, a hajtás kialakulása,
- reproduktív fejlődés, a virág és a termés kialakulása, ivarsejtek képzése.
A nyugalmi állapot
A megtermékenyítést követően létrejövő mag, benne a kialakult embrióval, nyugalmi állapotba kerül.
A nyugalmi állapot kialakulását külső és belső feltételek biztosítják.
- Külső gátló tényező lehet, ha nem megfelelőek a csírázás környezeti feltételei pl. hőmérséklet.
- A belső korlátozó feltételeket különféle csírázást gátló anyagok – pl. abszcizinsav - felszabadulása jelenti, ugyanis a legtöbb magnak létrejötte után még utóérésre van szüksége.
A nyugalmi állapot kialakulása során:
- a mag víztartalma csökken,
- az enzimek inaktiválódnak,
- csírázást gátló anyagok szabadulnak fel, pl. az abszcizinsav, amelyek általában a termésből kerülnek a magba (dinnyemagvak kiszedve rögtön csíráznak, dinnyelébe áztatva nem).
A csírázás
Amikor a csírázást gátló anyagok lebomlanak, a csírázás véget vet a magvak nyugalmi állapotának. A csírázás során az embrió a mag tartalék anyagainak felhasználásával kinő a magból és megkezdi vegetatív fejlődését.
A csírázás során
- a mag vizet vesz fel (duzzadás),
- enzimjei aktiválódnak,
- megkezdi a tartaléktápanyagok lebontását, átalakítását,
- megindul a sejtek osztódása,
- a növény áttöri a maghéjat és intenzív növekedésnek indul.
A csírázás külső feltételei
A víz
A magok vízfelvételét duzzadásnak nevezzük, mivel a folyamat közben a mag térfogata megnő - akár duplájára is - és a maghéj felreped. Az inaktív, légszáraz magvak vízfelvétellel aktivizálják enzimjeiket.
Az oxigén
Az oxigén a tartaléktápanyagok biológiai oxidációval történő lebontásához szükséges. A biológiai oxidáció csak a maghéj felrepedése után indulhat meg, ugyanis a maghéj meggátolja az oxigén felvételét, ezért addig a tápanyagok erjedéssel bomlanak le.
A hőmérséklet
A csírázás minimum pontja 0, maximum pontja 50 fok körül van. Az egyes növények származási helyüknek megfelelő hőmérsékletet igényelnek csírázásukhoz.
A csírázást a növényi fejlődés hőszakaszának nevezzük.
A fény
A legtöbb esetben a fény közömbös a csírázásra nézve, ugyanakkor ismertek fényben, ill. sötétben csírázók, ahol a fény szükséges, ill. gátló hatású a csírázásra.
Belső feltételek
Életképes embrió
A magban található embriónak életképesnek kell lennie.
Megfelelő érettség
A csírázást gátló anyagoknak – pl. abszcizinsav - le kell bomlaniuk.
Tartalék tápanyagok
A mag belsejében található tápanyagok (pl. keményítő, fehérjék, olajok) lebomlása biztosítja az energiát az első növekedési szakaszhoz.
Csírázási kísérlet bemutatása
Vizsgáljuk a víz, az oxigén és a hőmérséklet hatását a csírázásra.
Anyagok
Babmag, vatta/papírtörlő, víz, edények, hűtőszekrény, lezárt üveg.
Kezelési csoportok
- 1. Nedves vattára helyezett mag szobahőmérsékleten → csírázik.
- 2. Száraz vattán tartott mag → nem csírázik (nincs víz).
- 3. Nedves vattán, de lezárt edényben → nem csírázik (nincs oxigén).
- 4. Nedves vattán, de hűtőben → nem vagy lassan csírázik (alacsony hőmérséklet).
https://www.youtube.com/watch?v=evVXN1TMKj4&t=199s
Értelmezzen auxin hormonokkal végzett kísérletet.
Auxin hormonnal végzett kísérletek, kísérletek pázsitfüvek koleoptiljával
A koleoptil egy védőburok, amely a pázsitfüvek csírázásakor a hajtáskezdeményt védi, sejtjei megnyúlásával növekedik.
Darwin és fia kísérletei
- Megfigyelték, hogy a zab csíranövény koleoptilja egyoldali megvilágítás hatására a fény felé görbül.
- Amennyiben a hajtáscsúcsot eltávolították, az elhajlás elmaradt.
- Észlelték, hogy a hajtás csúcsát eltakarva átlátszatlan fóliával, szintén elmarad a görbülés (növekedés egyébként ekkor is van), noha a növekvő részt éri a fény.
- A növekvő részt eltakarva és a hajtáscsúcsot szabadon hagyva görbülést tapasztaltak.
A kísérletek alapján levonható következtetések:
- a növekedési hormon a koleoptil csúcsban képződik.
- A csúcsban képződő hormon gyökér irányába szállítódik és kiváltja a hajtás egyenlőtlen növekedését.
Peter Boysen-Jensen (dán) kísérletei
- Ha levágta a hajtáscsúcsot, majd azt agar-agar zselatin kocka közbeiktatásával helyezte vissza, a növény elhajlott a fény felé, ezzel bizonyította, hogy a jel át tud jutni a levágott és visszahelyezett csúcs alatti agardarabon.
- Ha vajat tett a csúcs alá, a görbülés elmaradt, amiből az következik, hogy az anyag, ami a csúcsból vándorol lefelé, vízoldékony természetű.
 |
Paál Árpád kísérletei
- Paál Árpád sötétben végezte a kísérleteit a koleoptillal.
- A levágott csúcsot aszimmetrikusan tette vissza a koleoptilra és az ellenkező oldalra görbülést tapasztalt.
Következtetése:
- a csúcsban termelődik a növekedésszabályozó anyag,
- lefelé vándorol, és azon az oldalon, amelyiken levándorol, serkenti a növekedést, ezért
- a növény a másik oldalra görbült, mint amelyikre a hajtáscsúcsot ragasztotta.
Frits Went (holland) kísérletei
- A levágott csúcsokat agarkockákra tette
- A tiszta agar nem okozott sem növekedést, sem görbülést, de amelyik érintkezett a hajtáscsúcsokkal, igen.
- Középre visszahelyezve egyenes, míg aszimmetrikus pozícióban görbülő növekedést tapasztalt,
- a kockák növekedésserkentő hatása, az elhajlás szöge egy határon belül arányos avval az idővel, ameddig a hajtáscsúcsot a kockán tartotta.
- Went adta az auxin nevet.
Következtetések
- A növekedést serkentő hormon bekerült az agar-agar kockákba.
- A növekedést serkentő hatás bizonyos határok között egyenesen arányos a hormon koncentrációjával.
- A hormont tartalmazó agar-agar kocka féloldalas elhelyezése az egyoldalú megvilágításhoz hasonló eredményt hozott.
A fenti jelenség az auxin hormon hatásával magyarázhatók. Összefoglalva az auxin
- hormon a növekedéshez szükséges,
- a sejtek megnyúlását serkenti, hatására a cellulóz molekulák közötti kötések lazulnak,
- továbbá fokozza a sejtosztódást, a sejtlégzést, a sejtek K+ felvételét, így a víz felvételét is.
- A hajtástenyésző csúcsokban termelődik,
- kis mennyiségben fiatal rügyekben, levelekben, éretlen termésekben, magvakban is termelődik,
- A megvilágítással ellentétes oldalon – az árnyékos részen - vándorol lefelé a háncsrészben,
- a fény felőli oldalon az auxin szállítás gátlódik.
- A fény egyenlőtlen hormoneloszlást idéz elő, ez váltja ki az egyenlőtlen növekedést és ezáltal a görbülést. Az egyenlőtlen hormoneloszlás mesterségesen is előidézhető és ekkor sötétben is görbülés következik be.
Ismerje fel fénymikroszkópos készítményen, illetve képeken a következő szöveteket: többrétegű elszarusodott laphám, vázizom szövet, csontszövet, idegszövet, emberi vér.
Ismerje fel rajz alapján vagy mikroszkópos képek alapján a következő szöveteket: simaizom szövet, szívizom szövet, csillós hám, üvegporc. Értelmezze a látott struktúrák szerepét a szövet működésében.
A többrétegű hámszövetek olyan felületeken fordulnak elő, amelyek fokozott mértékben vannak kitéve mechanikai, ill. kémiai behatásoknak. A felülethez közel eső hámsejtek aránylag távol esnek a hámszövetet tápláló vérerektől, ezért gyakran a hám és a kötőszövet érintkező felülete megnövekszik, hullámos, ún. kötőszöveti szemölcsök alakulnak ki.
Többrétegű elszarusodó laphám
E hám legfelső sejtsorai elhalnak, elszarusodnak, aminek következtében egy szaruréteg alakul ki a felszínen, amely véd a kiszáradástól és a mechanikai hatásoktól. Emiatt jellemzően a szárazföldi gerincesek – hüllők, madarak, emlősök - bőrének felszínén fordul elő.
A felületen elpusztuló sejtek pótlása a szövet legalsó sejtsorában mitótikus osztódással történik. A sejtek, miközben felfelé tolódnak, szarusodási folyamaton mennek keresztül, a folyamat egyes lépéseinek megfelelően, különböző rétegek alakulnak ki.
A hám legalsó sejtsora hengerhámsejtekből áll, melyek folyamatosan osztódnak, ezért e réteget osztódó rétegnek nevezzük. E rétegben barna pigmentet, melanint tartalmazó nyúlványos pigmentsejtek vagy melanociták is előfordulhatnak.
Legfölül az elhalt szaruréteg található. Az eltávozó anyagok miatt az elszarusodó sejtek zsugorodnak, lapossá válnak. A szarupikkelyekké átalakult laphámsejtekben már sem a sejtmag, sem a sejtalkotók nem ismerhetők fel. A szaruréteg területén a sejtek már szinte csak keratin fehérjét tartalmaznak.
Az egyrétegű csillós hengerhám
Hosszúkás, hatszögletű sejtekből épül fel, sejtjei csillókat viselnek a felületükön. A csillók mozgékony, apró sejtnyúlványok.
E hámféleség megtalálható:
- az orrüreg belső felszínén,
- a légcsőben,
- a petevezetékben.
A porcszövet (üvegporc)
A porcszövet sejtekből és sejtközötti állományból áll. A porcsejtek egyesével vagy csoportosan helyezkednek el az alapállományban szigetszerűen, kisebb-nagyobb távolságra egymástól. A porcsejtek termelik a sejtközötti állományt.
A sejtközötti állomány
- fehérje-szénhidrát jellegű ragasztóanyagból (chondroitin-szulfátból),
- és kötőszöveti rostokból áll.
A porcot - az ízületi porc kivételével - erekben, idegekben gazdag porchártya borítja. Mivel vérerek hiányoznak a porcszövetből, így táplálása diffúzió útján történik a porchártya felől. A porcszövet tápanyag- és oxigénellátottsága gyenge, ezért anyagcseréje lassú, sérülés esetén regenerációra nem képes.
Idősebb szervezetben, az egyébként rugalmasan szilárd porc amúgy is rossz táplálékellátottsága a víztartalom csökkenése miatt tovább romlik, a porcsejtek egy része elpusztul, s az alapállományba mészsók rakódhatnak bele, a porc rideggé, törékennyé válik.
Porcszövet építi fel többek között
- a légutak porcait,
- az ízületi porcokat.
A lemezes csontszövet
Az állati szervezet legkeményebb szövete, a csontos halaktól kezdve minden gerinces állat belső vázának az alapanyaga. A csontszövetet
összetétel szerint:
- szerves és
- szervetlen állományra,
felépítés szerint:
- sejtekre és
- sejtközötti állományra oszthatjuk.
A szerves állományt:
- csontsejtek és
- kollagénrostok alkotják.
A szervetlen állományt a sejtközötti állomány szervetlen sói alkotják:
- Ca5(PO4)3 X, ahol X= F, OH (apatitok),
- CaCO3.
A szerves állomány a csontok rugalmasságát, a szervetlen állomány a csontok szilárdságát biztosítja.
- A szervetlen állományt sósavas kezeléssel eltávolíthatjuk, ekkor a csontok gumiszerűen hajlíthatók lesznek.
- A szerves állományt égetéssel távolíthatjuk el, ekkor a csontok elvesztve rugalmasságukat ridegek és törékenyek lesznek.
A csontszövet koncentrikus felépítésű, ún. oszteonokból épül fel. Egy oszteon kb. 10 mm magas henger, melynek közepén a centrális csatornában vérér fut. Az oszteonban a nyúlványos csontsejtek összekapcsolódva koncentrikus körökbe rendeződnek.
A sejtek között az alapállomány szintén körkörösen rendeződött lemezei futnak, amelyet szervetlen sók, kollagénrostok és szerves kötőanyag épít fel.
A csontszövet vérerekkel átszőtt, élénk anyagcseréjű szövet.
A csontszövet felületét az ún. csonthártya borítja, amely sok rostot tartalmazó kötőszövetes lemez. A csonthártyában futnak a csontszövetet ellátó erek és idegek.
A csonthártya belső rétegében csontképző sejtek helyezkednek el, innen indul a sérült csont regenerációja, ill. növekedés során a csontok vastagodása.
A csontszövet az egész élet folyamán mindig az aktuális terhelésnek megfelelően épül fel, azaz, ha pl. öregedéskor a test súlypontja eltolódik, azt a csont belső szerkezetének megváltozása követi. A folyamat során ún. csontfaló sejtek lebontják a csontszövetet, a csontképző sejtek pedig az új terhelés irányában építik át.
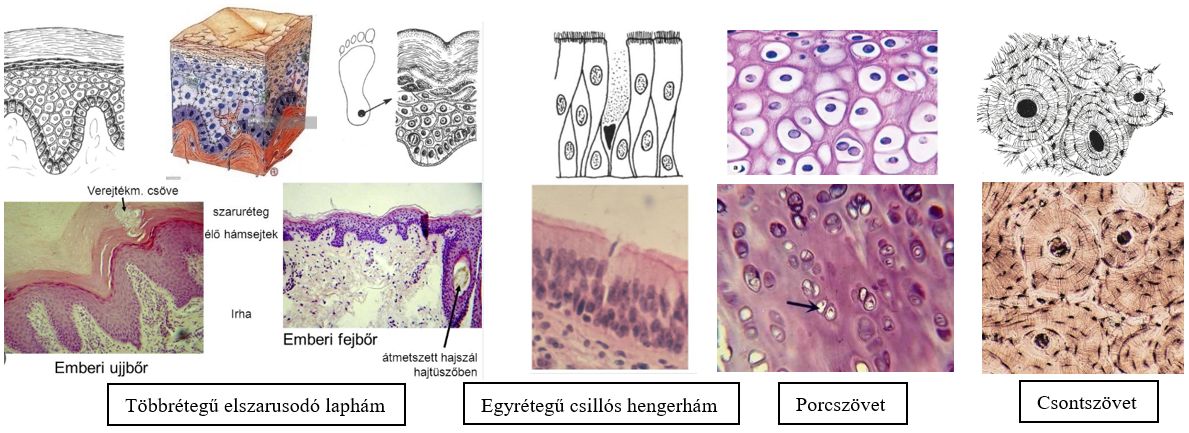 |
Az izomszövetek típusai
- harántcsíkolt izomszövet, amely lényegében a vázizomszövet,
- szívizomszövet,
- simaizomszövet.
Harántcsíkolt izomszövet, vázizomszövet
Megtalálható:
- a gerincesek és az ízeltlábúak vázát mozgató izmokban,
- a nyelvben, garatban, nyelőcső felső szakaszában, végbélnyílásban.
- Ilyenek a légzőizmok.
Jellemzői:
- általában akaratunktól függően működik,
- gyors,
- nagy erőkifejtésre képes,
- fáradékony.
A harántcsíkolt izomszövet szöveti egysége a sokmagvú izomrost. A sejtmagvak száma egy rostban meghaladhatja a 7OOO-t.
- Sok izomrost,
- köteggé – izomnyalábbá - szerveződik, amelyet kötőszövetes hártya tart össze,
- az izomrostkötegek tovább rendeződve alkotják az izmokat, amelyeket szintén egy kötőszövetes lemez, az ún. izompólya határol.
Az izomroston belül továbbá megkülönböztetünk
- miofibrillum-kötegeket, amelyek
- izomfonalakból (miofilamentumok) állnak.
Tehát az izomrostok citoplazmájában a hossztengellyel párhuzamosan húzódnak az összehúzódásért felelős miofibrillumok, amelyek már fénymikroszkóppal is láthatók.
A miofibrillumok között nagy mennyiségű mitokondrium helyezkedik el, melyek biztosítják az izom-összehúzódáshoz szükséges energiát.
A Ca-raktárként funkcionáló SER harisnyaszerűen veszi körbe az egyes miofibrillumokat.
A nagyszámú sejtmag perifériásan, az izomrost membránja alatt található.
A miofibrillumok felépítése és az izom-összehúzódás
A vázizomzatban az izomrostok harántirányban - a hossztengelyre merőlegesen - csíkozottságot mutatnak, azaz világosabb és sötétebb sávok váltják egymást.
A harántcsíkolat látszatát az okozza, hogy a hosszanti szerkezeti elemek, a miofibrillumok, szakaszosan váltakozva más-más felépítésűek, s az egyforma szakaszok azonos hosszúságúak és egy magasságban helyezkednek el.
A miofibrillumok izomfonalakból épülnek fel, ezek azonban már csak elektronmikroszkópban láthatók.
Kétféle izomfonál különböztethető meg:
- a vékony fonál, világos, aktin fehérjéből épül fel,
- a vastag fonál, sötét, miozin fehérje alkotja.
Az izom-összehúzódás röviden (részletesen lásd 4.3. fejezet)
Egy elernyedt, nyugalomban lévő izomban a vékony fonalak csak kis mértékben nyúlnak be a vastag fonalak közé, a vékony és a vastag fonalak közötti kapcsolódások száma csekély (izomtónus).
Az izom-összehúzódás során, a vékony fonalak becsúsznak a vastag fonalak közé, aminek köszönhetően az izomrostok megrövidülnek. Az izom-összehúzódás mindig az izom hossztengelyével párhuzamosan zajlik.
Lépések röviden
- Az izomrostokat beidegző idegrostok végződéseiből felszabaduló ingerületátvivő anyagok hatására az izomrostok citoplazmájában Ca2+-ok szabadulnak fel (SER-ből), aminek hatására a miozinmolekulák feji részei hozzákötődnek az aktinfonalakhoz.
- A kötődést követően a miozin feji részei bólintó mozgást végeznek (konformációváltozás), aminek köszönhetően a vékony fonalak beljebb csúsznak a vastag fonalak közé, egységnyi távolságot megtéve.
- Ezután az ATP és Mg2+ megkötésekor a miozinfejek leválnak az aktinfonalakról.
- ATP bontásakor felszabaduló energia hatására a miozin fejek kiegyenesednek, majd már egy másik kötőhelyen ismét hozzákötődnek a vékony fonalakhoz és kezdődik minden elölről.
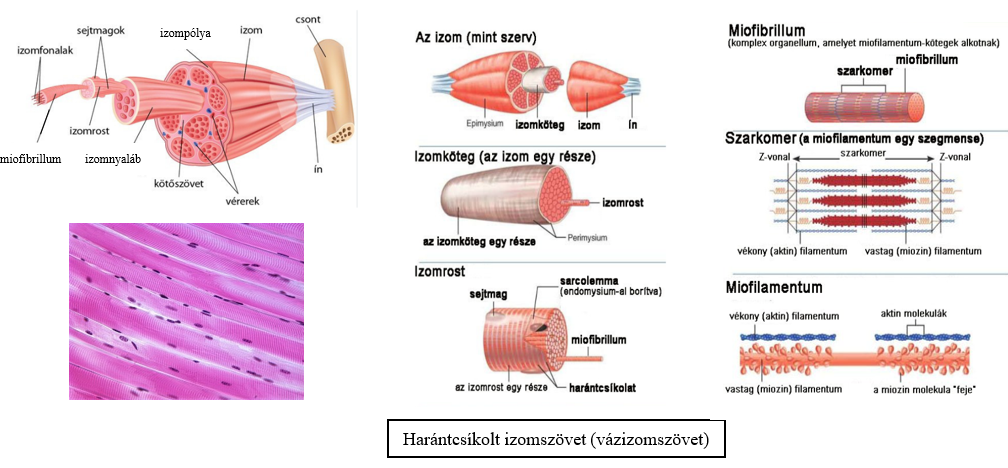 |
A szívizomszövet
A szívizomszövet, mely a harántcsíkolt izomszövetnek egy speciális félesége, a szív falának középső rétegében található meg.
Jellemzői:
- akaratunktól függetlenül működik,
- gyors,
- nagy erőkifejtésre képes,
- nem fáradékony.
Alaktanilag két sajátságban különbözik a vázizomszövettől:
- egymagvú izomsejtekből áll, melyek elnyújtott térrácsot alkotnak, a sejtek helyenként elágaznak,
- a sejtmagvak a sejt közepén helyezkednek el.
A simaizomszövet
A simaizomszövet elnyúlt orsó alakú sejtekből épül fel. A sejtmag a sejt közepén helyezkedik el. A sejtekben megtalálható az aktin és a miozin, de a harántcsíkolt izomszövetre jellemző nagyfokú rendezettség nem jellemző.
Előfordulása:
- a gerincesek zsigeri izomzata: erek falában, bélcső falában, légcső falában stb.,
- bőrben,
- egyes gerinctelenek bőrizomtömlőjében.
Tulajdonságai:
- akaratunktól függetlenül működik,
- lassú,
- kis erőkifejtésre képes,
- nem fáradékony.
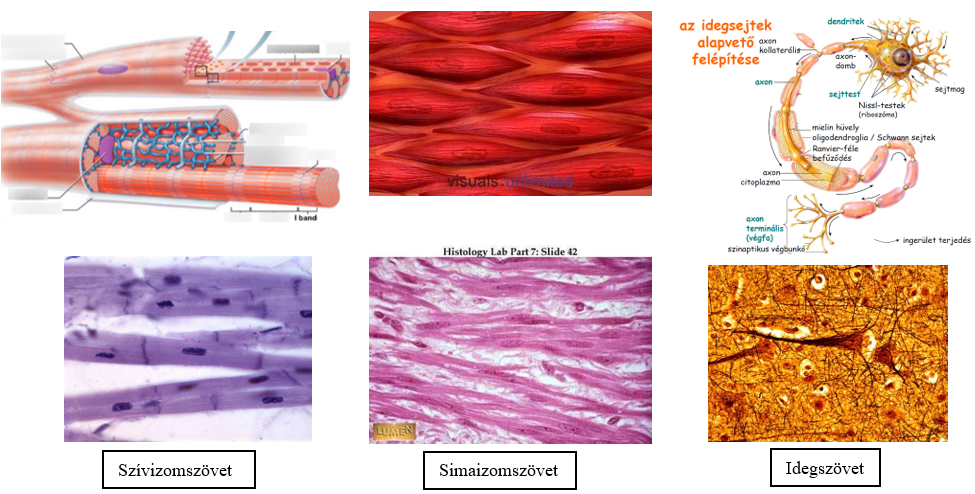 |
Az idegszövet
Az idegszövet tartalmaz:
- nyúlványos idegsejteket, ún. neuronokat,
- gliasejteket.
A neuronok (lásd még 4.8.1. jegyzet)
A neuronok ingerlékeny sejtek, amelyek ingerfelvételre és idegi ingerületek vezetésére specializálódtak. Egymagvú, nyúlványos sejtek, számuk emberben hozzávetőleg 100 milliárd.
A neuronok részei:
- sejttest, amely a sejtmagot, ill. a sejtszervecskék zömét tartalmazza,
- dendritek, rövidebb, elágazó nyúlványok, amelyek nagy számban fordulhatnak elő,
- axon, vagy tengelyfonal, a hosszú, ingerület vezetésére specializálódott főnyúlvány, amely gyakran elágazik.
A sejttest
Tartalmaz:
- sejtmagot,
- mitokondriumokat, melyek kielégítik a neuronok nagy energiaigényét,
- RER-t,
- Golgi-készüléket.
A fenti alkotórészekből következően, igen intenzív fehérjeképzés folyik az idegsejtekben.
Nincs azonban sejtközpont, tehát osztódni nem képesek. Az idegsejtek végleges száma már a születés környékén kialakul.
A dendritek
Felépítésük megegyezik a sejttestével, ezért plazmanyúlványoknak is tekinthetjük őket.
Rendkívül elágazók. Felületükön kiemelkedések, ún. tövisek lehetnek. Feladatuk a sejt felszínének a növelése, az idegsejtek közötti kapcsolódásokban – a szinapszisokban - játszanak szerepet.
Az axon (neurit, tengelyfonal)
Sejtenként max. egy fordul elő. Hossza emberben elérheti az 1 m-t (piramissejtek). Az axon eredését a sejttesten axondombnak, végelágazódását végfácskának (végbunkó) nevezzük. A végfácska rendkívül elágazó, akár több száz végződésből is állhat, amelyek a sejtek közötti kapcsolódásban - szinapszis - rendkívüli jelentőségűek.
Az axonok oldalelágazásokat képezhetnek, aminek köszönhetően egy idegsejt több másik idegsejttel is kapcsolatot létesíthet.
Egyes gliasejtek az axonok köré tekeredve ún. velőshüvelyt hoznak létre. A velőshüvelyes axon az idegrost.
Neuroglia jellemzői, funkciói
Az idegrendszer „kötő- és támasztószövete”, a gliasejtek biztosítják a neuronok működéséhez szükséges feltételeket.
- Számuk kb. 10 x-es mennyiségben haladja meg az idegsejtekét.
- Életünk végéig osztódóképesek maradnak.
- Biztosítják az idegszövet kémiai védelmét, létrehozzák az agy-vér gátat.
- Biztosítják az idegszövet homeosztázisát,
- táplálják a neuronokat.
- Velőshüvely képzése révén elektromos szigetelést biztosítanak az axonok körül, gyorsítják az ingerületvezetést.
- Degenerált neuronokat eltávolítják, majd kitöltik a megüresedett tereket.
Elemezze leírt vizsgálatok/kísérletek alapján a felsorolt magatartásformákat: öröklött
magatartásforma, tanult magatartásforma, önfenntartó viselkedés, fajfenntartó viselkedés, taxis, öröklött mozgásmintázat, kulcsinger, motiváció, feltétlen reflex, bevésődés, érzékenyítés, megszokás, feltételes reflex, operáns tanulás, belátásos tanulás, önzetlenség, agresszió.
Lásd 3.4.4. Az állatok szövetei, szaporodása, viselkedése
https://bioszfera.com/downloads/3.4.4.Azallatokszovetei-szaporodasa-viselkedese.pdf
4.1. Homeosztázis, rendszerszemlélet |
Magyarázza ábra, szöveges leírás, táblázatban vagy grafikonon megadott adatok alapján a pozitív és negatív visszacsatolás szerepét az élettani folyamatok során.
Az élő rendszerekben végbemenő folyamatok nem véletlenszerű irányban, sebességgel stb. mennek végbe, hanem meghatározott irányítás alatt állnak.
Az irányítás alapműveletei
- Alapjelképzés: valamilyen tényezőnek – pl. testhőmérséklet - tervezett értéket kell adni (kell érték).
- Mérés: ennek a jellemzőnek a tényleges értékét méri a rendszer (van érték).
- Összehasonlítás: a különbség meghatározása a kell és a van érték között (hibajel).
- Döntés: szükség esetén – hibajel - beavatkozás a rendszer működésébe.
Irányítás módszerei a vezérlés és a szabályozás
Vezérlés során a vezérlő az irányított rendszert egyirányúan működteti, az irányított rendszer működése során nem hat a vezérlő központra (egyirányú kapcsolat).
A szabályozás
A szabályozás során az irányított rendszer folyamatosan visszajelez a központnak, befolyásolva annak működését (kétirányú kapcsolat).
Negatív visszacsatolás
A biológiai szabályozás alapja a negatív visszacsatolás. A negatív visszacsatolás az a folyamat, amikor a szabályozórendszer a hibajellel ellentétes előjelű hatással módosítja a rendszer működését (csökkenésre növelés, növekedésre csökkentés). Ilyenkor a rendszerből kilépő hatások csökkentik a folyamatok mértékét.
A negatív visszacsatolás a rendszert stabilizálja. Önszabályozás jön létre, a rendszer egy dinamikus egyensúlyi állapotban tartja fenn önmagát. A sejtek, élő szervezetek optimális működésének előfeltétele egyfajta belső stabilitás, a belső környezet egyfajta állandósága. A belső környezet szabályozott állandóságának az állapotát homeosztázisnak nevezzük. Az önszabályozó folyamatok az élő szervezetek sokféle jellemzőjét - kémhatás, ionkoncentráció, térfogat stb. - tartják állandó értéken, de legalábbis egy meghatározott tartományon belül.
A negatív visszacsatolás szerepe az élettani folyamatok során
Testhőmérséklet szabályozása
- Alacsony környezeti hőmérséklet esetén a testhőmérséklet csökken (36 °C, van érték),
- a központ összehasonlítja a kell (37 °C) és a van értéket, létrejön a hibajel,
- a fűtőközpont aktivitása fokozódik, a testhőmérséklet emelkedik, ami csökkenti a fűtőközpont aktivitását.
Hormonális szabályozás
- A hipotalamusz TRH-t (Thyroidea Releasing Hormon), az agyalapi mirigy pedig TSH-t (pajzismirigy-stimuláló hormon) bocsát ki.
- A TSH és a TRH hatására a pajzsmirigy több tiroxint termel.
- Ha a tiroxinszint elér egy adott értéket, gátolja a TRH és TSH termelődését → ezek elválasztása csökken → a tiroxin termelés csökken.
Vízháztartás szabályozása
- A víz visszaszívódását elősegítő hormon, az ADH (antidiuretikus hormon, vazopresszin).
- A hipotalamuszban termelődik és az agyalapi mirigy hátsó lebenyében raktározódik.
- Vízhiányos állapotban – amikor a vér ozmotikus koncentrációja (ozmózisnyomása) megnő, fokozódik az ADH felszabadulása,
- ennek hatására a gyűjtőcsövek fala áteresztővé válik a víz számára, így az passzív transzporttal a csatornákat körülvevő nagyobb koncentrációjú szövetnedvbe, ill. a vérplazmába áramlik.
- Ennek eredményeképpen csökken a vér ozmotikus koncentrációja,
- emiatt pedig csökken az ADH elválasztása.
Vérnyomásszabályozás, az aortaív reflex
- Inger: vérnyomás emelkedés.
- Receptor az aortaív falában,
- depressor kp. aktiválás, pressor kp. gátlás,
- az érfalak tónusának csökkenése, értágulás, vérnyomás csökken,
- X. paraszimpatikus agyideg közvetlenül gátolja a szinuszcsomót,
-
- szívfrekvencia csökken,
- perctérfogat csökken.
Vércukorszint szabályozása
- Ha a vércukorszint megemelkedik (pl. evés után), a hasnyálmirigy β-sejtjei inzulint termelnek.
- Az inzulin segíti a glükóz sejtekbe való bejutását → a vércukorszint csökken.
- A csökkenő vércukorszint miatt csökken az inzulin elválasztása.
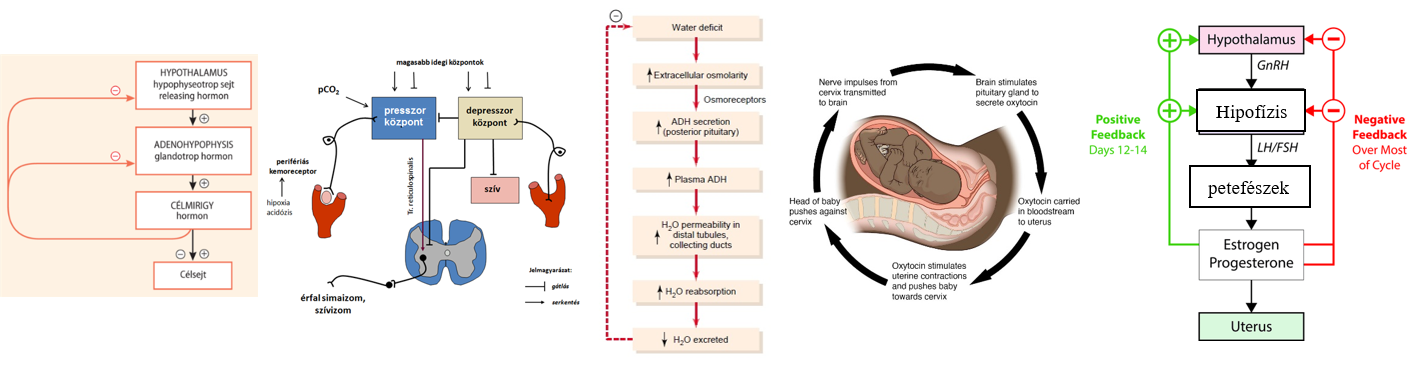 |
Pozitív visszacsatolás
Pozitív visszacsatolásról akkor beszélünk, ha a rendszerből kilépő hatások tovább erősítik a folyamatok mértékét, (növekedésre növelés, csökkenésre csökkentés).
A pozitív visszacsatolás (pozitív feedback) az élettani folyamatokban egy olyan szabályozási mechanizmus, amely során a válaszreakció erősíti a kiváltó inger hatását. Ez egy önerősítő folyamat, ellentétben a negatív visszacsatolással, amely gátolja vagy csillapítja a kiinduló változást, és ezzel fenntartja a homeosztázist.
A pozitív visszacsatolás szerepe az élettani folyamatok során
A női nemi ciklus szabályozása
- A ciklus elején, a menses első napjaiban a petefészek hormonok – ösztrogén és progeszteron – koncentrációja a vérben alacsony.
- Ennek következtében a hipofízis tüszőserkentő hormonszintje a vérben emelkedik, gyorsul a tüsző érése.
- A fejlődő tüsző termeli az ösztrogént, ami fokozza az FSH, ill. az LH elválasztását (úgy, hogy hatására nő az FSH-t és az LH-t elválasztó sejteken a GnRH receptorok száma).
- Emiatt felgyorsul a tüsző érése, egyre nagyobb mennyiségű ösztrogént termel, ami tovább emeli az FSH és az LH szintjét a vérben.
Véralvadás
- A véralvadás láncreakciója lassan indul, de lépésről-lépésre egyre jobban felgyorsul.
- A gyorsulás egyik oka az egymás után aktivált faktorok egyre növekvő koncentrációja a plazmában, minden egymást követő lépésben egyre több aktív enzim lesz jelen.
- A gyorsulás egy másik oka a kaszkádban később aktiválódó enzimek pozitív visszahatása a megelőző reakciókra.
Szülés (vajúdás) – oxitocin visszacsatolás
- A méh összehúzódásai hatására a magzat feje nyomást gyakorol a méhszájra, méhszáj tágul.
- A növekvő nyomás hatására a hipotalamusz fokozottan termel oxitocint.
- Az oxitocin tovább fokozza a méh összehúzódásait → a méhszáj még jobban tágul → még több oxitocin szabadul fel.
Társas kapcsolatok
- A társak közelsége magasabb oxitocinszinttel jár, ami a kölcsönös társas interakciók – pl. érintés, megkapaszkodás, simogatás – révén mind magasabb oxitocinszintet idéz elő.
- Ennek jótékony hatását érzékelve az egyedek még inkább nyitottabbá válnak a baráti, pozitív társas interakciókra, és így továbbemelkedik az oxitocinszint.
Elemezzen vizsgálatokat a homeosztázis és az egészség kapcsolatára vonatkozóan a hőszabályozás és a cukorbetegség kapcsán.
Hőszabályozás
Az ember állandó testhőmérsékletű; maghőmérséklete 37 oC, ez a végtagokon lehet változó.
A szabályozott folyamatok, melyek az állandó testhőmérsékletet biztosítják:
- hőtermelés,
- hőleadás,
- hőfelvétel, egyensúlyuk biztosítja az optimális állandó testhőmérsékletet, amely a homeosztázis egyik fontos paramétere.
A hőtermelés nyugalomban főleg a zsigerekhez, munkában az izmokhoz kapcsolható.
- Az alaphőtermelést az alapanyagcsere lebontó folyamatai biztosítják, melyek során a szerves vegyületek energiatartalmának csupán 40 %-a konzerválódik ATP formájában, a többi – 60% - hővé alakul. A hőtermelés a testtömeggel arányosan nő.
- Didergéses hőtermelés során, az izmok remegése, fokozott munkája fejleszt további hőt (4x-esen), hiszen az izom összehúzódáshoz szükséges ATP bontásakor felszabaduló energia egy jelentős része szintén hővé alakul.
- Nem didergéses hőtermeléskor egyes hormonok hatására ( tiroxin) fokozódó zsír, glikogén bontásakor felszabaduló hő biztosítja az állandó testhőmérsékletet.
Hőtermelő szervek az izmok, a máj és a vesék.
Hőleadás módjai:
- vezetés – különböző hőmérsékletű érintkező felületek között játszódik le,
- áramlás – amikor a meleg levegő kisebb sűrűségű, ezért felszáll,
- sugárzás – infrasugárzás által,
- párolgás útján, mely különböző mértékben, de állandóan zajlik, a bőr felszínéről a verejték párolgása hőt von el a kitágult bőr ereinek véréből.
A hőszabályozás központja a hipotalamuszban van, ahol két ellentétes hatású mag – fűtőközpont, hűtőközpont - működésének az egyensúlya biztosítja az állandó testhőmérsékletet. A szabályozás a negatív visszacsatolás elvén alapszik.
A fűtőközpont integrálja azokat a mechanizmusokat, amelyek alacsonyabb külső környezeti hőmérséklet esetén is biztosítják az optimális testhőmérsékletet:
- a bőr erei összehúzódnak,
- a lebontó anyagcsere mértéke nő (az adrenalin, tiroxin elválasztás nő),
- didergéses hőtermelés beindul,
- a szőrzetborzolás kialakul, aminek köszönhetően a szőrszálak között egy jól hőszigetelő légréteg alakul ki,
- viselkedésbeli változások tapasztalhatók.
- Összességében szimpatikus reakció.
A hűtőközpont aktivitása során
- a bőr erei tágulnak, a bőr véráramlása nő,
- a verejtékezés fokozódik,
- a légzés fokozódik, miáltal a tüdőn keresztül leadott hőmennyiség nő,
- a lebontó anyagcsere mértéke csökken,
- viselkedésbeli változások következnek be.
- Paraszimpatikus reakció.
Termoreceptorok a belső szervekben (máj), ill. a hipotalamuszban és a bőrben találhatók.
A hőszabályozás negatív visszacsatolás elvén alapul. A negatív visszacsatolás az a folyamat, amikor a szabályozórendszer a hibajellel ellentétes előjelű hatással módosítja a rendszer működését (csökkenésre növelés, növekedésre csökkentés). Ilyenkor a rendszerből kilépő hatások csökkentik a folyamatok mértékét. Pl.:
- Alacsony környezeti hőmérséklet esetén a testhőmérséklet csökken (36 °C, van érték),
- a perifériás és centrális hőérzékelő receptorok a érzékelik hőmérsékleti változást,
- a központ összehasonlítja a kell (37 °C) és a van értéket, létrejön a hibajel,
- a fűtőközpont aktivitása fokozódik, a testhőmérséklet emelkedik, ami csökkenti a fűtőközpont aktivitását.
Láz
Lázról akkor beszélhetünk, ha a testhőmérséklet 38,2 Celsius fok fölé emelkedik. Lázas állapotban a maghőmérséklet azért emelkedik, mert a hipotalamikus kell érték magasabbra állítódik különféle anyagok pl. egyes fehérvérsejtek által termelt vegyületek, továbbá a különféle kórokozók bomlástermékeinek hatására.
A hipotalamusz hőérzékeny neuronjai a normális testhőmérsékletet alacsonynak érzékelik és ennek megfelelően működtetik a lehűlés elleni szabályozást. A láz az immunválasz fontos eleme, mivel magasabb hőmérsékleten a kórokozók szaporodása csökken, az immunrendszer működése fokozódik.
- Első szakasza a didergés. A hőtermelés beindul, bőr erei szűkülnek (hőleadás csökken), így a testhőmérséklet emelkedik.
- A láz állandó szakasza alatt a hipotalamusz neuroncsoportjai ezt az emelkedett maghőmérsékletet tartják stabil szinten, a hőleadás ekkor egyenlő a hőtermeléssel, a bőr meleg, de a verejtékezés még nem indul meg.
- Harmadik szakasz a verejtékezés, mivel a kell érték visszaállítódik, így beindul a hűtés. 1 fokos hőváltozás 500-1000 ml folyadékveszteséget jelent! A testhőmérséklet visszaáll az eredeti értékre.
Lázcsillapítás
A láz bizonyos értéktartományban segíti a gyógyulási folyamatokat, azonban 41 Celsius fok felett életveszélyessé válik, mivel magasabb hőmérsékleten megváltozik az enzimek térszerkezete, aktív centruma, ami az anyagcsere folyamatok leállásához vezet. Ezért a lázat 39 Celsius fok felett csillapítani kell. Erre több lehetőség van.
Gyógyszeresen paracetamol, aspirin, amidazophen, ibuprofen segítségével, melyek gátolják a lázat okozó vegyületek felszabadulását.
Fizikai módszerekkel, pl. hűtőfürdő (újabban ellenjavalt), borogatás alkalmazásával.
Hőszabályozás zavarai
Hipertermia
A hipertermia a testhőmérséklet kóros, szabályozatlan emelkedését jelenti, amely túlhaladja a normális fiziológiás határokat (> 38,5–39 °C, súlyos esetben > 40–41 °C). A hipertermia nem azonos a lázzal (amely a hipotalamusz által szabályozott hőmérséklet-emelkedés), hanem a hőtermelés és hőleadás egyensúlyának felborulásából ered, és súlyos élettani következményekkel járhat.
Hipertermia élettani következményei
- Fejfájás, szédülés
- Zavartság
- Görcsök
- Eszméletvesztés, esetleg kóma
- Súlyos esetben agyödéma és neuronális károsodás
- Perifériás értágulat → vérnyomásesés
- Vérnyomásesés → szervi vérellátási zavar (különösen agy, vese, máj)
- Fehérjék denaturálódnak → enzimek működése leáll
- Sejtpusztulás → szöveti károsodás
- Izzadás → folyadék- és nátriumvesztés, dehidráció
- Akut májkárosodás: májenzimek emelkednek, véralvadási zavar
- Akut vesekárosodás: → veseelégtelenség
Hipotermia
A hipotermia olyan állapot, amikor a test központi hőmérséklete 35 °C alá csökken. Ez a szervezet hőtermelése és hőleadása közötti egyensúly felborulása miatt következik be, és súlyos élettani zavarokat okozhat.
Hiportermia élettani következményei
- A hőmérséklet csökkenésével az idegrendszer működése fokozatosan lelassul:
- Zavartság, memóriaromlás
- Koordinációs zavar, beszédzavar
- Aluszékonyság, eszméletvesztés
- 28 °C alatt gyakori a mély tudatzavar vagy kóma
- Lelassult szívverés
- Alacsony vérnyomás
- 30 °C alatt kamrafibrilláció
- Súlyos esetben: szívmegállás
- Légzésszám csökkenése
- 24–25 °C alatt gyakori a légzésleállás
- Az enzimaktivitás lelassul → anyagcsere lassulása
- A szervezet oxigénfelhasználása és CO₂-termelése is csökken
- Acidózis: a keringési és légzési elégtelenség miatt laktát halmozódhat fel
- Hiperglikémia: a hideg gátolja az inzulin hatását
- Súlyos esetben: akut veseelégtelenség a keringési és metabolikus zavarok miatt
- Paradoxális levetkőzés: súlyos hipotermiánál előfordulhat, hogy az egyén levetkőzik, mivel az agy hőérzékelő rendszere zavart szenved.
Az inzulin
Az inzulin egy 51 aminosavból álló polipeptid. Az inzulin az anyagcserét anabolikus irányba tolja el, a tápanyagok, testépítő vegyületek beépülését, raktározását serkenti.
Szénhidrát-anyagcserére gyakorolt hatás
- Egyetlen hormonként csökkenti a vércukorszintet, mivel segíti a szövetek, sejtek cukor felvételét, segíti a cukor bejutását a sejtekbe, (elsősorban a máj-, zsír-, izomszövetekre hat, az idegsejtek glükóz felvételét nem befolyásolja),
- serkenti a máj glikogén szintézisét, csökkenti a glükóz leadását, gátolja a glükoneogenezist,
- növeli az izom glikogén szintézisének mértékét,
- fokozza a szövetek - főleg a zsírszövet - cukor oxidációját, serkenti az ebből meginduló zsírszintézist, ami biztosítja, hogy a feleslegben felvett szénhidrátok (csoki) zsírok formájában raktározódjanak.
Az inzulin termelődésének a mértékét alapvetően a vércukorszint határozza meg. Táplálkozást követően az emelkedő vércukorszint már néhány perc alatt megnöveli az inzulin elválasztását, melynek mértékét a bélben termelődő bélhormonok jelentősen fokozzák.
A cukorbetegség (diabetes mellitus (mézédes húgyár))
Alapvetően kétféle cukorbetegséget szokás megkülönböztetni:
- 1-es típusú cukorbetegség: a betegség oka a hasnyálmirigy Langerhans-szigeteiben az inzulintermelő béta-sejtek pusztulása. Ez a szigetsejtek elleni autoimmun-reakció következménye. Bármely életkorban kialakulhat. A betegség csak inzulinpótlással kezelhető. Gyermekkori diabetes elsődleges oka.
- 2-es típusú cukorbetegség: a csökkent inzulinérzékenység – ún. inzulinrezisztencia - következtében alakulhat ki. Az inzulinrezisztencia az a jelenség, amikor a sejtek nem érzékelik az inzulin jelenlétét, aminek hátterében az inzulinreceptorok számának csökkenése, ill. szerkezetük megváltozása áll. A betegség kezdetén még vércukoreltérés nem jelentkezik, a hasnyálmirigy fokozott inzulintermelése miatt. A betegség előrehaladtával a hasnyálmirigy inzulin-elválasztása a béta-sejtek kimerülése miatt elégtelenné válik és emiatt kialakulnak az 1-es típusú diabetes tünetei. Kialakulásának hátterében örökletes hajlam, túlsúly, inaktív fizikai életmód, egészségtelen étrend (magas cukor- és szénhidráttartalommal) áll.
- Másodlagos cukorbetegség: kialakulhat alkoholizmus vagy epeúti kövesség következtében.
- A terhességi cukorbetegség: oka, hogy a terhesség alatt a méhlepényben termelődő hormonok csökkenthetik az anyai szervezetben termelődő inzulin hatását.
A diabetesz elsődleges tünetei
- a nagy mennyiségű édes vizelet (mézédes húgyár), tehát cukorürítés,
- a vízvesztés miatt kialakuló szomjúságérzet, sok folyadék fogyasztása,
- acetonos lehelet,
- gyakori eszméletvesztés,
- testsúlycsökkenés stb.
A tünetek hátterében a magas vércukorszint és az ennek ellenére fennálló sejtéhezés áll (mivel a sejtek inzulin hiányában nem képesek felvenni a glükózt (éhezés terített asztalnál)).
Összefoglalva:
Az inzulin elősegíti a glükóz felvételét, lebontását, zsírokká (máj, zsírszövetek) vagy glikogénné (máj, vázizom) alakulását.
Inzulin hiányában
- magas a vércukorszint, a cukor a szövetekből vizet vonva el a vizelettel távozik, (a vizelet mennyisége megnő, mivel a szűrlet koncentrációja a benne maradó cukortól magas, így kevesebb víz tud visszaszívódni, a betegek vízfogyasztása a vízvesztés következtében megnő),
- a sejtek glükóz hiányában fokozott zsírbontásra kapcsolnak, aminek következtében számos a vér pH-ját savas irányba mozdító anyagcseretermék ún. ketontestek halmozódnak fel, mint pl. béta-hidroxi-vajsav, acetecetsav, aceton stb. Ennek eredménye az acidózis, ami eszméletvesztést, súlyosabb esetben kómát okoz.
Hosszútávon a cukorbetegségnek számos szövődménye lehet, aminek hátterében az áll, hogy a magas vércukorszint miatt a szervezet fehérjéihez a normálisnál sokkal több cukor kapcsolódik, ami megváltoztatja a szerkezetüket, rontja a működésüket.
Ezért:
- a fertőzésekkel szemben csökken az ellenállóképesség,
- a sebgyógyulás elhúzódik, sportsérülések valószínűsége megnő,
- érelmeszesedés kockázatának növekedése várható,
- a betegek látása romlik, a retinán vérömlenyek alakulnak ki.
- A vese kisereinek károsodása, mely fehérjevizelést, magas vérnyomást, végül veseelégtelenséget eredményezhet.
- Érzéskiesések, fájdalom, vegetatív működési zavarok alakulnak ki. A vegetatív zavarok közé tartozik az impotencia, hasmenés vagy székrekedés, fokozott izzadás, szapora szívverés.
Az 1-es típusú cukorbetegség mai tudásunk szerint nem előzhető meg, mivel kialakulásában örökletes tényezők döntőek.
A 2-es típusú cukorbetegség kialakulásában örökletes és életmódbeli tényezők is szerepet játszanak.
Életmódbeli tényezők:
- elhízás,
- sok cukor fogyasztása,
- mozgásszegény életmód,
- stressz,
- alkoholizmus.
Kezelés
Az 1-es típusú diabetes kezelése diétával és az inzulin bőr alatti zsírszövetbe juttatásával történik a végtagok, ill. a hasfal területén. Az inzulint be kell fecskendezni, szájon át jelenleg nem adható, mert peptid lévén a gyomorban kicsapódik, ill. lebomlik.
A 2-es típusú cukorbetegségben a betegség kezdetén ajánlott az életmód-változtatás, szénhidrátszegény diétával, testsúlycsökkentéssel, sporttal. Ha ez nem elégséges gyógyszerek segítségével, amelyek csökkentik a vércukorszintet, növelik az inzulin hatását, fokozzák a sejtek inzulinérzékenységét. Ha a béta-sejtek kimerültek, az inzulint teljesen pótolni kell.
4.2. Kültakaró |
Ismerje fel mikroszkópos metszeten és ábrákon a bőr szöveti szerkezetét, ismertesse a részek funkcióit.
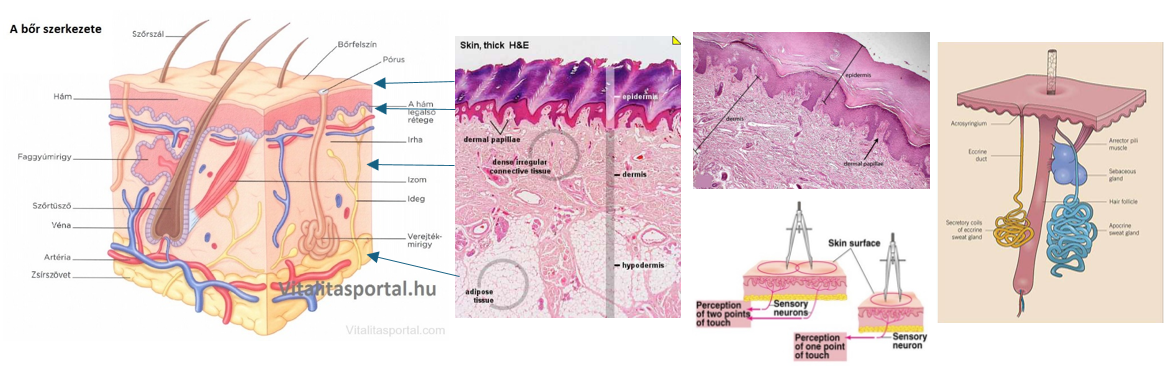 |
Az emberi bőr felépítése és feladatai
Bőrünk
- a szervezetünket elhatárolja a környezettől, első védelmi vonal,
- véd a külső, károsító mechanikai hatásokkal szemben,
- véd a kórokozóktól,
- véd a káros UV-sugárzástól,
- véd a kiszáradástól,
- részt vesz a hőszabályozásban,
- hőszigetelésben,
- kiválasztásban,
- tápanyagraktár,
- D vitamint termel,
- legnagyobb érzékszervünk.
- A szervezetünk egészségi állapotának indikátora.
A gerinces állatokéhoz hasonlóan az emberi bőrt is három réteg építi fel:
- a felhám,
- az irha és a
- bőralja.
Felhám (epidermis)
A bőr legfelső rétege a felhám, szövettanilag többrétegű elszarusodó laphám.
- A sejtek igen szorosan záródnak.
- A hámréteg alsó sejtjei folyamatosan mitózissal osztódnak,
- és az idősebb sejteket fokozatosan a felszín felé tolják,
- a sejtek ellapulnak, szaruanyag (keratin) halmozódik fel bennük,
- végül elpusztulnak, vékony szarupikkellyé alakulnak.
- Egy hámsejt a h ám aljáról mintegy 28 nap alatt jut ki a legkülső hámsorba.
A szaruréteg véd a
- kémiai és mechanikai hatásoktól,
- csökkenti a párologtatást (véd a kiszáradástól),
- akadályozza a kórokozók bejutását a szervezetbe.
- Legvastagabb a talpon, tenyéren, legvékonyabb a szemhéjon.
A bőr színét a felhám alsó részén elhelyezkedő pigmentsejtek adják. A bennük termelődő sötétszínű melanin elnyeli a nap ultraibolya sugárzását, így védi a mélyebb rétegeket. UV hatására a festékanyag termelése fokozódik, és a bőr sötétebbé válik. A nagy intenzitású UV károsítja a sejteket, mert megváltoztatja a DNS-molekulák szerkezetét, mutációt idéz elő (bőrrák). A melanin lokális felszaporodásával alakulnak ki az anyajegyek, a szeplők, ritkábban a májfoltok.
Az irha (dermis)
- A felhámréteget az alatta található irha rögzíti és táplálja.
- Erekben és idegekben, mirigyekben gazdag lazarostos kötőszövet.
- A nagy érintkezési felület biztosítja a hám hatékonyabb táplálását.
- Kollagén- és rugalmas rostokban
- A felső réteg jellemző mintázata a felelős az ujjakon, a tenyéren és talpon lévő, egyénekre jellemző bőrlécrajzolatért is (ujjlenyomat).
- A gazdag érhálózat a hőszabályozásban is jelentős, pl. melegben az erek kitágulnak, nő a hőleadás (kipirulunk).
A bőr receptorai
Eloszlásuk nem egyenletes, az ujjbegyen és az ajkakon nagy sűrűségűek. A kétpont-küszöb az a legkisebb távolság, melynél két vékony pálca érintését különbözőnek (kettőnek) érezzük. Ez az ujjbegyen a legkisebb, hiszen itt a receptorok sűrűsége a legnagyobb.
Az irha felszíni rétegeiben található receptorok a finomabb tapintást érzékelik.
A bőr mélyebb rétegeiben, a zsírszövetek közelében helyezkednek el az erőteljesebb mechanikai hatásra aktiválódó nyomásérzékeny receptorok.
A szabad idegvégződések magas ingerküszöbű, szövetkárosodás veszélyével járó ingereket fájdalomként érzékelő receptorok, továbbá ezek felelősek a hőmérséklet-változás érzékeléséért.
A bőr mirigyei
Ide tartoznak a faggyúmirigyek, a verejtékmirigyek, az illatmirigyek, és a tejmirigyek. Hámszármazékok.
Faggyúmirigyek
- Külső elválasztású mirigyek.
- A szőrtüszőkhöz csatlakozva váladékukat szőrszálakhoz ürítik.
- Zsíros váladékukkal elősegítik a szőrszálak és a bőr vízállóságát és rugalmasságát,
- puhán tartják a szaruréteget.
- A faggyú rétegében szimbióta baktériumok tapadnak meg, savas váladékot (pH 5.5) termelve védik a bőrt.
Verejtékmirigyek
Hosszú feltekeredett végkamrájú csöves mirigyek az irha aljában találhatók. Két fajtájuk ismert.
A kisebb verejtékmirigyek megtalálhatóak az egész testen.
- Szerepük a hőszabályozásban van, túlhevülés esetén a verejték párolgása a bőr felületéről hőt von el, ezáltal hűti a bőrön átfolyó vért.
- A pangó verejték az elszaporodó baktériumok miatt kellemetlen szagúvá válhat.
- Kiválasztó szerepet is játszanak, a verejték nagyon híg vizeletnek tekinthető (némi túlzással persze).
A nagy verejtékmirigyek (illatmirigyek) kivezetőcsövei a szőrtüszőkbe nyílnak.
- Működésük pubertáskorban indul meg hormonális hatásokra, nemi vonzalomban van szerepük.
- Izgalom hatásra lúgos kémhatású váladék termelődik, amely gyorsan bomlik, ezért könnyen kellemetlen szagú lehet.
Elhelyezkedésük:
- hajlatokban,
- emlők környéke,
- nemi szervek környéke,
- végbélnyílás környéke.
Szőrök és szerepük
- Kizárólag emlősállatokra jellemzők (kivétel pl. cetek).
- A tenyér és a talp kivételével teljes testfelületünket szőrök fedik.
- Mechanikai védelemben,
- hőszigetelésben játszanak szerepet.
- Cserélődésük a vedlés, amely folyamatosan zajlik (a szaruréteggel együtt).
A szőr szerkezete
- Bőrből kiemelkedő része a szőr szára, mely ferdén nő ki a bőrből.
- A bőrben levő része a szőrtüsző, amely a hám csőszerű betüremkedéséből jön létre az irhába.
- A szőrtüszőhöz szőrmerevítő simaizmok kapcsolódnak (szőrborzolás, libabőr).
- A szőrtüsző alapján található egy hagymaszerű megvastagodás, aminek osztódó sejtjei hozzák létre a szőrszál szaruanyagát.
- A haj, szőr anyaga a keratin.
A bőralja
Rostos kötőszövete az alatta levő izomhoz vagy csonthoz kapcsolja a bőrt. Helyenként jelentékeny mennyiségű zsírszövetet tartalmaz.
A zsírszövet
- egyrészt tápanyagraktár,
- másrészt fontos szerepe van a hőszigetelésben és a
- mélyebben húzódó szövetek, szervek mechanikai védelmében.
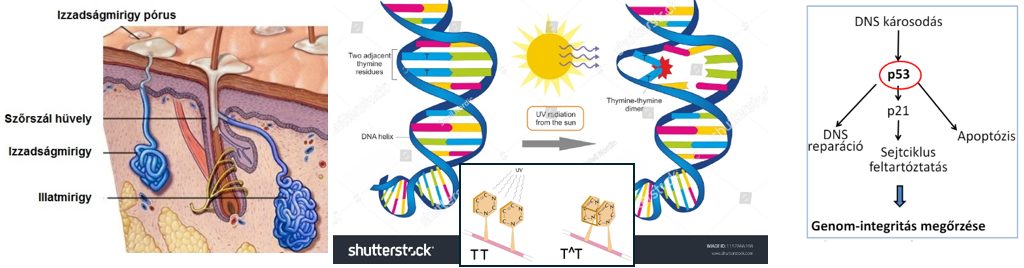 |
Hozza összefüggésbe a bőr kiválasztó feladatát a szervezet víztartalmának szabályozásával.
A bőr kiválasztó feladata szorosan összefügg a szervezet víztartalmának szabályozásával, mivel a bőr – elsősorban a verejtékmirigyek révén – fontos szerepet játszik a víz- és elektrolitháztartás fenntartásában. Ez a funkció különösen a homeosztázis megőrzése szempontjából lényeges.
Az élettani egyensúly érdekében a szervezet folyamatosan szabályozza a verejtékezés mértékét az aktuális belső és külső körülményekhez igazodva.
Összefüggés a bőr és a vízháztartás között
- A kisebb verejtékmirigyek magas víztartalmú, enyhén sós váladékot termelnek.
- A verejtékezés elsődleges célja a testhőmérséklet szabályozása, de ezzel együtt víz- és sóvesztést is eredményez.
- A verejték összetevői: víz, nátriumion, kloridion, káliumion, karbamid stb.
Kiválasztás és méregtelenítés
- A verejtékkel nemcsak víz, hanem bomlástermékek is távoznak (pl. karbamid, tejsav), így a bőr bizonyos értelemben kiválasztószervként is funkcionál.
- Ez azonban másodlagos jelentőségű a vese kiválasztó funkciójához képest.
Vízvesztés és annak következményei
- Meleg környezetben vagy fizikai aktivitás során a verejtékezés jelentősen nőhet → akár több liter víz is távozhat naponta.
- Ha ez a víz nem kerül pótlásra, dehidratáció alakulhat ki, ami befolyásolja a vér térfogatát, vérnyomást és a sejtek működését.
- A szervezet osmoreceptorai (pl. a hipotalamuszban) érzékelik a vér ozmotikus koncentrációjának, ozmózisnyomásának növekedését. Ha a vízvesztés fokozódik:
- Antidiuretikus hormon (ADH) szabadul fel → a vese gyűjtőcsöveiben fokozódik a víz visszaszívódása.
- A verejtékezés is csökken, a test próbálja minimalizálni a további vízvesztést.
Magyarázza ábra segítségével az UV-sugárzás DNS-re gyakorolt hatását a bőr egyes daganatainak kialakulása során.
Az UV-sugárzás DNS-re gyakorolt hatása a bőr egyes daganatainak kialakulása során
Az UV-sugárzás (ultraibolya sugárzás), különösen a UV-B típusú sugárzás (280–315 nm), károsíthatja a DNS-t, és ezzel kulcsszerepet játszik a bőrdaganatok kialakulásában.
Az UV-sugárzás DNS-károsító hatása, DNS-mutációk létrejötte
Az ultraibolya sugárzás a DNS-ben fotodimer termékeket eredményez, amelyek szomszédos timin bázisok kémiai összekapcsolódásával keletkeznek. Ezek a termékek torzulást eredményeznek a kettős spirál szerkezetében, és megakasztják a DNS-polimeráz haladását.
Onkogének és tumorszuppresszor gének mutációi
Az UV-sugárzás DNS-károsító hatása a bőrsejtekben mutációkhoz vezethet. Azok a mutációk okoznak daganatot, amelyek a sejtciklust szabályozó fehérjék működését érintik. A daganatkialakulás szempontjából érintett gének
- a protoonkogének, melyek olyan fehérjéket kódolnak, amik növekedési jeleket fognak fel a sejtekben. E géneknek mutációi eme fehérjék aktivációját idézik elő, aminek hatására az érintett sejt úgy érzékeli, mintha a növekedési jelzés folyamatosan jelen volna. Az aktivált protoonkogéneket onkogéneknek nevezzük.
- Tumor-szuppresszorok, daganatelnyomó gének fehérjéi (p53) működésük során féken tartják a sejtosztódást, a sejtciklust hiba esetén leállítják, beindítják az apoptózist.
- A DNS-hibajavítók és
- az apoptózis-szabályozók (sejthalálgének), amelyek olyan fehérjéket kódolnak, melyek a programozott sejthalált beindítják.
Leggyakoribb célpont: p53 tumorszuppresszor gén
- A p53 fehérje segít felismerni és javítani a DNS-károsodást, illetve beindítani az apoptózist.
- UV-sugárzás hatására mutálódhat → sejtek hibás DNS-sel is tovább osztódhatnak, ami tumor kialakulásához vezethet.
Melanoma malignum a legsúlyosabb, leginkább áttétet adó bőrrák, a direkt UV-hatás, az intenzív napozás (pl. leégés) jelentős kockázati tényező.
Megelőzés lehetősége
- Fényvédő krémek (UV-szűrővel)
- Napozás kerülése 11–15 óra között
- Védőruházat, kalap, napszemüveg
- Rendszeres bőrgyógyászati szűrés (anyajegyek ellenőrzése)
4.3. A mozgás |
Értelmezzen a csontok kémiai összetételére vonatkozó vizsgálatokat.
A csontszövetet összetétel szerint:
- szerves és
- szervetlen állományra oszthatjuk.
A szerves állományt:
- csontsejtek és
- kollagénrostok alkotják.
A szervetlen állományt a sejtközötti állomány szervetlen sói alkotják:
- Ca5(PO4)3 X, ahol X= F, OH (apatitok),
- CaCO3.
A szerves állomány a csontok rugalmasságát, a szervetlen állomány a csontok szilárdságát biztosítja.
- A szervetlen állományt sósavas kezeléssel eltávolíthatjuk, ekkor a csontok gumiszerűen hajlíthatók lesznek.
- A szerves állományt égetéssel távolíthatjuk el, ekkor a csontok elvesztve rugalmasságukat ridegek és törékenyek lesznek.
A csontszövet koncentrikus felépítésű, ún. oszteonokból épül fel. Egy oszteon kb. 10 mm magas henger, melynek közepén a centrális csatornában vérér fut. Az oszteonban a nyúlványos csontsejtek összekapcsolódva koncentrikus körökbe rendeződnek.
A sejtek között az alapállomány szintén körkörösen rendeződött lemezei futnak, amelyet szervetlen sók, kollagénrostok és szerves kötőanyag épít fel. A csontszövet vérerekkel átszőtt, élénk anyagcseréjű szövet.
A csontszövet felületét az ún. csonthártya borítja, amely sok rostot tartalmazó kötőszövetes lemez. A csonthártyában futnak a csontszövetet ellátó erek és idegek.
https://www.youtube.com/watch?v=eEDgZTlpNCc
Magyarázza a mozgási szervrendszer lényegi működését fizikai (emelő-elv, erő, erőkar), biokémiai (aktin, miozin, kreatin-foszfát, ATP, biológiai oxidáció, erjedés), szövettani (vázizomszövet) ismeretei alapján.
Magyarázza, miért szükséges az izomműködéshez Ca2+- ion, illetve Mg2+-ion.
Magyarázza az izom saját energiatároló és oxigéntároló molekuláinak szerepét.
A mozgási szervrendszer fizikai működése (emelő-elv, erő, erőkar)
Biomechanika
A biomechanika a mechanika törvényszerűségeit alkalmazza az élő szervezetekre. Az emberi test mechanikus működése egyes esetekben hasonlatos az egyszerű emelők működéséhez.
Az emelő egy rögzített (forgás)tengely (forgáspont) körül elfordítható merev rúd, amely használható teher emelésére vagy mozgatására. A tengely vagy forgáspont az emelő elforgathatóan megtámasztott pontja.
Kétoldalú (kétkarú) emelőről beszélünk, ha a teher(erő) (súly) és az aktív erő a tengelyhez képest az emelő különböző oldalán található.
Egyoldalú emelőről beszélünk, ha a teher és az aktív (emelő) erő az emelő ugyanazon oldalán van a tengelyhez képest.
- A forgáspont és a teher közötti távolság a teherkar (k1),
- a forgáspont és az aktív erő közötti távolság pedig az erőkar (k2).
Az erő forgató hatását megadó fizikai mennyiséget forgatónyomatéknak hívjuk. Egy erő forgatónyomatékát megkapjuk, ha az erő nagyságát megszorozzuk az erőkarral.
Forgatónyomaték = Erő x Erőkar
Erő = Tömeg x Gyorsulás F = m x g
Az emelő törvénye kimondja, hogy egyensúly esetén az aktív erőnek a forgáspontra vonatkozó forgatónyomatéka egyenlő a teher forgatónyomatékával:
Emelőelv: Teherkar × Tehererő = Erőkar × Erő azaz G×(k1) = F×(k2)
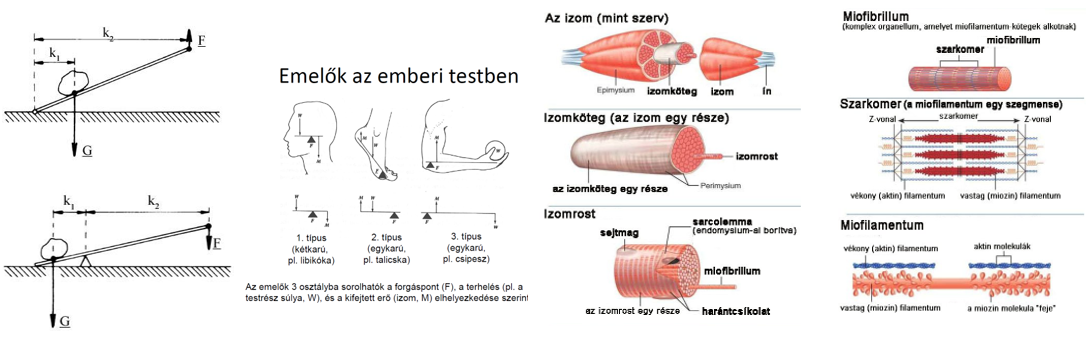 |
A mozgási szervrendszer biokémiai működése (aktin, miozin, kreatin-foszfát, ATP, biológiai oxidáció, erjedés), szövettani szerkezete
A Ca2+- ion, illetve Mg2+-ion szerepe az izomműködésben
A vázizmok felépítése lásd még 3.4.4. fejezet
- A harántcsíkolt vázizomszövet szöveti egysége a hajszálszerű, akár több ezer sejtmagot is tartalmazó, óriás sejteknek tekinthető izomrost (muscle fiber).
- Az izomrostokat membrán határolja, mely helyenként betüremkedve eléri a SER-t.
- Sok izomrost köteggé – izomnyalábbá – szerveződik, amelyet kötőszövetes hártya (perimysium) tart össze,
- az izomrostkötegek tovább rendeződve alkotják az izmokat, amelyeket szintén egy kötőszövetes lemez, az ún. izompólya (epimysium) határol.
Az izomroston belül továbbá megkülönböztetünk
- izomfonalkötegeket (miofibrillumokat),
- amelyek izomfonalakból (miofilamentumokból) állnak.
- Az izomfonalkötegek között nagy mennyiségű mitokondrium helyezkedik el.
- Az ugyancsak nagyszámú sejtmag perifériásan az izomrost membránja alatt található.
- A Ca-raktárként működő endoplazmatikus retikulum(SER) harisnyaszerűen veszi körbe az egyes izomfonalkötegeket.
A miofibrillumok felépítése és az izom-összehúzódás
A vázizmot fény- vagy elektronmikroszkópban vizsgálva az izomrostok harántirányban – a hossztengelyre merőlegesen – csíkozottságot mutatnak, azaz világosabb és sötétebb sávok váltják egymást. A sávozottságot az okozza, hogy az izomfonalkötegek a hossziránynak megfelelően szakaszosan váltakozva más-más felépítésűek, de az egyforma szakaszok pontosan egymás mellett helyezkednek el. Az izomfonalak már csak elektronmikroszkópban láthatók. Kétféle izomfonal váltakozik hosszirányban:
- a vékony fonal csak a világos szakaszokat építi fel,
- a vastag fonal nagyrészt a sötét sávokban található.
A vékony fonalakban a globuláris aktin monomerek – G-aktin – összekapcsolódva két egymás köré tekeredő, gyöngysorszerű fonalat képeznek, az F-aktint. A vékony fonalakat a Z-lemeznek nevezett képlet tartja egybe. A két Z-lemez közé eső szakaszt szarkomernek nevezzük. A szarkomera az izomfonalköteg periodikusan ismétlődő egysége.
A vastag fonal a miozin fehérjéből áll, melynek ATP-bontó enzimhatása van. A vastag fonalat 24 miozinmolekula építi fel (mint a kötegbe fogott golfütők).
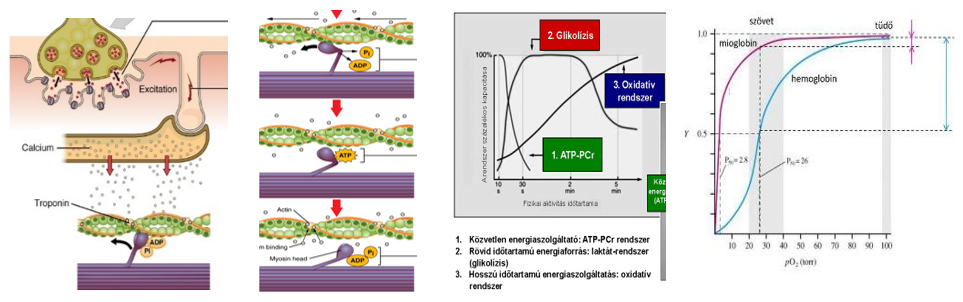 |
Az izom-összehúzódás csúszómechanizmusa
Egy elernyedt, nyugalomban lévő izomban a vékony fonalak csak kis mértékben nyúlnak be a vastag fonalak közé, a vékony és a vastag fonalak közötti kapcsolódások száma csekély. Az izom-összehúzódás során a vékony fonalak becsúsznak a vastag fonalak közé, aminek köszönhetően a Z-vonalak egymáshoz közelítenek, s így a szarkomerek megrövidülnek. Az egyes szarkomerek megrövidülése kismértékű, de mivel az izom-összehúzódás mindig az izom hossztengelyével párhuzamosan zajlik, méretcsökkenésük összeadódik, ami az izom nagymértékű megrövidülését eredményezi.
Lépések röviden
- Az izomrostokat beidegző idegrostok végződéseiből felszabaduló ingerületátvivő anyagok hatására az izomrostok citoplazmájában Ca2+-ok szabadulnak fel (SER-ből), aminek hatására a miozinmolekulák feji részei hozzákötődnek az aktinfonalakhoz.
- A kötődést követően a miozin feji részei bólintó mozgást végeznek (konformációváltozás), aminek köszönhetően a vékony fonalak beljebb csúsznak a vastag fonalak közé, egységnyi távolságot megtéve.
- Ezután az ATP és Mg2+ megkötésekor a miozinfejek leválnak az aktinfonalakról.
- ATP bontásakor felszabaduló energia hatására a miozin fejek kiegyenesednek, majd már egy másik kötőhelyen ismét hozzákötődnek a vékony fonalakhoz és kezdődik minden elölről.
A fenti lépések sokszori ismétlődése miatt a szarkomerek megrövidülnek. Az összehúzódás addig tart, ameddig a Ca-ion jelen van. A miozinfejek mozgása, alakváltozása energiaigényes, melynek fedezetét az ATP-hidrolízis energiája jelenti. Ez az izom-összehúzódás csúszómechanizmusa. A csúszás a Z-vonalakat egymás felé közelíti, a vékony fonalaknak megfelelő világos sáv szélessége csökken, ugyanakkor a sötétebb átfedési sáv szélessége nő, az összehúzódás alkalmával tehát a szarkomerek rövidülnek meg és nem az izomfonalak.
A vázizomrostokat az idegrendszer idegrostok útján működteti. Az idegrostok ingerülete szinapszisok közvetítésével átterjed az izomrostokra és akciós potenciált vált ki. A Ca2+ hiánya fokozza az ideg-izom ingerlékenységet, csökkenti az izomrostok ingerküszöbét, ami izomgörcsökhöz vezet. A Mg2+ hiánya hasonló következményekkel jár, hiszen az ATP nem önmagában, hanem Mg2+-ionnal együtt kötődik a miozin feji részéhez. A Mg-ATP-komplex kötődés hatására az aktin-miozin kapcsolat bomlik. Ha nincs Mg-ATP, akkor a miozinfej hozzákapcsolódva marad az aktinhoz (rigor állapot).
Az izomműködés energiaforrásai, az izom saját energiatároló molekuláinak szerepe
A tápanyagokat – zsírokat, szénhidrátokat (glümolekuláinak szerepe;kózt), aminosavakat – az izomrostok a vérből veszik fel. A pihenő izom a szénhidrátokat jelentős mennyiségben képes tárolni glikogén formájában. Az izom-összehúzódás közvetlen energiaforrása az ATP-hidrolízis energiája. Az ATP egyrészt a biológiai oxidáció, másrészt relatív oxigénhiány esetén a tejsavas erjedés során jön létre. A vázizomszövetben az ATP-n kívül az energiatárolásban egy másik vegyület, a kreatin-foszfát is közreműködik. A kreatin-foszfát feladata, hogy a kimerülő ATP-készleteket regenerálja.
kreatin-P + ADP = kreatin + ATP
Az ábrán az egyes energiaforrások százalékos megoszlásának változása látható egy 5 perces maximális terhelés során.
- Az első 7 másodpercben az ATP-kreatin-foszfát rendszer a domináló,
- míg a 7-60 másodperc közötti időszakban a glikolízis,
- majd a biológiai oxidáció veszi át a főszerepet.
Az izom saját oxigéntároló molekuláinak szerepe
Ismert a hemoglobinon kívül egy másik oxigénkötő fehérje is az emberi szervezetben, a mioglobin, amely a vázizmokban található. A mioglobin csupán egy polipeptidláncból és egy vastartalmú hemből álló összetett fehérje. A két fehérjében az oxigénkötődés mechanizmusa hasonló, ugyanakkor a mioglobin alacsonyabb pH-n és alacsonyabb oxigénkoncentráció esetén is képes kötni az oxigént, hiszen feladata az oxigén raktározása. Tartós összehúzódás vagy hosszantartó izomműködés esetén, amikor a vázizmok oxigénellátásának hatékonysága csökken, a mioglobinhoz kötött oxigén leválva átmenetileg képes kompenzálni a kialakult oxigénhiányt.
4.4. A táplálkozás |
Tervezzen egyszerű biokémiai kísérletet a szénhidrát-, zsír- és fehérjeemésztésre vonatkozóan.
Szénhidrátbontás
Amiláz enzim hatása a keményítőre
Anyagok, eszközök
- Nyál (emberi vagy mesterséges amiláz)
- Keményítőoldat
- Jódoldat (Lugol-oldat)
- Kémcsövek
- Vízfürdő (37 °C)
- Cseppentő
A kísérlet menete
- Tegyünk egy kémcsőbe keményítőoldatot, majd adjunk hozzá amilázt.
- Helyezzük a kémcsövet 37 °C-os vízfürdőbe.
- 5 perc után cseppentsünk egy kis mintát egy fehér csempére, majd adjunk hozzá jódcseppet.
- Ha a jód nem színeződik liláskékre a keményítő lebomlott → az amiláz működött.
https://www.youtube.com/watch?v=eAUOV4vzhbs
Zsírbontás
Lipáz enzim hatása tejzsírra
Anyagok, eszközök
- Lipáz
- Zsíros tej
- pH-indikátor (pl. fenolftalein vagy univerzális indikátor)
- Nátrium-karbonát oldat (az enyhén lúgos környezethez)
- Kémcsövek
- Vízfürdő (37 °C)
A kísérlet menete
- Keverjünk össze tejet, lipázt, pH-indikátort, kis mennyiségű nátrium-karbonát oldatot egy kémcsőben.
- Helyezzük a kémcsövet 37 °C-os vízfürdőbe.
- Figyeljük meg a színváltozást, ahogy a lipáz zsírsavakat szabadít fel, a pH csökken → az indikátor színe megváltozik → a zsírbontás bizonyítéka.
https://www.youtube.com/watch?v=UhCm-cqd2ko
Emelt szintű írásbeli feladat
A neutrális zsírok és szerepük az ember szervezetében
Kísérletelemzés
A tejben levő zsír (elsősorban neutrális zsír) emésztését vizsgáljuk. A kémcsőben tej, fenolftalein indikátor, NaOH-oldat, lipáz oldat található.
A zsíremésztést fenolftalein sav-bázis indikátorral követjük nyomon. (A fenolftalein lúgos közegben rózsaszín, semleges és savas közegben színtelen.) A kémcsövek tartalmához nátriumhidroxid-oldatot csepegtetünk addig, amíg az indikátor gyengén lúgos kémhatást nem jelez.
Fehérjebontás
Pepszin hatása tojásfehérjére
Anyagok, eszközök
- Pepszinoldat
- Tojásfehérje (főzött, majd kis darabokra vágva)
- Sósav (0,1 M – gyomorsav modellezéséhez)
- Kémcsövek
- Vízfürdő (37 °C)
A kísérlet menete
- Tegyünk egy kis darab tojásfehérjét egy kémcsőbe.
- Adjunk hozzá pepszint és néhány csepp híg sósavat.
- Tegyük a kémcsövet 37 °C-os vízfürdőbe.
- Időnként rázzuk meg, majd figyeljük meg: a fehérje fokozatosan feloldódik, ami az emésztés bizonyítéka.
https://www.youtube.com/watch?v=RsV5z-7jCdA
Emelt szintű írásbeli feladat
Fehérjék emésztése
Tanulók egy kísérlet során azt vizsgálták, hogy a gyógyszertárban kapható pepszin milyen kémhatású közegben bontja le a fehérjéket. Négy számozott (1, 2, 3, 4) lombikot helyeztek a lombikállványra, melynek mindegyikébe egy főtt tojás fehérje részének vékony szeletét tették. A lombikokba a következő összetételű, azonos térfogatú oldatokat öntötték. (A sav, illetve a lúg pH-ja az emberi bélcsatorna megfelelő szakaszaira jellemző érték volt.)
- 1) pepszin + sav
- 2) pepszin + semleges oldat
- 3) pepszin + lúg
- 4) kontroll: (víz +) sav/lúg + fehérje, ami azt bizonyítja, hogy nem a sav/lúg hatására emésztődik a tojásfehérje.
Értelmezze a máj makroszkópos anatómiai és mikroszkópos szövettani, illetve a vékonybél keresztmetszeti képének szövettani ábráit.
A máj
A középbél nagy emésztőmirigyei a máj és a hasnyálmirigy. A máj emberben közel másfél kilogramm tömegű, vörösbarna színű szerv. A rekeszizom jobb kupolájába illeszkedik be. Felső része domború, alsó része lapos. Jobb és bal lebenyre osztható.
A májba két ér hozza a vért:
- a májartéria, mely az aortából ered, és friss, oxigéndús vért szállít, valamint
- a májkapuvéna, ami a máj funkcionális ere, a bélcsatorna felől – kivéve a végbél alsó kétharmadát, a hasnyálmirigyet és a lépet – szállít tápanyagokban gazdag vért.
A májba lépő erek az ún. májkapun lépnek be a máj állományába. A máj szöveti szerkezete rendkívül bonyolult. A máj szövettani és működési egysége a májlebenyke, mely egy hengeres test, sugarasan futó májsejtgerendák építik fel. A májba futó erek – a májartéria ill. a májkapuvéna – egyre kisebb erekre ágaznak, végül a májlebenyke szélén egymás mellé kerülnek és vértartalmuk keveredik. Belőlük erednek az egyes májsejtgerendák között futó májkapillárisok, amelyek a májlebenyke közepén a centrális vénában egyesülnek. A centrális vénák egyre nagyobb vénákká folynak össze, végül májvéna hagyja el a májkapun keresztül a májat, módosított összetételű vért szállítva a testvénába (alsó üres véna).
Az epeutak a májsejtek közötti epekapillárisokkal kezdődnek, melyek egyre nagyobb vezetékekké egyesülnek, végül az epevezetékbe torkollva ugyancsak a májkapun át hagyják el a májat.
https://www.youtube.com/watch?v=QA7HdYTmqMQ
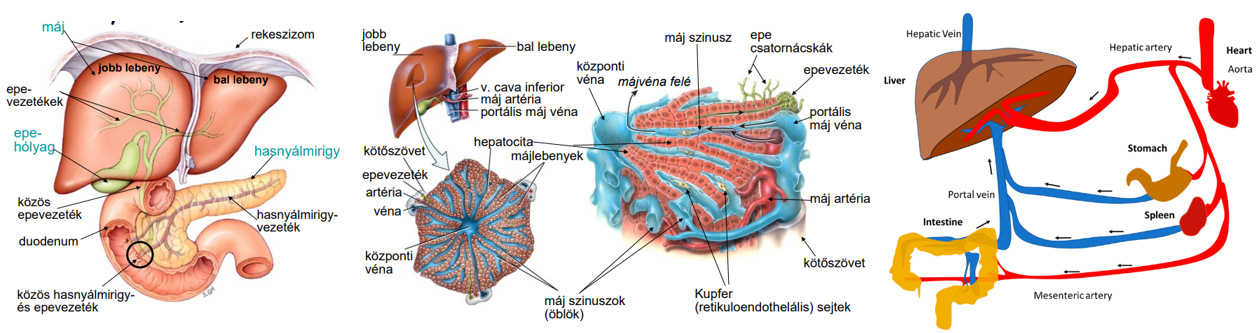 |
A vékonybél, középbél
A vékonybél a tápcsatorna középső szakasza – ezért lehet középbélnek is nevezni –, az emésztés befejezésének és a tápanyagok felszívásának fő helye. Az itt ható emésztőnedveket a középbélhez kapcsolódó máj és hasnyálmirigy termelik. Emberben kb. 6-7 m hosszú, és 3 fő szakaszra osztható:
- epésbél vagy patkóbél (duodeum),
- éhbél (jejunum) a nevét onnan kapta, hogy a nagymértékű felszívás miatt a halál beállta után nem található benne ételmaradék,
- csípőbél (ileum).
A patkóbél nevét alakjáról, illetve arról kapta, hogy ide nyílik a máj és a hasnyálmirigy közös vezetéke, tehát ide ömlik az epe és a hasnyál. A patkóbél kanyarulatában foglal helyet a hasnyálmirigy. A középbél szélesen kanyargó hurkokat vet, ezeket bélkacsoknak nevezzük.
A vékonybél nyálkahártyája az alatta fekvő réteggel együtt sűrűn álló, magas redőket vet. A hámréteget egyrétegű mikrobolyhos hengerhám alkotja. A középbél legjellegzetesebb képződményei a felületnövelő bélbolyhok, amelyek kesztyűujjszerű kiemelkedései a nyálkahártyának. A bélbolyhok tengelyét
- laza kötőszövet,
- simaizomsejtek,
- hajszálérhálózat,
- idegfonat,
- és egy, a csúcs közelében vakon kezdődő centrális nyirokér tölti ki.
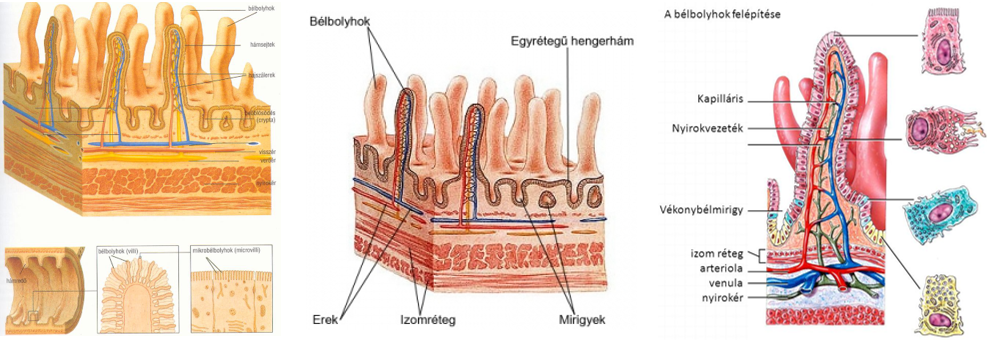 |
Értelmezzen a táplálékfelvétel és a testtömeg szabályozására vonatkozó ábrát, szöveget, illetve adatokat, a szabályozásban résztvevő peptidek szerepével, a folyamatok hátterében álló magatartási folyamatokkal összefüggésben.
A táplálkozás során a táplálékkal vesszük fel a szervezetünk felépítéséhez és az életfolyamatok fenntartásához szükséges tápanyagokat. Szűkebb értelemben táplálkozás alatt csak a táplálék bevitelét értjük a szervezetbe, de tágabb értelmezésben ide soroljuk a táplálék anyagainak megemésztését, felszívódását és az anyagcserében történő hasznosulását, illetve az emészthetetlen anyagok kiürítését is.
A táplálék tartalmazza a létfontosságú tápanyagokat, ill. a szervezet számára fel nem használható, felesleges anyagokat (salakanyagok).
A tápanyagok funkció szerint csoportokba sorolhatók:
- testépítő anyagok, pl. fehérjék,
- energiaforrások, pl. szénhidrátok, zsírok,
- enzimek koenzimjeinek összetevői, pl. vitaminok,
- enzimek prosztetikus csoportjai, pl. egyes ásványi anyagok.
Továbbá fontos tápanyag a víz, mely szerkezetmeghatározó, oldószer, reakciópartner és reakcióközeg, szállítóközeg.
Minőségi éhezés kialakulása során elegendő mennyiségű táplálék kerül a szervezetbe, de abban nem megfelelő az egyes tápanyagok mennyisége, ill. aránya, leggyakrabban ásványi anyagok, vitaminok, esszenciális aminosavak hiánya okoz problémát.
Mennyiségi éhezés során az elfogyasztott tápláléknak a kelleténél kevesebb a mennyisége és az energiatartalma, ezért a bevitt táplálék nem fedezi a napi energiaszükségletet.
Az éhség egy motivációs állapot, a táplálék felkeresésére, a táplálék elfogyasztására irányuló belső késztetés. Kiváltásában többféle élettani tényező – pl. alacsony vércukorszint, gyomor üressége stb. – játszik szerepet.
- A fiziológiás éhségérzet kialakulása – a hipotalamuszban – alapvető biológiai szükséglet kielégítését teszi lehetővé, ettől meg kell különböztetnünk
- a mentális éhség fogalmát, amit étvágynak nevezünk, mely létrejöttének hátterében inkább külső tényezők – stresszhatások, ingerszegény környezet (unalomevés), különféle beidegződések-társítások (TV, moziéhség, egy kedvenc étel látványa, szaga, íze, emléke) stb. – keresendők.
A táplálkozással kapcsolatos idegi szabályozó mechanizmusok központjai a hipotalamuszban vannak. Itt található
- az éhségközpont, amelynek aktivitása éhségérzetet kelt és ez a táplálék felvételét eredményezi, ill. ugyanitt van
- az ellentétes hatású jóllakottsági központ.
Az éhség érzetét elsősorban az alacsony vércukorszint és olyan peptidhormonok – pl. ghrelin szöveti hormon – váltják ki, amiket a gyomor üressége esetén elsősorban a gyomor fala választ el.
Ez a szöveti hormon továbbá
- mivel hat a májra és a hasnyálmirigyre, elősegíti a tápanyagok glikogén, ill. zsír formájában történő raktározását,
- a központi idegrendszerben serkenti az agy jutalmazó központjában a dopaminfelszabadulást, aminek hatására fokozódik az evéssel együttjáró örömérzet.
Szintje a vérben étkezés előtt a legmagasabb és közvetlenül utána a legalacsonyabb. Testsúlycsökkenéskor nő a vérben a ghrelinszint, hízáskor pedig csökken.
A jóllakottság jelzései
- a gyomor teltsége (mechanoreceptorok révén),
- a magas vércukorszint és inzulinszint, továbbá
- a zsírszövet sejtjeiben termelődő leptin szöveti hormon.
A leptin a ghrelinhez hasonlóan a hipotalamikus neuronokon – jóllakottsági központ – keresztül fejti ki a hatását. A tartósan alacsony leptinszint folyamatos éhségérzetet okozhat, ezért a leptinhiány már kora gyermekkortól jelentkező súlyos elhízást eredményez. Ennek ellenére sok esetben az elhízás magas leptinszint mellett is kialakul. Ezt a jelenséget leptinrezisztenciának nevezzük.
Az alapanyagcsere a szervezet fenntartásához szükséges minimális energia termelése. A napi energiaszükséglet 60-70 %-a létfenntartó működések energiaszükségletét fedezi, mint pl. keringés, kiválasztás, májműködés, nyugalmi idegi szabályozás, alapizomtónus fenntartása, hőszabályozás.
Meghatározása a hőleadás vagy indirekt módon, az oxigénfogyasztás mérésével történik, a következő körülmények között:
- éber, nyugalmi állapotban,
- ruhátlanul, semleges külső hőmérsékleten (28-30 °C, ahol a hőtermelés mértéke minimális),
- 12–24 óra előzetes éhezést követően, gyógyszerhatástól mentesen.
Nagyrészt örökletes tényezők határozzák meg, de mértékét befolyásolja
- a kor (gyerekeknél magasabb),
- a nem (férfiaknál magasabb a nagyobb izomtömeg miatt),
- egyes hormonok (pl. tiroxin, adrenalin fokozzák),
- különféle élettani állapotok (pl. terhesség, szoptatás növeli).
Az alapanyagcsere átlagos energiaértéke nőknél 6000 KJ, férfiaknál 7000 KJ körüli naponta.
Az étrendi elvárások ennek megfelelően változnak tevékenységtől, kortól, nemtől és élettani állapottól függően. A napi energiaszükségletet számolással is meghatározhatjuk közelítőleg úgy, hogy az alapanyagcsere értékét megszorozzuk a fizikai aktivitás mértékétől függő korrekciós faktorral.
Energiaigény-kalkulátor https://www.okostanyer.hu/energiaigeny-kalkulator/
Alultápláltság akkor alakul ki, ha nem kerülnek be a szervezetbe az egészséges táplálkozásnak megfelelően a szükséges mennyiségű és minőségű tápanyagok, továbbá, ha a tápanyagok emésztésében, ill. felszívódásában zavarok lépnek fel. Mindezek hatására hosszútávon
- különféle hiánybetegségek alakulnak ki – esszenciális aminosavak, vitaminok, ásványi anyagok hiánya miatt –, ill.
- kialakulhat ödéma a vérfehérjék koncentrációjának csökkenése miatt,
- felborulhat a szervezet víz- és ionegyensúlya (K+),
- nőhet a fertőzésekre való hajlam az immunrendszer gyengülése miatt,
- izomerőgyengeség, vérszegénység, csontritkulás stb. következhet be.
Ha az elfogyasztott táplálék energiatartalma rendszeresen meghaladja az energiaszükségletet, túltápláltság alakul ki, ami elhízáshoz vezet. Minden felesleges mennyiségben felvett tápanyag az emberi szervezetben zsírrá alakul és a zsírszövetekben halmozódik fel. Az elhízás számos egészségügyi problémát eredményez,
- fokozódik az érelmeszesedés, aminek következménye a magasvérnyomás-betegség, nő a szívinfarktus és az agyi infarktus kockázata,
- emelkedik a cukorbetegség, ill.
- a daganatos betegségek veszélye,
- a csontrendszer, az ízületek túlterhelődnek.
Az elhízás részben örökletes tényezőkre, részben környezeti okokra – helytelen táplálkozási szokások, mozgásszegény életmód – vezethető vissza.
A testtömegindex (BMI) a felnőtt szervezet egészséges testsúlyának meghatározására szolgál, melynek értékét úgy kapjuk meg, hogy a testsúlyt elosztjuk a méterben megadott testmagasság négyzetével (kg/m2). Kiszámolásával megtudható, mennyi a súlyfelesleg,
- 20 és 25 között egészséges a testsúly,
- 20 alatti érték esetén soványságról,
- 25 feletti értéknél túlsúlyosságról,
- 35 feletti értéknél már kóros elhízásról beszélünk.
A testtömegindex értékét továbbá befolyásolja a nem, az életkor. A testtömegindex értéke olykor megtévesztő lehet, pl. nagyobb testmagasság esetén kisebb, ill. sportolóknál jelentősebb izomtömeg esetén magasabb értéket mutathat.
4.5. A légzés |
Értelmezze a Donders-modellt bemutató ábra alapján a légzőműködéseket.
Légzőmozgások
A külső légtér és a tüdő légtere közötti légcserét a mellkas térfogatát változtató légzőmozgások teszik lehetővé. A tüdőnek saját izomzata nincs – leszámítva az egyes légutakat szűkítő simaizmokat –, így önálló mozgásra képtelen, a mellhártyák közvetítésével belülről rátapad a mellkasfalra, passzívan követi annak mozgását, térfogatváltozásait. Ez a térfogatváltozás a vázizmok közé sorolható, légzőizmoknak nevezett
- bordaközi izmok és
- a rekeszizom működésének köszönhető.
Nyugodt belégzéskor
- a külső bordaközi izmok felemelik a mellkast, ill.
- a kupolaszerű rekeszizom összehúzódva ellaposodik.
Az aktív izomösszehúzódásoknak köszönhetően
- a mellüreg térfogata megnő,
- a tüdő kitágul,
- s benne a nyomás lecsökken,
- így a levegő kívülről a tüdőbe áramlik.
Attól függően, hogy a belégzésben mely izmok működése dominál, megkülönböztetünk hasi légzést és mellkasi légzést.
A hasi légzésben a rekeszizom vesz főleg részt, ez inkább a tüdő alsóbb részeit tölti meg levegővel, a férfiak, a gyerekek, ill. a sportolók körében gyakoribb.
A mellkasi légzésnél inkább a bordaközi izmok működése hangsúlyosabb, így elsősorban a tüdő felső része telik meg levegővel, ez inkább a nőkre jellemző.
Erőltetett belégzéskor a vállizmok is szerepet játszanak a váll felemelése révén a mellkas térfogat növekedésében.
Nyugodt kilégzés során
- a mellkas visszatér alaphelyzetébe,
- térfogata csökken,
- a tüdőben a levegő nyomása megnő,
- aminek következtében kiáramlik a tüdőből.
A nyugodt kilégzés passzív folyamat, a belégzőizmok elernyedése miatt történik, a mellkas súlyánál fogva nehezedik rá a tüdőre, s összenyomja. Ezt segíti a tüdő szövetének nagyfokú rugalmassága, ami a tüdőt összehúzni igyekszik.
Erőltetett kilégzéskor már ún. járulékos kilégzőizmok is közreműködnek, mint pl.
- a belső bordaközi izmok, amelyek összehúzzák a mellkast, ill.
- a hasizmok, amelyek a belekre gyakorolt nyomásuk révén erősebben bedomborítják a rekeszizmot a mellüregbe (hasprés).
A légzőműködések szabályozásáért felelős idegi központok az agytörzsben vannak. A légzőmozgások szabályozásában a legfontosabb inger a vér szén-dioxid-tartalma, koncentrációjának növekedése a belégzés kiváltásáért felelős neuroncsoport aktivitását fokozza.
A légcsere térfogati jellemzői
- Nyugodt légzés esetén az egyszeri légvétellel ventillált 0,5 l (dm3) levegőt légzési térfogatnak nevezzük.
- A légvételenként kicserélt levegő térfogatát megszorozva a percenkénti légvételek számával, megkapjuk a légzési perctérfogatot. Nyugodt légzés esetén a perctérfogat 8 l, mivel percenként 16-szor veszünk levegőt, légvételenként 0,5 l levegőt kicserélve.
- Fizikai, ill. pszichés terhelés hatására a perctérfogat jelentősen fokozódhat, ami egyrészt a megnövekedett légzésszámnak, ill. a megnövekedett légzési térfogatnak köszönhető.
- Erőltetett belégzéskor további 2,5 l levegőt tudunk beszívni, amit belégzési tartaléknak hívunk.
- Erőltetett kilégzés során 1,5 l levegőt tudunk még kipréselni a tüdőből, ezt kilégzési tartaléknak tekintjük.
- Ekkor még mindig marad levegő a tüdőben, amit semmilyen körülmények között nem tudunk kifújni, ez a maradék levegő, kb. 1 l.
- Az egy légvétellel maximálisan beszívott, majd erőteljesen kifújt levegő térfogatát vitálkapacitásnak nevezzük, kb. 4,5 l.
A légcsere térfogati változásait spirométerrel lehet meghatározni.
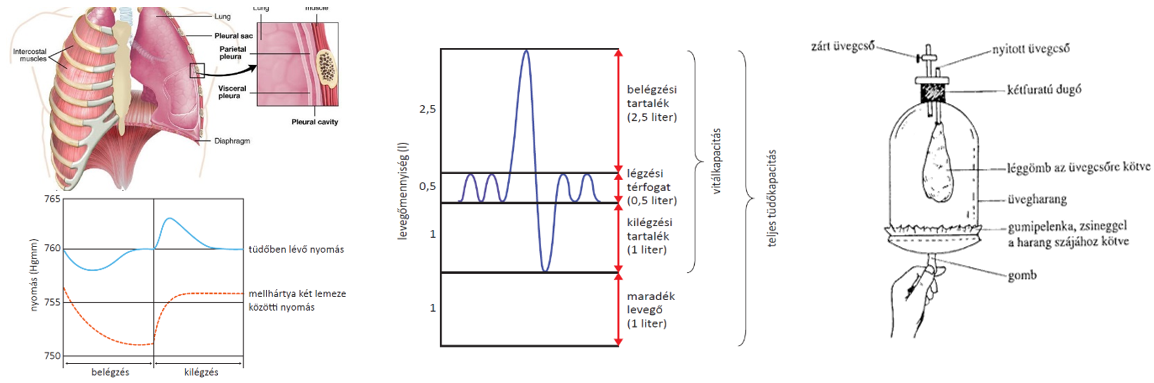 |
A légzőmozgásokkal kapcsolatos nyomás- és térfogatváltozások
Belégzés során a tüdőben eleinte csökken a nyomás, majd a folyamat második felétől ismét növekedik, amíg el nem éri újra a légköri nyomás – 760 Hgmm, 101 kPa – értékét.
Kilégzéskor eleinte növekedik a nyomás a tüdő légterében, majd a folyamat második felétől elkezd csökkenni addig, ameddig egyenlő nem lesz ismét a légköri nyomással.
A mellhártya két lemeze között a nyomás végig kisebb, mint a tüdőben, ami biztosítja, hogy a tüdő rátapadjon a mellkas belső felszínére. A különbség a tüdőben mérhető nyomásváltozáshoz képest, hogy a belégzés során mindvégig csökken, a kilégzés során pedig mindvégig nő a nyomás a két lemez között. Amennyiben a mellhártya két lemeze közé levegő kerül, a tüdőben található rugalmas rostoknak köszönhetően a tüdő kis gombóccá ugrik össze, s így képtelen követni a mellkas mozgását. Ez a jelenség a légmell.
A légzőrendszer nyomás- és térfogatváltozásainak összefüggéseit a Donders-féle tüdőmodellel demonstrálhatjuk.
A kísérletet
- egy olyan harang alakú üvegbúrával (mellkas) végezzük,
- amit alulról egy gumimembrán (rekeszizom),
- felülről pedig egy kétfuratú gumidugó zár el.
- Az egyik furatban egy elzárható üvegcső található (melynek nyitásával a légmellt demonstrálhatjuk),
- a másikban egy olyan üvegcső (légcső), amelyre
- egy rugalmas léggömb (tüdő) csatlakoztatható.
- A Donders harang és a léggömb közötti tér a mellhártyák közötti teret,
- a léggömb belső tere a tüdő belső terét modellezi.
A rekeszt modellező gumimembránt lehúzva a harang és a tüdő közötti térben a légköri nyomásnál alacsonyabb nyomást hozunk létre, a változást a léggömb rugalmas fala passzívan követi, kitágul és levegő áramlik a belsejébe (belégzés).
A gumilapot elengedve, az visszatérve eredeti helyzetébe, fokozza a nyomást a harang belsejében, a léggömb összenyomódik és a levegő kiáramlik (kilégzés).
Értelmezze a tüdő makroszkópos anatómiai és mikroszkópos szövettani ábráit.
A légcső két főhörgőre oszlik, melyeket porcgyűrűk merevítenek, de egyébként a légcsővel azonos felépítésűek.
A két főhörgővel a légutak a tüdőben folytatódnak. A főhörgők egyre kisebb átmérőjű hörgőkre és hörgőcskékre ágaznak, amik végül a léghólyagokban végződnek. Az elágazódások bonyolult rendszere az ún. tüdőfát hozza létre.
A főhörgőktől a hörgőcskék felé haladva folyamatosan változik a légutak falának szerkezete. A fal felépítésében egyre kevesebb a porcszövet – a hörgőcskék falából már teljesen hiányoznak a porcok –, egyre több a kötőszövet és a simaizomszövet, a hámszövet is egyre laposodik, a léghólyagok falában már laphám található.
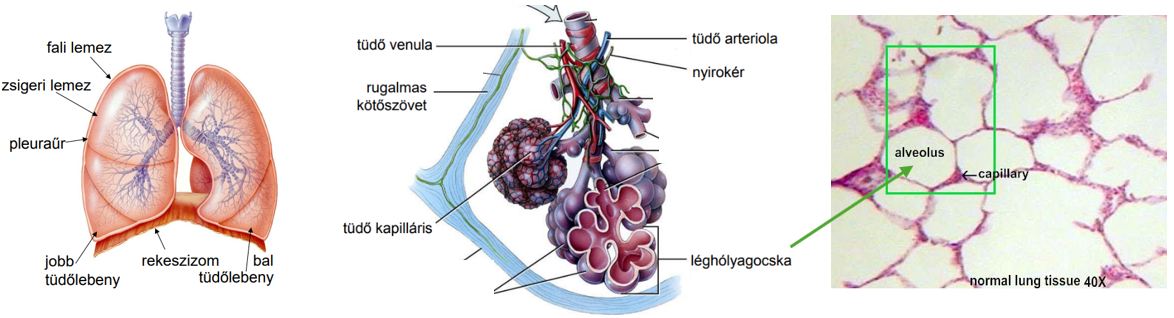 |
A tüdő a mellüregben található páros szerv, oldalról a mellkasfal, alulról a rek eszizom határolja, felső csúcsa a kulcscsontig nyúlik. Alakja kúphoz, állománya szivacshoz hasonlítható. A két tüdő egymás felé eső felszínein találhatók a tüdőkapuk, ahol a tüdők erei és a főhörgők lépnek be, ill. ki. A tüdőt mély hasadékok lebenyekre tagolják, a jobb 3, a bal 2 lebenyből áll.
A tüdőben a legutolsó hörgőcskeelágazások a szőlőfürtszerű léghólyagokba nyílnak, amelyeket léghólyagocskák bonyolult rendszere épít fel. A léghólyagocskák felülete a légzőfelület, emberben elérheti a 150 m2-t, itt zajlik a külső gázcsere. Ennek megfelelően faluk igen vékony.
A léghólyagocskák felszínén többféle sejttípust különböztetünk meg, találhatók itt
- laphámsejtek, ezek képezik a légzőfelületet, továbbá
- fagocitáló sejtek (makrofágok), melyek a léghólyagokig eljutott porszemcséket kebelezik be, ill.
- felületaktív anyagot termelő sejtek, ezek váladéka csökkenti a léghólyagocskákat borító folyadékfilm felületi feszültségét, s megakadályozza, hogy a nagyon kis átmérőjű (10 μm) léghólyagocskák felszíne kilégzéskor összetapadjon.
A léghólyagocskák közötti igen vékony válaszfalban vannak
- a tüdő hajszálerei,
- továbbá olyan kötőszöveti sejtek, amik a tüdő falában található nagy mennyiségű
- rugalmas rostot termelik.
A tüdő a mellkasban saját térfogatánál nagyobb teret tölt ki, a mellhártyák által a tüdő mintegy ki van feszítve a mellkas belső felszínére. A mellhártya a nagy savós hártyák – hashártya, szívburok, mellhártya – egyike. A két tüdőnek külön mellhártyarendszere van, melyek zsákszerűen veszik körül a tüdőfeleket. A mellhártyának két lemeze van, a külső lemez a mellkas belső felszínére, a belső lemez a tüdő külső felszínére tapad. A két lemez között súrlódást csökkentő savós folyadék található, aminek a nyomása mindig kisebb, mint a tüdőben a levegő nyomása, így a tüdő kifeszül a mellkas falára.
Elemezzen kísérletet az egyes szabályozóelemek feladatának bemutatására.
A légzés szabályozása
A légzés szabályozása lényegében a légzőizmok – rekeszizom és bordaközi izmok – működésének szabályozását jelenti. Automatikus reflexes működés, amely bizonyos határok között akaratlagosan szabályozható.
Az agytörzs területén – nyúltvelő és a híd – találhatók a légzés szabályozásáért felelős idegi központok. A nyúltvelőben megkülönböztetünk belégző- és ún. kilégzőközpontot.
- Belégzőközpont, a nyugodt légzés alapritmusát generálja automatikusan,
- a kilégzőközpontnak csak erőltetett légzésben van szerepe.
A légzés szabályozásában
- kémiai – a vér és az idegszövet sejtközötti állományának CO2- és O2-tartalma – és
- a tüdőből származó mechanikai ingerek játszanak szerepet.
A kémiai ingereket
- részben a szívhez közeli erek falában található kemoreceptorok,
- részben maguk a szabályozó központok képesek felfogni.
A belégzésben a vér széndioxid-koncentrációja ill., ami ezzel arányos, az idegszövet H+-koncentrációja a legfontosabb inger, mivel ezek a belégzőközpont működését serkentik.
A nyugodt kilégzésben a tüdő falában található feszülésérzékeny mechanikai receptorok játszanak szerepet, mivel a tüdő kitágulásakor – belégzés végén – gátolják a belégzőközpont aktivitását, megszakítva a belégzőmozgásokat, utat adva a passzív kilégzésnek (Hering-Breuer-reflex).
A nyugodt légzésben tehát csak a belégzőközpont ritmikus működésének van szerepe, a kilégzőközpont csak erőltetett (be-) és kilégzésnél szerepel, a járulékos légzőizmokat idegzi be (pl. belső bordaközi izmok, hasizmok stb.).
A szén-dioxid hatásának vizsgálata a légzésszámra
Eszközök
- stopperóra vagy telefon időmérő funkciója
- zacskó vagy papírzacskó
- jegyzetfüzet és toll
- kísérleti alany
Módszer
- Nyugalmi légzésszám mérése
- A kísérleti alany 5 percig nyugalmi helyzetben ül.
- Ezután 1 percig számoljuk a légvételeket (egy belégzés + kilégzés = 1 légzés).
- Jegyezzük fel a légzésszámot.
- Szén-dioxiddús levegő belégzése
- A kísérleti alany 30 másodpercig lélegezzen be és ki egy papírzacskóba.
- Ezzel a zacskóban a CO₂ koncentrációja nő.
- Ezután azonnal ismét mérjük meg az 1 perces légzésszámot.
- Fizikai aktivitás után is megismételhető a mérés a gyakorlat hatásának vizsgálatára (pl. 1 perc helyben futás után légzésszám mérése).
Eredmények
| Állapot | Mért légzésszám (1 perc alatt) |
| Nyugalomban | pl. 16 |
| Zacskóval való légzés után | pl. 22 |
| Fizikai aktivitás után | pl. 28 |
Elemzés
- A kísérlet során a nyugalmi állapothoz képest a szén-dioxiddal dús levegő belégzése jelentősen növeli a légzésszámot.
- Ez azt mutatja, hogy a vér CO₂-szintje kulcsszerepet játszik a légzésszabályozásban.
- A légzőközpont (az agytörzsben) érzékeli a vér pH-jának csökkenését, amit a CO₂ növekedése okoz, és ennek hatására fokozza a légzési frekvenciát.
- A fizikai aktivitás hatására is nő a CO₂-termelés, így az is fokozott légzéshez vezet.
Következtetés
A kísérlet jól szemlélteti, hogy a szén-dioxid koncentrációja a vérben az egyik legfontosabb tényező a légzés szabályozásában. A szervezet nem az oxigén, hanem főként a CO₂-szint alapján szabályozza a légzés ritmusát és mélységét.
Kapcsolja össze fizikai ismereteivel a légmell és a keszonbetegség kialakulását.
A mellhártya két lemeze között a nyomás végig kisebb, mint a tüdőben, ami biztosítja, hogy a tüdő rátapadjon a mellkas belső felszínére. Amennyiben a mellhártya két lemeze közé levegő kerül, a tüdőben található rugalmas rostoknak köszönhetően a tüdő kis gombóccá ugrik össze, s így képtelen követni a mellkas mozgását. Ez a jelenség a légmell.
A vérben nemcsak a légzési gázok oldódnak, hanem a levegőben lévő egyéb összetevők is, mint pl. kisebb mértékben a nitrogén. A keszonbetegség vagy más néven dekompressziós betegség olyan, akár halálos kimenetelű tünetegyüttes, ami akkor alakul ki, ha egy nagyobb nyomású térből hirtelen egy alacsonyabb nyomású térbe kerülünk. A keszonbetegség különösen búvárokat és a magasnyomású környezetben dolgozókat (pl. mélyépítők) fenyegeti. Nagy nyomású környezetben a vérgázok oldékonysága megnő a vérplazmában majd, ha a szervezet hirtelen alacsonyabb nyomású térbe kerül, a gázok oldékonyságának csökkenése miatt buborékok (elsősorban nitrogéngáz) keletkeznek a vérben és a szövetekben, melyek elzárhatnak kisebb ereket. A tüneteik annak megfelelően alakulnak ki, hogy milyen szerveket érinti az érelzáródás, kialakulhat rossz közérzet, fáradtság, ízületi fájdalom, bőrviszketés, izomgyengeség, bénulás, látászavarok, szédülés, egyensúlyzavar stb., súlyosabb esetben halál.
Fizikai alapismeretek
A levegőt, mint gázelegyet ún. állapotjelzőkkel jellemezhetjük, ezek a hőmérséklet, a nyomás, a térfogat, az anyagmennyiség.
A gázok térfogata az a térrész, ahol a molekulák mozoghatnak. Jele: V, mértékegysége: m3.
A gázok nyomása a felületre merőleges nyomóerő és a felület nagyságának a hányadosa. Jele: p, mértékegysége: N/m2 = Pa
Az állapothatározók kapcsolatát az ideális gázok állapotegyenlete írja le.
p.V = n.R.T, ahol R az egyetemes gázállandó, értéke 8,31 J/mol.K, a T a gáz hőmérséklete Kelvinben kifejezve, n az anyagmennyiség.
Eszerint adott hőmérsékleten, adott mennyiségű gáz nyomásának és térfogatának a szorzata állandó, másképpen, ha a gáz térfogata nő, nyomása csökken.
Hozza összefüggésbe a tüdő-léghólyagocskákat borító folyadékréteg felületi feszültségének változását a dohányzással.
A tüdő léghólyagocskáit egy nagyon vékony folyadékréteg borítja, amely nedvesíti azok felszínét. Ebben a rétegben jelentkezik egyfajta felületi feszültség. Ez az a fizikai jelenség, amikor a folyadék felszínén lévő molekulák kohéziós ereje egyfajta "összehúzó" hatást kelt. Ez a felületi feszültség össze akarja húzni a léghólyagocskákat, ami megnehezítené a belégzést.
A tüdő II-es típusú hámsejtjei surfactant, egyfajta felületaktív anyagot termelnek. Ez a foszfolipid-alapú anyag csökkenti a léghólyagocskákat borító folyadékfilm felületi feszültségét, s megakadályozza, hogy a nagyon kis átmérőjű (10 μm) léghólyagocskák felszíne kilégzéskor összetapadjon.
A dohányfüst számos toxikus anyagot (pl. szabadgyököket, kátrányt, formaldehidet) tartalmaz, amelyek többféle módon rontják a surfactant működését.
A surfactant termelő sejteket károsítja
- A II-es típusú hámsejtek károsodnak vagy elpusztulnak, így kevesebb surfactant termelődik.
- Ez növeli a felületi feszültséget, mivel nincs elegendő surfactant a feszültség csökkentésére.
A meglévő surfactant szerkezetét is károsítja
- A dohányfüst oxidatív hatása módosíthatja a surfactant foszfolipidjeit, így azok kevésbé hatékonyan csökkentik a felületi feszültséget.
A növekvő felületi feszültség miatt
- az alveolusok összeeshetnek (főleg kilégzéskor),
- megnő a légzési munka (több energiába kerül a belégzés),
- romlik a gázcsere hatékonysága (oxigénfelvétel, CO₂-leadás),
- hosszú távon ez hozzájárulhat COPD (krónikus obstruktív tüdőbetegség) kialakulásához.
4.6. Az anyagszállítás |
Elemezze az emberi vérből készült vérkenetet bemutató fénymikroszkópos képet vagy rajzolt ábrát.
A vért alkotó sejtek számának és alakjának meghatározása céljából vérkenetet készítenek, melyet fénymikroszkópban tanulmányoznak.
A vérben levő sejtes állományt, a sejtek felépítése és működése alapján 3 csoportba soroljuk.
- Vörösvértestek
- Vérlemezkék
- Fehérvérsejtek
A vörösvértestek
A vörösvértestek emlősökben sejtmag nélküli, önálló mozgásra képtelen, középen homorú, korong (bikonkáv) alakú sejtek.
Számuk férfiakban 5 millió/mm3, nőkben 4.5 millió/mm3.
A vörösvértestek szárazanyagának 95%-át a hemoglobin tölti ki.
A vörösvértestekben sejtmag, belső membránrendszer, mitokondrium nincs, s így szinte teljes tömegét a hemoglobin adja. Anyagcseréje minimális, mivel mitokondriumokat nem tartalmaz, a sejt csupán glikolízist, ill. tejsavas erjedést folytat.
A vörösvértestek feladata az oxigén és részben a szén-dioxid szállítása. A vörösvértestek átlagos élettartalma 120 nap, keletkezésük, pusztulásuk folyamatos.
A vörösvértest képzéshez szükséges B12 vitamin, folsav és vas.
Az elöregedett sejteket a lép, a máj vonja ki a forgalomból.
A vérszegénység olyan állapot, amelyben a vörösvértestek száma vagy a hemoglobin mennyisége túl alacsony a vérben.
A hemoglobin egyik közismert rendellenessége a sarlósejtes vérszegénység. A betegség oka, hogy egy pontmutáció miatt a globin egyik láncában egy aminosav kicserélődik. A rendellenes vörösvértestek citoplazmájában a hemoglobin kicsapódik, megváltoztatva a sejtek működését és az alakját. A beteg sejtek egyrészt már csak részben képesek az oxigén szállítására (kb. 50%-ban), másrészt sarlóalakúvá válnak, összecsapódnak, vérrögöket képeznek, melyek elzárhatnak kisebb ereket.
A vérlemezkék (trombociták)
A vér legkisebb alakos elemei, számuk 300 000/mm3. Nevük arra utal, hogy emlősökben szabálytalan alakú, sejtmag nélküli, apró sejttöredékek, így élettartalmuk 5-10 nap, igen rövid.
A vörös csontvelőben keletkeznek úgy, hogy az őssejtekből differenciálódó óriássejtek (megakariocyta) sejtplazmájából hasadnak le. A bennük található enzimek révén véralvadásban játszanak szerepet.
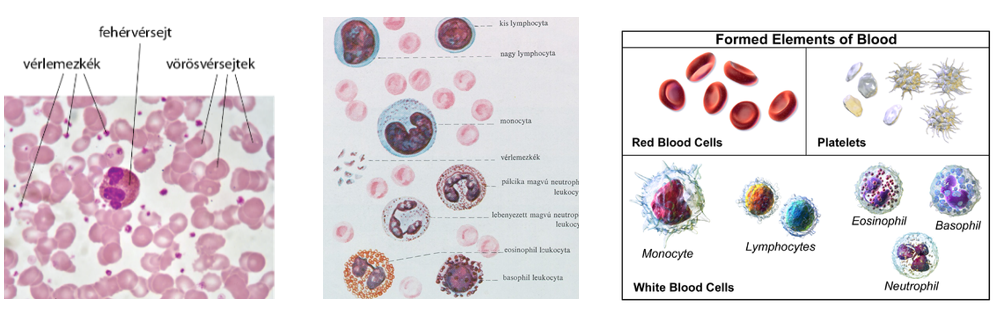 |
A fehérvérsejtek lásd még részletesen 4.8.5.1. fejezet
A fehérvérsejtek - szemben a vér alakos elemeinek másik két csoportjával - sejtmaggal rendelkező, teljes értékű sejtek. Számuk a szervezet állapotától függően széles határok között változhat, általában 4000 - 10 000/mm3, fertőzés, gyulladás esetén számuk jelentősen emelkedhet. Élettartalmuk néhány naptól akár évtizedekig is terjedhet.
A fehérvérsejtek az immunitásban játszanak szerepet, felépítésük és további szerepük alapján a sejteket 3 csoportba osztjuk.
- Granulociták
- Limfociták
- Monociták
A granulociták
Nevüket a sejtplazmában megtalálható nagy mennyiségű szemcséről (granulum) kapták, melyek antibakteriális anyagokat tárolnak. A granulociták amőboid mozgásra képesek, átlépnek a hajszálerek falán, a szövetek közötti határfelületeken és a fertőzés irányába vándorolnak kemotaxissal.
A kórokozókat bekebelezik – kis falósejtek - és a sejtekben levő szemcsék segítségével elpusztítják őket. Amikor a granulociták a kórokozókkal kapcsolatba kerülnek, belőlük is sok elpusztul. A fertőzött sebek körül keletkező genny az ilyenkor elhalt nagy tömegű granulocitától, kórokozótól és szétesett szövetelemektől származik.
Viszonylag kis méretük és fagocitáló képességük miatt mikrofágoknak is nevezzük őket.
A granulociták minden típusa – festődés alapján: eozinofil, bazofil, neutrofil - a vörös csontvelőben termelődik.
A limfociták (nyiroksejtek)
Elnevezésük onnan származik, hogy a sejtek képződése részben vagy egészben, ill. elhelyezkedésük a nyirokrendszerhez kapcsolható
A sejteknek érési helyük és funkciójuk alapján két nagy csoportjuk van.
1. Természetes ölősejtek (NK).
2. Kis limfociták, melyeknek további 2 típusa ismert:
- B limfociták, melyek a vörös csontvelőben termelődnek és az ún. szerzett (adaptív) humorális immunválasz kialakításában vesznek részt.
- T limfociták, amelyek szintén a vörös csontvelőből származnak, azonban a sejtek a fejlődésük során kivándorolnak a tímuszba (csecsemőmirigy) és érésüket ott fejezik be.
A T-sejtek az ún. szerzett sejtes, azaz celluláris immunválaszért felelősek.
A monociták – makrofágok – antigénbemutató sejtek
A monociták nagyméretű, fagocitáló, vándorló sejtek, azonban ezeket a képességüket nem a vérben fejtik ki, hanem átlépve a hajszálerek falán a szövetekben makrofágokká alakulva végzik működésüket.
Funkciójuk alapján hívjuk őket antigénbemutató sejteknek is, mivel bizonyos T-limfociták számára ők teszik lehetővé az idegen anyagok (antigének) felismerését.
A granulocitákat és a makrofágokat szokták falósejteknek is nevezni.
A vérkenet elemzésével kimutathatók
- különféle fertőzések (megnövekedett fehérvérsejtszám),
- egyes vérképzőszervi betegségek (vérsejtek számának megváltozása),
- vérszegénység (vörösvértestek számának csökkenése),
- véralvadási zavarokkal kapcsolatos problémák, mint pl. vérlemezke rendellenességek (kóros vérlemezkeszám),
- gyulladásos folyamatok (fehérvérsejtek száma, típusai alapján).
A fehérvérsejtek
- számának növekedése, egyes típusai arányának megváltozása, valamilyen fertőzésre, gyulladásra, esetleg leukémiára utal.
- Csökkent fehérvérsejtszám autoimmun betegségben, csontvelői elégtelenségben szenvedőknél fordulhat elő, ill. kemoterápia vagy sugárterápia mellékhatásaként alakulhat ki.
A vérlemezkék
- számának csökkenése kóros vérzésre, illetve vérzékenységre, leukémiára,
- megnövekedett számuk pedig fokozott vérrögképződés lehetőségére utal.
Magyarázza a szövetnedv áramlási mechanizmusát a vérnyomás és a plazmafehérjék ozmotikus nyomásának viszonya alapján.
A szövetek alapállományát képező szövetnedv a hajszálerek területén jön létre, mivel a vérerek közül kizárólag a kapillárisok fala átjárható a víz és a benne oldott kis molekulájú anyagok számára. A kapillárisok fala szűrőként működik, féligáteresztő, mivel átbocsátja a vizet, az ionokat, kisebb szerves molekulákat, pl. glükózt, aminosavakat, azonban visszatartja a nagyobb méretű vérfehérjéket és az alakos elemeket. A szűrés során lényegében fehérjementes vérplazma (szövetközti folyadék) jön létre.
A hajszálerek területén a folyadék mozgásának az irányát ellentétes hatások eredője szabja meg.
- Egyrészt az erekben uralkodó vérnyomás, amely a vérplazmát kifelé préseli,
- másrészt ennek ellene hat a plazmafehérjék kolloidozmotikus nyomása – ozmotikus szívóereje.
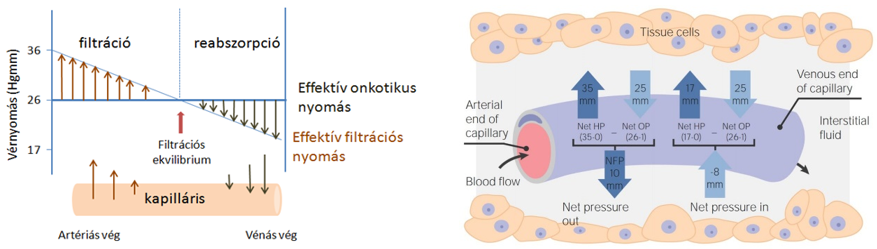 |
- A kapillárisok artériás szakaszán a vérnyomás nagyobb a kolloidozmotikus nyomásnál, ezért a vérplazma fehérjementes szűrlete kilép az érpályából (reverz ozmózis). A kapillárisokban a vénás oldal felé haladva a folyadék kilépése következtében a vér nyomása csökken.
- Amikor a vérnyomás egyenlővé válik a kolloidozmotikus nyomással, a folyadék kiszűrődése megszűnik.
- A kapillárisok vénás szakaszán a vérnyomás tovább csökken és a kolloidozmotikus nyomás alá esik. Ennek következtében a vénás szakaszon a vérfehérjék kolloidozmotikus szívóhatásának köszönhetően a szövetnedv egy része visszalép az érpályába, elszállítva a szövetekben keletkezett bomlásterméket. A szövetnedv oldott vérfehérjéket nem nagyon tartalmaz, ezért kolloidozmotikus nyomása alacsony.
Elemezzen a szív működésével kapcsolatos élettani kísérletet.
Emelt szintű érettségi feladat
Azt a feltételezést, miszerint az idegroston haladó ingerület hatása valamilyen vegyület közvetítésével jut át az izomra, 1921-ben az ausztriai születésű kutató, Otto Loewi bizonyította.
A gerincesek szívműködését két ellentétes hatású beidegzés szabályozza. Loewi mindkét ideget érintetlenül hagyva preparált ki egy, a testből eltávolított békaszívet.
Amikor a gátló ideget (a bolygóideget) ingerelte, a szívpreparátumból valamilyen anyag szabadult fel, melyet a szívet körülvevő sóoldatban felfogott, és egy másik, hasonlóan kipreparált békaszívet kezelt vele.
A második szív működése ekkor ugyanúgy lassult, mint az első szívé, noha az oda vezető gátló idegét nem ingerelte. Néhány éven belül Loewinek sikerült azonosítani a hatást kiváltó anyagot és bizonyította, hogy a gátló idegvégződésekből acetilkolin szabadult fel.
A kísérletből arra következtetett, hogy az agyideg és szív kapcsolatában kémiai jelátvitel valósul meg (és nem közvetlen elektromos jelátvitel).
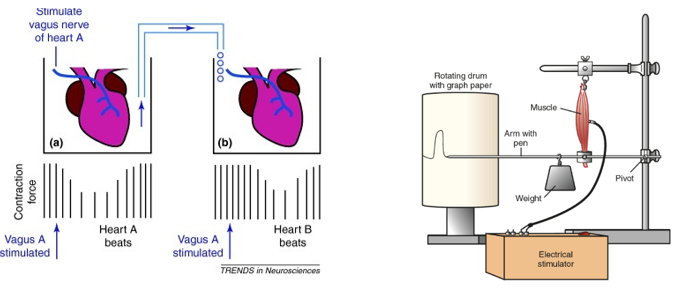 |
Béka szívének vizsgálata – izolált szívkísérlet
A kísérlet célja a szív önálló működésének és idegrendszeri szabályozásának megfigyelése, valamint az ingerlésre adott válaszok (pl. frekvencia, összehúzódás erőssége) tanulmányozása.
Módszer, eszközök
- Kísérleti állat: béka (pl. zöld levelibéka).
- A béka szívét óvatosan eltávolítják és Ringer-oldatban tartják (ez az oldat biztosítja az ionháztartást és életben tartja a szöveteket).
- A szívet rögzítik egy tartóra, és egy erőmérőre kapcsolják, hogy mérni lehessen az összehúzódások erejét.
- Elektromos ingerléssel vagy különféle oldatokkal (pl. adrenalin, acetilkolin) stimulálják a szívet.
Megfigyelhető jelenségek
Saját ingerképzés – automácia
- A béka szíve idegrendszeri kapcsolat nélkül is ritmikusan összehúzódik – ez bizonyítja, hogy saját ingerképző rendszerrel rendelkezik.
Ingerlésekkel kiváltott válaszok
- Elektromos impulzus hatására a szív összehúzódik.
- A túl gyors ingerlés során a szív tetanuszos válaszra nem képes, mert minden összehúzódás után következik egy elernyedés – ez a szívizom élettani sajátossága.
Neurotranszmitterek hatása
- Adrenalin (szimpatikus ingerületátvivő anyag): növeli a pulzust és az összehúzódások erősségét.
- Acetilkolin (paraszimpatikus ingerületátvivő): csökkenti a pulzusszámot.
Tervezzen kísérletet egy potenciális alvadásgátló gyógyszer hatásának vizsgálatára.
Az alvadási rendszer – belső út - állapotáról alvadási (trombin) idő mérése tájékoztat. Ez azt az időtartamot jelenti, amely alatt az üvegedénybe frissen levett vérben megjelenik az első fibrinszál. Az alvadási idő átlagos értéke 5-6 perc.
A véralvadási folyamat egy adott részéről – külső út - nyújt információt a protrombin idő. Másodpercben megadja, hogy a szöveti faktor hozzáadását követően mennyi idő telik el a vizsgált vérmintában az alvadék képződéséig. A vizsgálatra például akkor van szükség, ha a trombózisra hajlamos páciens valamilyen szájon át szedhető véralvadásgátló kezelést kap, segítségével vizsgálható, hogy a véralvadásgátló gyógyszerek megfelelőek-e a vérrög képződésének megelőzésére.
Alvadásgátló hatás vizsgálata in vitro (pl. humán vérmintán)
A kísérlet célja, hogy megállapítsuk, milyen mértékben képes a vizsgált anyag gátolni a véralvadást in vitro (laboratóriumi) körülmények között.
Cél
A vizsgált új hatóanyag (pl. „X vegyület”) véralvadásra gyakorolt hatásának meghatározása kontrollált laboratóriumi körülmények között, és összehasonlítása ismert alvadásgátlóval (pl. heparin).
Eljárás
1. Minták begyűjtése
- Önkéntes (egészséges) donortól vett friss vérminták.
2. Kezelési csoportok kialakítása
Csoport Kezelés
Kontroll Nincs kezelés (csak vér)
Pozitív kontroll Heparinnal kezelt vér
Kísérleti 1 Vegyület X – alacsony dózis
Kísérleti 2 Vegyület X – közepes dózis
Kísérleti 3 Vegyület X – magas dózis
3. Véralvadási idő mérése (koagulációs tesztek)
- Protrombin idő (PT) – a külső alvadási út vizsgálatára.
- Trombin idő (TT) – a fibrinogén → fibrin átalakulás sebessége.
Várható eredmények
- A kontrollcsoportban a normál alvadási idő mérhető.
- A pozitív kontroll (heparin) jelentősen meghosszabbítja az alvadási időt.
- A Vegyület X dózisfüggően befolyásolhatja az alvadást:
-
- Ha hatékony, az alvadási idők meghosszabbodnak.
- A PT vagy TT eredménye alapján megállapítható, hogy melyik alvadási útra hat a gyógyszer.
-
Élettani és klinikai jelentőség
Az alvadásgátlók kritikus szerepet játszanak a trombózis, embólia, szívinfarktus és stroke megelőzésében.
A kísérlet segít meghatározni:
- A gyógyszer hatékonyságát
- A terápiás dózistartományt
- A lehetséges mellékhatásokat (pl. túlzott vérzés)
4.7. A kiválasztás |
Elemezzen adatokat, grafikonokat, végezzen el megadott képlet alapján számításokat a vese működésének vizsgálatára, hogy egy adott anyag időegység alatt mekkora mértékben távozik a vérből a vesén keresztül. Tervezzen vizsgálatot a vizelet lehetséges összetevőinek kimutatására.
Egy kis elméleti bevezető a számolási feladatokhoz (nem követelmény)
Az érgomolyagok szűrési kapacitását a glomeruláris filtrációs rátával (GFR) lehet jellemezni, ami megadja az időegység alatt átszűrődő folyadék mennyiségét. Segítségével megadható egy adott anyag (marker) időegység alatti kiürülésének mértéke a plazmából. Átlagos testméreteket véve átlagértéke kb. 120 ml/min. A GFR meghatározása markervegyületek segítségével történik. A vizsgálat során a vérplazma és a vizelet elemzéséből a markerek clearance-ét (tisztulás) határozzák meg. A clearance (C) az a plazmatérfogat, amely tartalmazza az időegység alatt kiürült anyag mennyiségét, dimenziója ml/min. A leggyakrabban használt vegyületek az inulin és a kreatinin. Fontos követelmény a vizsgálatban használt anyaggal szemben, hogy 100%-a átszűrődjön, ne szívódjon vissza semmilyen mértékben, ne legyen bomlékony és ne szintetizálódjon a vesében. A megadott feltételeknek eleget tevő anyag esetében a clearance értéke megegyezik a (GFR) értékével. Az eljárás során a vérplazmában levő ismert mennyiségű anyag koncentrációját hasonlítják össze a vizeletben megjelenő mennyiségével. Ez alapján Cinulin = GFR = U.V/P
- ahol U = az adott anyag (inulin) vizeletben mért koncentrációja,
- V = időegység alatt kiválasztott vizeletmennyiség,
- P = az adott anyag (inulin) plazmában mért koncentrációja.
Számolási feladatok
Egy felnőtt ember vérkeringési rendszerébe 2,0 gramm inulint Rövid idő múlva, amikor az inulin a vérplazmában már egyenletesen eloszlott, de az érpályából még nem jutott ki, meghatározták, hogy az inulin koncentrációja a vérplazmában 0,667 gramm/dm3 volt.
Az inulin a vesében 100%-ban kiszűrődik és nem szívódik vissza. A szóban forgó vizsgálat során mérték a képződő vizelet mennyiségét és benne az inulin töménységét. Az egyénben percenként 1,2 cm3 vizelet képződött, s benne az inulin koncentrációja 72,25 gramm/dm3 volt.
Az adatok felhasználásával számolja ki, hogy a vizsgált egyénben percenként hány cm3 vérplazma haladt át a vesén!
- 1,2 cm3 = 0,0012 dm3
- 1 dm3 vizeletben van 72,25 g inulin, akkor
- 0,0012 dm3 vizeletben x = 0,0867 g inulin van.
- 0,667 gramm inulin van 1 dm3 vérplazmában (lásd fent), akkor
- 0,0867 g inulin x = 0,13 dm3 = 130 cm3 vérplazmában található.
Egy klinikán a vizsgált személy veséjének vérellátottságát szeretnék vizsgálni, ezért meg kívánják határozni a szerv vérátáramlását. Ehhez a gyakorlatban paraaminohippursavat (PAH) használnak segédanyagként, egy nagyvéna vérébe beadva. A vese ezt az anyagot meghatározott arányban választja ki, a vérplazma PAH-tartalmának átlagosan 90%-a ürül a vizelettel. A mérés során a vizsgált személy veseartériájának vérplazmájában a PAH koncentrációja 1,2 mg/cm3, vizeletében pedig 672 mg/cm3 volt.
Mennyi a vesén átáramló vér mennyisége cm3/perc egységben, ismerve, hogy a vérplazma a vér térfogatának 55%-át teszi ki, a napi vizeletmennyiség pedig 1500 cm3!
- A vizelettel ürülő PAH mennyisége: 1500 · 672 mg = 1 008 000 mg/nap.
- Ez a vesén áthaladó PAH 90%-a, a teljes áthaladó napi mennyiség: 1 008 000 : 0,9 = 1 120 000 mg/nap.
- Ez a mennyiség 1 120 000 : 1,2 = 933 333,33 cm3 vérplazmában található meg.
- percenként: 933 333,33 : 24 : 60 = 648,14 cm3/perc vérplazma halad át a vesén.
- Ennyi vérplazmát 648,14 : 0,55 = 1178,4 cm3 vér tartalmaz.
A veséken percenként átlagosan 0,67 dm3 vérplazma áramlik át. Az érgomolyagokon az átáramló vérplazmának kb. 20%-a szűrődik át. Mennyi a szűrlet napi mennyisége? Mennyi vér áramlik át percenként a veséken (a sejtes elemek aránya 44%)?
- Az átszűrődő plazma térfogata 0,67 dm3/perc x 0,2 = 0,134 dm3/perc.
- Egy nap alatt: 0,134 dm3/perc x 60perc x 24h = 193 dm3 szűrlet képződik.
- A percenként átáramló vér térfogata 0,67 dm3 / 0,56 = 1,2 dm3
Tervezzen és értelmezzen állatkísérletet a vazopresszin (ADH) vízmegtartó szerepének vizsgálatára.
A vazopresszin vízmegtartó hatásának vizsgálata patkányokon
Kísérleti alanyok
Felnőtt, egészséges, azonos nemű (pl. hím) laborpatkányok (n=24), 3 csoportba osztva.
Csoportok
Kontroll csoport (n=8)
- Normál körülmények között tartva.
- Injekcióban fiziológiás sóoldatot kapnak (placebo).
ADH csoport (n=8)
- Injekcióban szintetikus ADH-t (vazopresszin) kapnak.
ADH-gátló csoport (n=8)
- Injekcióban ADH-antagonistát (pl. tolvaptán, mely ADH-receptor antagonista) kapnak, amely gátolja az ADH hatását.
Kísérlet menete
- Minden csoport standard mennyiségű vizet és tápot kap.
- Injekciókat (ADH, placebo vagy ADH-antagonista) adott időpontban adják (pl. reggel).
- 24 órán keresztül mért paraméterek:
-
- Elfogyasztott vízmennyiség (ml)
- Kiválasztott vizelet mennyisége (ml)
- Vizelet koncentrációja (ozmolalitás)
Eredmények
| Csoport | Várható vizeletmennyiség | Vizelet koncentráció | Értelmezés |
| Kontroll | Közepes | Normál | Normál ADH működés |
| ADH (vazopresszin) | Alacsony | Magas | Fokozott vízvisszatartás, ADH hatása érvényesül |
| ADH-gátló | Magas | Alacsony | ADH hatása blokkolva, vízvesztés fokozott |
Értelmezés
- Az ADH a vesében a gyűjtőcsatornák vízáteresztő képességét növeli → több víz szívódik vissza → kevesebb, koncentráltabb vizelet jön létre.
- Az ADH-antagonisták gátolják ezt a folyamatot → több víz ürül ki → hígabb, nagyobb mennyiségű vizelet keletkezik.
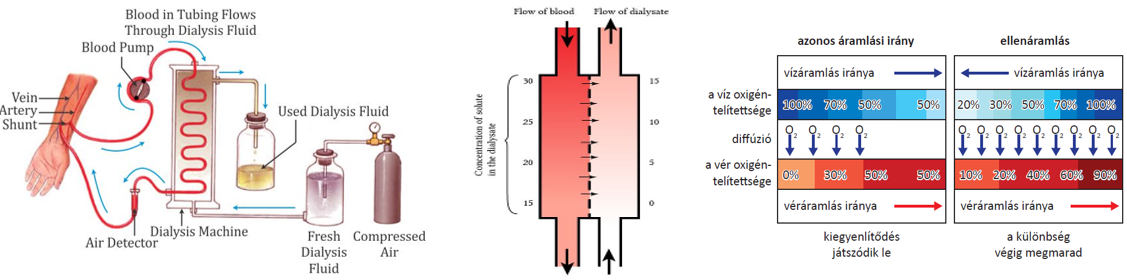 |
Alkalmazza az ellenáramlás elvét a művesekezelés folyamatának magyarázatában.
A veseelégtelenségben szenvedő betegeknél alkalmazott művesekezelés az ozmózis jelenségén alapszik. Az eljárást hemodialízisnek nevezzük.
A beteg vérét egy szemipermeábilis csövön keresztül dializálógépbe vezetik, ahol a toxikus anyagokban gazdag vért tartalmazó csövek körül ún. steril dializáló folyadék áramlik. A berendezésben a mérgező salakanyagok a koncentráció-különbségnek megfelelően kijutnak a vérből a féligáteresztő hártyán keresztül a dializáló oldatba, ugyanakkor a plazmafehérjék és a vér alakos elemei a vérben maradnak. Az ilyen módon megtisztított vért visszavezetik a testbe.
A dializáló berendezés hatékony működése is az ellenáram elvén alapul, a csőben áramló vérrel ellentétes irányban mozog a dializáló folyadék, ami a cső teljes hosszában fenntartja a káros anyagok koncentráció-különbségét (koncentrációgradiensét), és emiatt egyenletes diffúziósebességet tart fenn a két folyadéktér között.
Amennyiben a folyadékok azonos irányban áramlanának, akkor anyagtartalmuk egyre jobban kiegyenlítődne.
A halak külső gázcseréje hasonlóan ellenáramlás elvén alapszik.
4.8. A szabályozás |
Értelmezzen a drogok, agonisták, antagonisták biológiai hatásának a bemutatására vonatkozó kísérletet vagy tanulmányt.
A drog általában olyan hatóanyag, ami megváltoztatja a szervezet működését. A tudatmódosító vagy pszichoaktív szerek azok a természetes vagy mesterséges drogok, amelyek a központi idegrendszer működését változtatják meg (lásd részletesen még 4.8.2. fejezet).
A drogok ill. egyes mérgek megváltoztatják – serkentik vagy gátolják - a szinapszis működését
- a neurotranszmitterek felszabadulásának, lebomlásának, visszavételének befolyásolásával
- vagy módosíthatják a receptorok működését.
1. Hatásukat tehát kifejthetik a jelátvivő anyag
- felszabadulásának fokozása (serkentők, pl. amfetamin), ill. gátlása (lassító drogok, pl. opiátok),
- visszavételének gátlása (serkentők, pl. kokain),
- lebontását végző enzimek hatásának módosítása útján.
Tehát a gyorsító – serkentő - drogok
- gátolják az átvivőanyagok visszavételét (kokain a dopaminét),
- lebomlását,
- serkentik az ürülésüket (amfetamin). Az extasy pl. növeli a szerotonin, dopamin és a noradrenalin felszabadulását.
A lassító – depresszáns - drogok, nyugtatók
- gátolják a transzmitterek ürülését (opiátok: morfin és társai),
- vagy GABA-receptorokat serkentenek, melyek Cl--ioncsatornák, hiperpolarizációt okoznak (nyugtatók pl. benzodiazepinek (pl. xanax), a barbiturátok, ill. az alkohol).
2. Receptormódosítás - a receptorokra ható agonista-antagonista hatás változása - révén.
Agonistának nevezzük azokat a hatóanyagokat, ún. ligandumokat, amelyek receptorokhoz képesek kötődni aktiválva működésüket. Ezek lehetnek
- természetes hormonok, neurotranszmitterek,
- de lehetnek kívülről a szervezetbe juttatott vegyületek, mérgek, drogok.
Tehát a drogok egyes fajtái is az idegsejtek receptoraihoz kötődve fejtik ki a hatásukat.
- A gátló morfin K+-csatornák nyitása révén hiperpolarizációt eredményez, ami gátolja a serkentő átvivőanyagok ürülését.
- A THC (tetrahydrocannabinol) a kannabinoid-receptorokat aktiválja (CB1-receptorok), amelyek szerepet játszanak a hangulat, a fájdalomérzet, az étvágy és más funkciók szabályozásában.
- A nikotin serkentő hatását úgy éri el, hogy a központi idegrendszerben az idegsejtek acetilkolin-receptoraihoz kötődik, aktiválva a működésüket (hatására a vérnyomás, szívfrekvencia nő, az idegsejtek fokozzák dopamin leadásukat, ami örömérzet és így a függőség kialakulásában játszik szerepet).
- Az alkohol két ingerületátvivő anyag receptorához is kötődik, megváltoztatva azok működését.
- A gátló GABA-receptorokat serkenti,
- a serkentő glutamát-receptorát gátolja, miáltal összességében pl. szorongásoldó hatást ér el.
Az alkoholfüggést azzal magyarázzák, hogy az alkoholbetegeknél csökken a GABA-receptorok száma, ugyanakkor megnő a glutamát-receptoroké, melyek gátlásához egyre több alkohol bevitelére van szükség, az alkohol hiánya elvonási tünetekhez vezet.
Antagonisták azok a vegyületek, amelyek valamilyen módon gátolják az agonisták hatását. A receptorokhoz kötődve inaktiválhatják azokat, ezáltal gátló hatást válthatnak ki.
- A koffein a gátló adenozin átvivőanyag receptorához kötődve meggátolja annak hatását. Az adenozin természetes körülmények között gátolja a dopamin felszabadulását, tehát a koffein a gátlás gátlásával – a dopamin leadás növelésével - élénkítő hatást ér el.
- Az idegmérgek egyes csoportjai antagonista hatásuk révén szinapszisok működését befolyásolják. Ilyen pl. a curare, amely dél-amerikai indiánok nyílmérge, mely különböző fák kérgéből készül. A curare az izmok acetilkolin-receptoraihoz kapcsolódva megakadályozza az ideg-izom szinapszis működését, így az izom mozgásképtelenné válik.
A rendszeres droghasználat során függőség – addikció - alakul ki, azaz a szer csökkent bevitele vagy abszolút hiánya megvonásos tünetekkel jár, mivel a negatív visszacsatolás miatt a droghoz hasonló hatású, ugyanahhoz a receptorhoz kötődő, a központi idegrendszerben termelődő természetes transzmitterek termelődése csökken.
További következménye a különféle idegrendszerre ható anyagok – drogok - folyamatos alkalmazásának a tolerancia kialakulása, melynek során a szer tartós használata miatt annak hatása egyre csökken, s az anyag ugyan azt a hatást csak egyre nagyobb dózisban képes kifejteni. A tolerancia hátterében egyrészt a drogot lebontó enzimek aktivitásának növekedése, másrészt a receptorok számának a csökkenése áll.
Kísérlet
A drogok, agonisták és antagonisták hatásának vizsgálata simaizom-preparátumon
Cél
Megfigyelni, hogyan hat egy agonista (pl. acetilkolin), egy antagonista (pl. atropin), illetve ezek kombinációja egy izolált simaizom (pl. béka vagy patkány bélizom) összehúzódására.
Anyagok és eszközök
- Izolált béka/patkány bélizom-preparátum
- Fiziológiás oldat
- Erőmérő (izomösszehúzódások mérésére)
- Agonista: pl. acetilkolin (ACh)
- Antagonista: pl. atropin
- Időzítő, regisztráló eszköz (grafikonrajzoló szoftver vagy mechanikus eszköz)
Módszer
- Az izolált bélizmot az fiziológiás oldatba helyezzük, és egy erőmérőhöz rögzítjük.
- Alapértéket rögzítünk drog nélkül – ez az izom nyugalmi állapota.
- Hozzáadunk acetilkolint (agonista) → figyeljük és mérjük az izom összehúzódását.
- Öblítjük a rendszert, visszatérünk a kiindulási állapothoz.
- Hozzáadunk atropint (antagonista) → nem várunk izomválaszt.
- Ismételten acetilkolint adunk atropin jelenlétében → az izomreakció csökken vagy elmarad.
- Dokumentáljuk az erőgörbéket minden lépésnél.
Eredmények értelmezése
- Acetilkolin önmagában → izomkontrakció, mivel agonistaként serkenti a kolinerg receptorokat.
- Atropin önmagában → nincs kontrakció, mivel antagonistaként blokkolja a receptorokat, de nem aktiválja őket.
- Acetilkolin + atropin együtt → csökkent vagy teljesen gátolt válasz, mert az atropin elfoglalja a receptorokat, így az ACh nem tud hatni.
Következtetés
- Az agonisták aktiválják a receptorokat → biológiai válasz (pl. izomösszehúzódás).
- Az antagonisták blokkolják a receptorokat → meggátolják a természetes agonista hatását.
- A drogok hatása tehát nemcsak a jelenlétüktől, hanem a receptorhoz való kötődésüktől is függ.
Elemezze a gerincvelői keresztmetszetet bemutató rajzolt ábrát vagy szövettani metszeti képet (felépítés és funkció, szürke-és fehérállomány, kötegek, szarvak, központi csatorna, gyökerek, gerincvelői idegek).
Váltson ki térdreflexet, és magyarázza funkcióját.
Az idegsejteket nemcsak alakjuk, hanem feladatuk alapján is csoportosíthatjuk:
- Az érzőidegsejtek a receptoroktól a központ felé továbbítják az ingerületet, mindig a környéki idegrendszerhez tartoznak, ilyenek vannak pl.
- a retinában, a csigában (bipoláris idegsejtek),
- a csigolyaközti dúcokban (pszeudouniporáris sejtek).
- A mozgatóidegsejtek a végrehajtószerv (izmok, mirigyek) felé közvetítik a központok utasításait.
- A köztes idegsejtek (interneuronok) összekapcsolják az érző- és a mozgatóidegsejteket. A legtöbb idegsejtünk ilyen.
Reflexműködés
A reflexműködés a környezet – külső, belső – ingereire szervezett válaszreakció.
A reflexműködés kivitelezésében szerepet játszó anatómiai képletek reflexívet alkotnak, melynek részei:
- a receptor, ami felfogja az ingereket és ingerületté alakítja,
- az érzőidegsejt, ami továbbítja a központ felé az ingerületet (akciós potenciált),
- az interneuron, ami összekapcsolja az érző- és a mozgatósejtet,
- a motoros neuron, aminek az axonja a végrehajtószervben – pl. izomban - végződik.
Magasabb rendű szervezetekben – emlősökben - reflexkörök alakulnak ki, ebben az esetben a receptor-központ, ill. a végrehajtószerv-központ közti kapcsolat kétirányú, ilyenkor
- a központ szabályozza a receptor ingerküszöbét, ill.
- a végrehajtószerv az állapotáról visszajelzést küld a központba.
Különbséget teszünk a szomatikus és a vegetatív reflexkörök lefutása között.
 |
A gerincvelő keresztmetszete
Kétféle részt különböztetünk meg.
- Kívül fehérállomány,
- belül szürkeállomány.
Szürkeállomány
- Pillangó alakú.
- Neuronok sejttestei alkotják.
- Közepén van a központi csatorna.
Részei:
- Hátsó szarv: interneuronok.
- Oldalsó szarv: központi vegetatív interneuronok.
- Mellső szarv: szomatikus motoros neuronok.
Fehérállomány
- Pályák alkotják.
- Elülső köteg: leszálló – mozgató - pályák.
- Hátulsó köteg: felszálló – érző - pályák.
- Oldalsó köteg: vegyesen.
A gerincvelő szelvényezettségét a belőle kilépő, ill. belépő idegrostok, az ún. gerincvelői gyökerek okozzák.
Mellső gyökér: idegrostjai a mellső szarvi mozgatóneuronokból és az oldalsó szarvi központi vegetatív neuronokból erednek, így kizárólag mozgatórostokat tartalmaznak.
Hátsó gyökér: külső részén egy duzzanatot, az ún. csigolyaközti dúcot találjuk, benne pszeudounipoláris érzőidegsejteket, melyek centrális nyúlványai hozzák létre a hátsó gyökeret. Így a hátsó gyökérben kizárólag érzőrostok – szomatikus és vegetatív- futnak a receptorok felől a központok felé.
A két gyökér a gerinccsatornából való kilépés előtt gerincvelői ideggé egyesül, amely már kevert, érző- és mozgatórostokat egyaránt tartalmaz (31 pár). A gerincvelői idegek szelvényezetten hagyják el a gerincvelőt, s rostjaik szelvényesen hálózzák be a testet.
A szelvényezettség a dermatómák kialakulásában is megnyilvánul. A dermatóma a bőr, egy adott gerincvelői ideggel beidegzett területe.
 |
Izomeredetű szomatikus reflex: feszítő reflex vagy nyújtási reflex (monoszinaptikus)
Ilyen a térd-, ill. a könyökreflex. Amennyiben reflexkalapáccsal a combfeszítő izom térdkalács alatti inára ütést mérünk, a lábszár kilendül. Az ütés következtében – mivel az inak nem nyújthatók - a combfeszítő izom megnyúlik, aminek hatására - a reflexkörnek köszönhetően - az izom aktív összehúzódással válaszol. A reflex vizsgálata tájékoztatást nyújt a gerincvelő állapotáról.
A receptor az izomorsó,
- amely a combfeszítő izomban található,
- az izomrostokra csavarodott idegvégződés,
- mechanoreceptor, mivel adekvát ingere az izom megnyúlása (inakban az ínorsó van).
A reflexív lefutása
A bevezetőszárt az érzőidegsejt centrális nyúlványa képezi, mely belépve a gerincvelőbe 3 felé ágazik:
- felszállópályán a hátsó kötegben az agyvelőbe tart, ill.
- az azonos szelvényben elhelyezkedő mellső szarvi mozgatóneuronhoz kapcsolódik, melynek rostjai az elülső gyökéren keresztül érik el ugyanazt az izmot (combfeszítő izmot), kiváltva annak összehúzódását (a láb kilendül),
- továbbá az alatta lévő gerincvelői szelvényben található hátsó szarvi gátló interneuronnal képez szinapszist, amely a hajlítóizom beidegzését végző mozgatóneuront gátolva, a hajlítóizom elernyedését váltja ki (elkerülendő az izomszakadást).
A nyújtási reflex feladata:
- Védi az izmot a túlzott passzív megnyúlástól, így az izomszakadástól, hiszen nyújtásra az izom összehúzódással reagál.
- Jelentős a testtartásban, azaz az antigravitációs izmok optimális feszítettségi állapotának beállításában, ezáltal testtartási reflexek alapját képezi.
- Alapvető az izomtónus fenntartásában (izomtónus: az izmok állandó kismértékű feszülése, a testtartásban alapvető a jelentősége).
 |
Értelmezzen gerincvelő-sérülési ábrákat és tudja megjósolni az egyes sérülések következményeit.
Gerincvelő felépítése – amit az ábrán látunk
A gerincvelőt a csigolyák és az idegpályák alapján különböző szakaszokra osztjuk. Egy ábrán általában az alábbiakat jelölik:
| Szint | Megnevezés | Mit idegez be? |
| C1–C8 | Nyaki (cervicalis) | Karok, nyak, rekeszizom |
| T1–T12 | Mellkasi (thoracalis) | Törzs, mellizmok, bordaközi izmok, hasizmok, bélcső felső szakaszának, mellkasi szerveknek szimpatikus beidegzése |
| L1–L5 | Ágyéki (lumbalis) | Alsó végtagok (pl. combfeszítők), bélcső alsó szakaszának, húgy-ivari szerveknek szimpatikus beidegzése |
| S1–S5 | Keresztcsonti (sacralis) | Alsó végtagok (pl. combhajlítók), medence izmai, bélcső alsó szakasza, húgy-ivari szervek paraszimpatikus beidegzése |
Az ábrákon gyakran látszanak sérülési szintek, például „C6 sérülés”, illetve a teljes vagy részleges átmetsződés.
Sérülések típusai és következményei
| Sérülés típusa | Jelentés | Következmény |
| Teljes (komplett) | A gerincvelő teljes keresztmetszete sérült | Teljes bénulás és érzéskiesés a sérülés alatt |
| Részleges (inkomplett) | A gerincvelő egy része sérült | Részleges mozgás/érzékelés megmaradhat |
Sérülés szintje szerinti következmények – példák
| Sérülés szintje | Lehetséges következmények |
| C1–C4 | Teljes négyvégtagbénulás (tetraplégia), légzőizom bénulás → gépi lélegeztetés szükséges lehet. |
| C5–C6 | Részleges karfunkciók, de lábak bénultak. Kézmozgás korlátozott. |
| C7–C8 | Kar- és kézmozgások többnyire megtartottak, alsó végtagok bénultak. |
| T1–T6 | Alsó végtagbénulás (paraplégiának hívják), felsőtest funkciók épek. |
| T7–T12 | Stabil törzs, ülés lehetséges, de alsó végtagok mozgásképtelenek. |
| L1–L5 | Egyes lábmozgások lehetségesek, de gyenge járás vagy segédeszköz szükséges. |
| S1–S5 | Leginkább hólyag-, bél-, nemi funkciók érintettek. |
Ábra értelmezésének lépései
- Azonosítsd a sérülés szintjét – például „T10” azt jelenti, hogy a 10. háti csigolya magasságában történt a sérülés.
- Nézd meg, teljes vagy részleges-e a sérülés.
- Következményeket a szint alapján jósolhatjuk meg, figyelembe véve:
-
- Mozgásképtelenség (mely végtagok érintettek)
- Érzéskiesés (mely bőrfelületeken)
- Vegetatív funkciók zavara (pl. légzés, hólyagkontroll)
-
Emelt szintű biológia érettségi esszégyűjtemény
A biológia esszé nem irodalmi fogalmazás, hanem strukturált szakmai válasz.
A sikeres biológia esszé kulcsa a feladatlap tételes kérdéseinek szigorú követése, a szaknyelv pontos használata és az adatok közti összefüggések feltárása. Nem a stílus, hanem a javítókulcs szerinti kulcsszavak és tények érnek pontot, ezért a kérdések sorrendjében haladva, vázlatpontokat követve érdemes kidolgozni a témát.
A biológia esszéírás lépései
- Választás és elemzés: Az emelt szintű érettségin (9. feladat) A és B opció közül választhatsz (általában ember- vagy ökológia/növénytan). Olvasd el figyelmesen a kérdéseket, és húzd alá a kulcsszavakat.
- Vázlat készítése: A feladat tételesen felsorolja, mire kíváncsi. Ezeket a kérdéseket érdemes sorban megválaszolni, hogy ne maradhasson ki semmi, és a logikai menet biztos legyen.
- Szaknyelv használata: A javítókulcs konkrét szavakat és kifejezéseket keres. Használj pontos biológiai terminológiát, ne csak általánosságban írj.
- Struktúra: Formai kötöttség nincs, a kifejtés a kérdések mentén haladjon. A "szép fogalmazás" helyett a pontos tartalom a lényeg.
- Időbeosztás: Gyakorolj otthon, mérd az időt, és szimuláld a vizsgahelyzetet.
Legfontosabb tanácsok
- Soha ne írd meg mindkét esszét: Ha mindkettőt megoldod, csak az "A" feladatot fogják értékelni, még ha a "B" jobb is lett!
- Kipipálás: Amelyik alpontra válaszoltál, pipáld ki a feladatlapot, hogy biztosan mindegyikről írtál-e.
- Összefüggések: Ne csak tényeket sorolj, hanem magyarázd meg az ok-okozati összefüggéseket.
A biológia esszé javítási útmutatója a tartalmi pontosságra, a szakszókincs helyes használatára és az irányító kérdések megválaszolására fókuszál. A pontok az elvárt kulcsszavak, folyamatok és összefüggések meglétéért járnak, nem a terjedelemért.
A javítási-értékelési útmutató főbb szempontjai:
- Irányító kérdések követése: A pontok az irányító kérdésekre adott konkrét, részletes válaszokhoz kötöttek.
- Kulcsszavak és szakkifejezések: A biológiai folyamatok pontos megnevezése elengedhetetlen. A szinonimák elfogadottak.
- Logikai sorrend: Az események, folyamatok oksági összefüggésének bemutatása fontosabb, mint a leírás sorrendje. Ha a válasz logikus, a javítókulcsban szereplőtől eltérő sorrend is pontozható.
- Helyesírás: A helyesírási hibákért nem vonható le pont, de az értelemzavaró megfogalmazások vagy ellentmondó válaszok nem pontozhatók.
- Pontozás: Általában minden pontos, helyes szakmai elem (pipa) 1 pontot ér. A részpontszámok (fél pont) nem adhatók.
2025 október
XI.A. Mozgásérzékelés – esszé 10 pont
Esszéjét összefüggő és logikus mondatokban a következő szempontok alapján építse fel!
1. Mutassa be a belső fül szerepét a testhelyzet és a testmozgás érzékelésében! (4 pont)
2. Ismertesse az izomorsók funkcióját, a térdreflexben részt vevő idegsejtek kapcsolódását, működését! Magyarázza meg, hogy miért van szerepe ennek a reflexnek a testérző rendszer működésében, ha alapvetően ez egy gerincvelői reflex! (6 pont)
XI.B. Biológiai sokféleség és regeneráció – esszé 10 pont
Fogalmazza meg a biodiverzitással, annak megóvásával és helyreállításával kapcsolatos összefüggéseket!
Esszéjét összefüggő és logikus mondatokban a következő szempontok alapján építse fel!
1. Definiálja a biodiverzitás következő szintjeit: genetikai diverzitás, fajdiverzitás, ökológiai diverzitás! Melyik diverzitási szint értelmezhető a havasi kecskék esetén, és ennek értéke hogyan változhatott meg a B) szövegrészlet alapján? (4 pont)
B) szövegrészlet: „[…] A Délkelet-alaszkai Egyetem, illetve a kanadai Victoria Egyetem vadbiológusai 17 évig tartó kutatásuk során igazolták, hogy a valóságban rengeteg havasi kecske (2. ábra) pusztul el lavina következtében. […] A lavinák komoly hatást gyakorolnak a kecskék populációdinamikájára. Volt olyan alaszkai populáció, amelyben a meghalt állatok 65 százalékát lavina pusztította el […]. Átlagosan évente az állatok nyolc százaléka pusztul el lavinaomlás következtében, de volt olyan különösen rossz év is, amikor a mortalitás 22 százalékos volt. […] A lavinaomlásban az adatok szerint nemcsak a tapasztalatlan és gyenge gidák pusztulnak el, hanem gyakorlatilag a populációk bármely egyede. Az áldozatok között többségben vannak a legtermékenyebb életszakaszukat élő nőstények […].”
2. Tegyen egy-egy természetvédelmi javaslatot, amelyekkel az A) és a B) szövegrészletben szereplő esetekben megőrizhető, megvédhető a biodiverzitás! (2 pont)
A) szövegrészlet: „2020 januárjában, amikor a világ nagy része még mit sem sejtett a közelgő világjárványról, sok millióan figyelték a soha nem látott vadsággal tomboló ausztrál erdőtüzeket. Egész Délkelet-Ausztrália füstbe merült, a tűzoltók hasztalan próbálták megfékezni a lángokat, amelyek felmérhetetlen kiterjedésű erdőt és állatok sokaságát emésztettek el. A 2019–2020-as „fekete nyár”, ahogy elnevezték a hónapokig tartó katasztrófát, ízelítőt nyújtott abból, ami a klímaváltozás következtében vár a Föld nagy erdőségeire, legyenek azok Kaliforniában, Kanadában vagy Dél-Európában. […] A szakirodalomban számos adatot találtak arról, hogy a legkülönfélébb állatok – nemcsak emlősök, de madarak és hüllők is – rendszeresen használják a vombatok járatait erdőtüzek idején. Ezért a Charles Sturt Egyetem ökológusai elhatározták, hogy szisztematikusabban megvizsgálják a vombatüregek szerepét az erdei ökoszisztémák tűzzel szembeni ellenálló képességében. A terepi vizsgálatokat a Woomargama Nemzeti Parkban végezték, amelynek 311 négyzetkilométernyi területéből nagyjából 180 km2 megsemmisült a 2019–2020-as erdőtüzekben. Amikor egy évvel később az ökológusok elkezdték a terepi felméréseket, azt tapasztalták, hogy a növényzet már látványosan regenerálódni kezdett. […] A vombatok (1. ábra) növényevők, és bár időnként agresszívek lehetnek a territoriális fenyegetést jelentő fajtársaikkal és más állatokkal szemben, még mindig sokkal biztonságosabb a közelségükben átvészelni a tüzet, mint egy ragadozó társaságában. […]”
3. Az A) szövegrészletben leírt erdőtüzeket követően a területek fokozatos regenerációja figyelhető meg. Fogalmazza meg, hogyan változik a regenerálódás során az életközösségekben a K-stratégiájú populációk aránya, a biomassza értéke, a populációs kölcsönhatások száma és a gyomnövényfajok száma! Indokolja, hogy miért adott irányba változnak ezek a tényezők! (4 pont)
Javítási-értékelési útmutató
Mozgásérzékelés – esszé 10 pont
Minden helyes válasz 1 pont.
1.
- A félkörös ívjáratokban a folyadék elmozdulása
- a fej gyorsuló mozgását / szöggyorsulását,
- a tömlőcskékben és a zsákocskákban levő receptorok (szőrsejtek) segítségével
- a fej statikus helyzetét / egyenletes lineáris gyorsulását érzékeli a szervezet.
2.
- Az izomorsók a harántcsíkolt (váz)izmok megnyúlását érzékelik.
- A combfeszítő izom megnyúlásakor az izomorsó ingerületi állapota átterjed az érzőidegsejtre.
- A gerincvelőben az érzőidegsejt axonja kettéágazik. Egyik nyúlványa a mozgatóidegsejten keresztül összehúzódásra készteti a combfeszítő izmot,
- másik nyúlványa a combhajlító izom összehúzódását gátolja.
- A reflexív egyik leágazása a testrész helyzetéről információt szállít
- a nagyagyi érzőkéregbe / a nagyagyba / az (elsődleges) érzőkéregbe.
Biológiai sokféleség és regeneráció – esszé 10 pont
1.
- A genetikai diverzitás egy adott populációban jelenlévő allélok számával és azok gyakoriságával / az allélösszetétel változatosságával jellemezhető. 1 pont
- A fajdiverzitás azt írja le, hogy egy adott életközösségben hány faj él, és a jelenlévő fajoknak mekkora a relatív gyakorisága (egyedszámarányosan vizsgálja a fajszámot). 1 pont
- Az ökológiai diverzitás megmutatja az adott életközösségben fellelhető ökológiai szerepek / populációs kapcsolatok sokszínűségét / összetettségét. 1 pont
- A havasi kecske populációi esetén a genetikai diverzitás értelmezhető csak, melynek értéke csökken a lavinák miatti egyedpusztulás miatt. 1 pont
Másként megfogalmazott definíciók is elfogadhatók.
2.
- Az A) szövegrészlet alapján: fajok visszatelepítésével / mesterségesen létrehozott föld alatti üregekkel / a talaj tápanyagtartalmának helyreállításával segíthető a helyreállás. 1 pont
- A B) szövegrészlet alapján: akár helybeni védelemmel, például lavina elleni védettséget biztosító mesterséges búvóhelyek létrehozásával / akár egyedek védett helyekre, rezervátumokba telepítésével megvédhető a populáció a kihalástól. 1 pont
Bármely más helyes természetvédelmi módszer elfogadható.
3.
- A regeneráció során a K-stratégisták aránya növekszik, mivel a környezethez stabil létszámban alkalmazkodó populációk jönnek létre a szukcesszió során (a klimaxtársulás felé haladva). 1 pont
- A biomassza értéke szintén növekvő tendenciát mutat, mivel egyre nő az egyedszám (ezáltal az adott területen jelenlévő élővilág össztömege is). 1 pont
- A populációs kölcsönhatások száma szintén növekszik a szukcesszió előrehaladtával, mivel a megjelenő új fajok miatt egyre komplexebb kapcsolatrendszer jön létre. 1 pont
- A gyomnövényfajok száma viszont csökkenni fog, ugyanis a gyomok rosszul viselik a kompetíciót (a náluk erősebb versengők kiszorítják őket a területről). 1 pont
Más helyes indoklás, érvelés is elfogadható.
2025 május
IX.A. Elemi idegjelenségek – esszé 10 pont
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra! Esszéjét a 19. oldalon írhatja meg!
1. Ismertesse, hogy a ligandfüggő és a feszültségfüggő ioncsatornák az idegsejt sejthártyájának jellemzően melyik részén találhatók meg! Adja meg, hogy mi váltja ki szerkezetük megváltozását, és működésüknek köszönhetően hogyan változhat meg a membránfeszültség! (4 pont)
2. Ismertesse, hogy mely ionok milyen irányú transzportjának köszönhető a membránpotenciál növekedése, illetve csökkenése! (2 pont)
3. Foglalja össze, hogy az ingerületvezetés az idegsejt melyik területén történik analóg módon, melyik részén digitálisan, és mi módon kódolja ez a kétféle jel az inger erősségét! (4 pont)
XI. B. Hegységeink természetvédelme – esszé
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra!
1. Említsen a hegyvidéki élőhelyeket átalakító, befolyásoló természetvédelmi szempontból káros vagy előnyös emberi hatások közül négyet, és kapcsolja össze ezeket az ivóvízellátással, a bányászattal, az árvízvédelemmel és a turizmussal! (8 pont)
2. Miért előnyösebb az élőhelyek jogi védelme az egyes fajok védelmével szemben? (1 pont)
3. A klímaváltozás mely lehetséges hatása és hogyan befolyásolja a medvefül kankalin elterjedését? (1 pont)
Javítási-értékelési útmutató
Elemi idegjelenségek – esszé 10 pont
1.
- A ligandfüggő ioncsatornák a dendrit / sejttest sejthártyáján / a posztszinaptikus membránon helyezkednek el. 1 pont
- A feszültségfüggő ioncsatornák az axon eredési dombján / az axon sejthártyáján / az idegrost befűződésén / az axon végbunkóján találhatók meg. 1 pont
- A ligandfüggő ioncsatornák szerkezete a (fehérjemolekulához) kapcsolódó ligand / ingerület átvivő anyag hatására változik meg, és ennek következtében helyi áram / hipo-, illetve hiperpolarizáció alakul ki. 1 pont
- A feszültségfüggő ioncsatornák szerkezete egy adott membránpotenciál / küszöbpotenciál kialakulása esetén változik meg, és ennek hatására (tovaterjedő) akciós potenciál alakul ki. 1 pont
2.
- A membránpotenciál csökkenése / hipopolarizációja a Na+ felvétele miatt alakul ki. 1 pont
- A membránpotenciál növekedése / hiperpolarizációja a Cl– felvétele / K+ leadása miatt alakul ki. 1 pont
3.
- Analóg módon a sejttesten / dendriteken / csupasz axonokon 1 pont
- digitálisan az (velőshüvelyes) axonon / idegroston történik az ingerületvezetés / kémiai szinpaszisban (a „minden vagy semmi” törvény alapján). 1 pont
- Az analóg jel a de- / hipo- / hiperpolarizáció / a membránpotenciálváltozás nagyságán keresztül kódolja az inger erősségét / nagyságát. 1 pont
- A digitális jel az akciós potenciálok frekvenciáján / időbeli gyakoriságán keresztül kódolja az inger erősségét / nagyságát. 1 pont
Hegységeink természetvédelme – esszé 10 pont
1. Négy fontos emberi hatás megfogalmazása, pl.:
- erdőirtások, fakitermelés: irtásrétek, kaszálók, települések megjelenése 1 pont
- erdészeti kezelés: kisebb diverzitású, egységesen kezelt állományok megjelenése 1 pont
- sziklagyepek helyén: települések, nyaralók, bányák külszíni fejtése, tájidegen fajok megjelenése 1 pont
- A bányászat (külszíni fejtések) főként szilagyepek helyén 1 pont
Más jó példák is elfogadhatók. A vizsgázó abban az esetben kapja meg az 1+1+1+1 pontot, ha mindegyik megadott szempontot figyelembe véve elemezte a problémát.
A vizsgázó által megadott négy (előnyös és/vagy káros) hatás elemzése a megadott szempontok szerint, pl.:
- Az erdős területek csökkenése fokozódó eróziót, árvízveszélyt jelent, / de az újraerdősítés / környezetkímélő gazdálkodás csökkenti ezt a veszélyt. 1 pont
- A települések, bányák, szeméttelepek stb. növelik a (karszt)vízszennyezés kockázatát, / de a csatornázás stb. csökkenti ezt a veszélyt. 1 pont
- A turizmus fokozhatja a környezeti terhelést (taposás, hegyi motorozás), / de a falusi turizmus előmozdíthatja a természetközeli gazdálkodást. 1 pont
- A bányászat csökkenti a fajok termőterületét / degradálja élőhelyét 1 pont
Négy összefüggés elemzése, mely a megadott témákat érinti.
2. Az élőhelyek / területek védelme jobban biztosítja az egyes fajok fennmaradását, mert a létükhöz szükséges környezeti tényezőket / kölcsönhatásokat fenntartja. 1 pont
3. A felmelegedés / egyenetlenné váló csapadékeloszlás / csökkenő páratartalom veszélyeztetheti a hűvös klímát kedvelő (reliktum) fajok, így a medvefül kankalin fennmaradását is. 1 pont
2024 október
IX.A. A máj sokrétű funkciói – esszé 10 pont
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra!
1. Az epesavak mely kémiai jellemzője biztosítja az emulgeáló funkciót, és mely egyéb feladata van még az epesavaknak? Miért előnyös az emésztés szempontjából az emulgeálás? (3 pont)
2. Mely, a vizeletbe kerülő vegyületet képes előállítani a máj az aminosavak lebontása során? Mely aminosavakat nem képes más aminosavakból előállítani? (2 pont)
3. Mely szénhidrát formájában raktározza a máj a glükóz feleslegét? Elsősorban mely hormon szabályozza a szintézis mértékét? (2 pont)
4. A hemoglobin lebontását is a máj végzi. Hova kerülnek és hogyan ürülnek a szervezetből a hemoglobin lebontási termékei abban az esetben, ha a máj egészséges, és mi történik velük, ha a máj súlyosan károsodott? Mely látható tünet jelzi ezt? (3 pont)
IX.B. Tűrőképesség és biológiai indikáció – esszé
Esszéjében az alábbi szempontokra térjen ki!
1. Határozza meg a tűrőképesség fogalmát! (2 pont)
2. Egy tűrőképességi görbe rajzát elemezve adja meg annak lehetséges dimenzióit és jellemző szakaszait! (5 pont)
3. Értelmezze az indikátor szervezet fogalmát, hozzon példát indikációra, megadva azt, hogy a vizsgált példában mi, milyen módon s mit indikál! (3 pont)
Javítási-értékelési útmutató
A máj sokrétű funkciói – esszé 10 pont
1.
- Az epesavak kettős oldékonyságú (amfipatikus) molekulák / képesek poláris és apoláris anyagokban is oldódni, így tudják emulgeálni a zsírokat. 1 pont
- További funkciójuk, hogy aktiválják a lipázokat (zsírbontó enzimek) / stabilizálják a lipidemulziót. 1 pont
- Az emulgeálás révén a lipázok nagyobb felületen tudnak hozzáférni a lipidekhez, így hatékonyabban tudják azokat lebontani. 1 pont
2.
- A máj az aminosavak lebontása során karbamidot hoz létre, 1 pont
- az esszenciális aminosavat nem tudja állítani (megfelelő mennyiségben). 1 pont
3.
- A glükózt glikogén formájában raktározza 1 pont
- ennek mértékét elsősorban az inzulin / glukagon szintje szabályozza. 1 pont
4.
- A hemoglobin lebontási termékeit (biliverdin, bilirubin) a máj bejuttatja az epébe / vékonybélbe epefestékként, azok a széklettel / vizelettel ürülnek. 1 pont
- Ha a máj súlyosan károsodott, akkor ezek a lebontási termékek felhalmozódnak a vérben / bejutnak a bőrbe és a szem ínhártyájába, 1 pont
- emiatt a májelégtelenség sárgasághoz vezet. (Hasi ödémát is okozhat.) 1 pont
A májelégtelenség más helytálló következménye is elfogadható.
Tűrőképesség és biológiai indikáció - esszé 10 pont
1.
- Tűrőképesség a környezeti tényezők értékeinek azon tartománya, 1 pont
- amelyen belül az adott faj populációja előfordulhat / amit a faj populációja elvisel 1 pont
2.
Egy toleranciagörbe felrajzolása után annak jellemzése:
- Az x / vízszintes tengelyen a vizsgált környezeti tényező értékeit, pl. hőmérséklet, páratartalom stb. (Bármely jó példa) 1 pont
- Az y / függőleges tengelyen a faj életképességét jellemző valamilyen mennyiséget pl. szaporodóképességet / egyedszámot / növekedési ütemet stb. tüntetjük fel. 1 pont
(Bármely jó példa)
- - minimum: éppen elviselhető alsó határ 1 pont
- - maximum: éppen elviselhető felső határ 1 pont
- - optimum: a legkedvezőbb értéktartomány. 1 pont
3.
- Indikátor szervezet: élőlény, amely jelenlétével vagy hiányával / egyedszámával / viselkedésével / gyarapodásával jelzi a (élettelen) környezet valamely tényezőjének értéktartományát. 1 pont
- Az indikátor fajok jellemzően szűktűrésűek (egy vagy néhány tényezővel szemben) 1 pont
- Pl. bizonyos zuzmók a kén-dioxid szennyezésre, ebben az esetben a zuzmók hiánya (indikáció) jelzi a kén-dioxid magas szintjét, 1 pont
Más példa is elfogadható, ha egyértelmű, hogy mely faj mi módon és mit indikál.
2024 május
IX.A. Kétivarúság és várandósság – esszé 10 pont
Írjon esszét a nemi jellegekért felelős hormonokról, a megtermékenyítésről és az ikerterhességről az alábbi vázlatpontok alapján!
1. A kromoszómális nem definiálása. (1 pont)
2. A másodlagos női, illetve hím ivari jellegek kialakításért felelős hormonok neve, termelődésük helye és időbeli jellege. Mely életszakaszban (életkorban/mettől meddig) termelődnek a női, illetve a hím ivari hormonok a születés utáni élet során? (4 pont)
3. A megtermékenyítés helye és lehetséges időpontja a női nemi ciklus során. (2 pont)
4. Az egypetéjű és kétpetéjű ikrek kialakulása, genetikai információtartalmuk összehasonlítása egymással. (3 pont)
IX.B. A zárvatermők szaporodása – esszé 10 pont
Mutassa be a zárvatermők szaporodását az alábbi szempontok alapján!
1. Az ivaros és az ivartalan szaporítás lényege, lehetséges célja és az ivartalan szaporítás két módjának megnevezése. (3 pont)
2. A termés kialakulása az ivarsejtek kialakulásától kezdődően. A virág és a termés részeinek kapcsolata. (7 pont)
Javítási-értékelési útmutató
Kétivarúság és várandósság – esszé 10 pont
1.
- A kromoszomális nem a testi sejtek ivari kromoszómakészlete, XX: nő, XY: férfi. 1 pont
2.
- Ösztrogén – petefészkek / tüszők termelik, 1 pont
- tesztoszteron – herék termelik. 1 pont
- Az ösztrogén ciklikusan, a tesztoszteron folyamatosan termelődik. 1 pont
- Az ösztrogén az ivaréréstől a menopauzáig, a tesztoszteron ivaréréstől élethossziglan termelődik. 1 pont
3.
- A megtermékenyítés a petevezetékben / a petevezeték felső harmadában, 1 pont
- a ciklus 14–15. napján/ az ovulációt követő 12–24 órában megy végbe. 1 pont
4.
- Egypetéjű ikrek: az embrió néhány sejtes állapotban kettéválik. 1 pont
- Kétpetéjű ikrek: egyszerre /időben egymáshoz közel két petesejt termékenyül meg. 1 pont
- Az egypetéjű ikrek genetikai információja azonos (ha eltekintünk a mutációktól),
a kétpetéjűeké csak annyira hasonlít, mint a különböző időpontban született testvéreké./ átlagosan felerészben hasonló. 1 pont
Más hasonló tartalmú megfogalmazás is elfogadható.
A zárvatermők szaporodása – esszé 10 pont
1.
- Az ivaros szaporítás lényege a genetikai változatosság biztosítása / két egyed genetikai állományának keveredése / nemesítés, 1 pont
- míg az ivartalan szaporítás célja, hogy a kiválasztott génállományú növényből minél többet hozzunk létre / pl. hibridek esetében az adott jelleg ne változzon. 1 pont
- két ivartalan szaporítási mód említése, pl. tőosztás / bujtás / szemzés / dugványozás / oltás 1 pont
2.
- A megtermékenyítés során az egyik hímivarsejt a petesejttel egyesülve zigótát, 1 pont
- majd csíranövényt / embriót alakít ki. 1 pont
- A másik hímivarsejt a központi sejttel egyesülve 1 pont
- a mag táplálószövetét hozza létre. 1 pont
- Ezzel párhuzamosan a magkezdemény burkából létrejön a maghéj, 1 pont
- a magkezdeményből a mag, 1 pont
- a magházból / termőből pedig termés lesz. 1 pont
2023 október
IX.A. Fehérjefunkciók az emberben – esszé 10 pont
Mutassa be a fehérjemolekulák különféle funkcióit az emberi szervezetben, a következő fehérjemolekulák példáján:
1. aktin és miozin;
2. amiláz;
3. keratin;
4. fibrinogén;
5. vazopresszin/antidiuretikus hormon.
Minden egyes fehérje esetében térjen ki arra, hogy mely szerv(ek)ben termelődik a szervezetben, és mi a funkciója!
IX.B. A zárvatermők ivaros szaporodása – esszé 10 pont
Esszéjében ismertesse a megtermékenyítésre kész embriózsák felépítését és a kettős megtermékenyítés folyamatát az alábbi szempontok alapján!
1. Mely sejtek találhatók a megtermékenyítésre váró embriózsákban (nevük, számuk, ploidiatípusuk- azaz melyek a haploid, diploid és triploid sejtek)?
2. Hogyan játszódik le a kettős megtermékenyítés (a lépések sorrendje, eredménye, milyen ploidiatípusú sejte(ke)t eredményeznek)?
Javítási-értékelési útmutató
Fehérjefunkciók az emberben 10 pont
1. aktin és miozin:
- Az izomszövetekben található filamentumok, 1 pont
- az izomösszehúzódást teszik lehetővé. 1 pont
2.amiláz:
- Termelik a nyálmirigyek és a hasnyálmirigy, 1 pont
- keményítőt / poliszacharidokat / oligoszacharidokat emészt a tápcsatornában. 1 pont
3. keratin:
- A bőrben, 1 pont
- mechanikai / kiszáradás elleni védelem. 1 pont
4. fibrinogén:
- A máj termeli, 1 pont
- a véralvadás fehérjéje / fibrinné alakul, majd hálót képez. 1 pont
5. vazopresszin/antidiuretikus hormon:
- A hipotalamusz termeli, 1 pont
- a víz visszaszívását serkenti a vesében. 1 pont
A zárvatermők ivaros szaporodása – esszé 10 pont
1.
- három ellenlábas sejt, 1 pont
- két segítő (kísérő) sejt 1 pont
- egy petesejt 1 pont
- ezek haploidok 1 pont
- egy központi sejt 1 pont
- ez diploid / két haploid sejt összeolvadásából jött létre 1 pont
2.
- vegetatív sejt – pollentömlőt hajt 1 pont
- generatív sejt (számtartó osztódás) – két hímivarsejt alakul ki 1 pont
- egyik hímivarsejt összeolvad a petesejttel: diploid zigóta 1 pont
- másik hímivarsejt összeolvad a központi sejttel: triploid sejt 1 pont
2023 május
IX.A. A kalcium szerepe az ember szervezetében – esszé (10 pont)
Az esszében térjen ki az alábbiakra!
1. A vázizmok működésében nélkülözhetetlenek a Ca2+-ionok. Írja le a harántcsíkolt izmok működésének folyamatát az alábbi szempontok figyelembevételével!
A Ca2+-ionok kötődése, ennek hatása a folyamatban résztvevő fehérjékre, az energiafelhasználás módja, a sötét és világos sávok szélességének változása és ennek magyarázata. (4 pont)
2. A Ca2+-ionok nélkülözhetetlenek a véralvadás folyamatában is. Írja le azokat a protrombinból kiinduló eseményeket, amelyek fibrinháló kialakulásához vezetnek! (2 pont)
3. A kalciumnak a csontok felépítésében is fontos szerepe van. A csontszövet mely alkotórészében, mely sók formájában (egy példa elég) fordulnak elő a Ca2+-ionok, s milyen tulajdonságát adják a csontoknak? (3 pont)
4. Nevezze meg azt a vitamint, amely elősegíti a Ca2+-ionok felszívódását! (1 pont)
IX.B. Megvilágítás és virágzás – esszé (10 pont)
A mikulásvirág virágoztatása sok esetben nehézségbe ütközik hazánkban, mivel virágzását befolyásolja a napi megvilágítás hossza. Mutassa be, hogyan befolyásolja a nappalhosszúság a növények virágzását. Esszéjében az alábbi szempontokra térjen ki!
1. A rövid- és a hosszúnappalos növény fogalma, származása. Magyarázza a különbséget a származási hely földrajzi jellemzőivel! (5 pont)
2. Hazánkban melyik évszakban jellemző a rövid- és a hosszúnappalos fajok virágzása? (2 pont)
3. Melyik kategóriába tartozik a mikulásvirág? Többnyire miért marad el az ablakpárkányon nevelt, szobai világításnak is kitett példányok virágzása? Javasoljon egy lehetséges módszert, amivel el lehet érni, hogy a mikulásvirág nálunk is virágozzon! (3 pont)
Javítási-értékelési útmutató
A kalcium szerepe az ember szervezetében – esszé 10 pont
1.
- a Ca2+-ionok hatására a miozin az aktinszálhoz (troponinhoz) kötődik 1 pont
- az aktinszálak egymás felé / a miozinfonalak közé csúsznak 1 pont
- ennek következtében a világos és sötét sávok (egymáshoz viszonyított) aránya változik 1 pont
- a folyamat ATP-igényes / a kalcium-ionok visszajutása is energiaigényes 1 pont
2.
- protrombinból trombin lesz 1 pont
- a trombin katalizálja azt a folyamatot, amelynek során az oldott állapotú fibrinogénből oldhatatlan fibrinszálak lesznek (majd ezekből alakul ki a fibrinháló) 1 pont
3.
- sejtközötti állományban 1 pont
- Ca-foszfát / karbonát / fluorid / (hidroxiapatit) 1 pont
- szilárdságát / keménységét / színét 1 pont
4.
- D-vitamin 1 pont
Megvilágítás és virágzás – esszé 10 pont
1.
- A rövidnappalos növények kb. azonos hosszúságú nappal és éjszaka / 10-12 óra megvilágítást követő min. 12 órás folyamatos sötétperiódus esetén virágoznak. 1 pont
- A hosszúnappalos növények az éjszaka tartamát meghaladó megvilágítás / 14-16 óra megvilágítás(t követő 8-10 órás sötétperiódus) esetén virágoznak. 1 pont
- A rövidnappalos növények trópusi / szubtrópusi területekről származnak, 1 pont
- a hosszúnappalos növények mérsékelt (vagy hideg) övi területekről származnak. 1 pont
- A különbség oka, hogy a trópusokon nincs jelentős évszaktól függő eltérés a nappalok és éjszakák hossza között, a mérsékelt övben a hosszú / hosszabbodó nappalok / rövidülő éjszakák a nyár közeledtét jelzik. 1 pont
2.
- A rövidnappalos növények hazánkban nyár végén / ősszel / koratavasszal virágoznak, 1 pont
- a hosszúnappalos növények hazánkban tavasszal / nyáron virágoznak. 1 pont
3.
- A mikulásvirág rövidnappalos növény, 1 pont
- virágzásának beindítását a mesterséges fény is gátolja. 1 pont
- Virágzásához hosszabb időre / legalább 12-14 órára teljes sötétségbe kell tenni a növényt. 1 pont
Más hasonló, a tartós sötétségre utaló válasz is elfogadható.
2022 október
IX.A. Szembetegségek – esszé 10 pont
Írjon rövid esszét a látást veszélyeztető felsorolt megbetegedésekről! Fogalmazásában térjen ki az alábbi szempontokra:
1. A rövid- (közel)látás fizikai oka, következménye, a nem műtéti kezelési lehetősége. (3 pont)
2. A vörös-zöld színtévesztés öröklődésének jellemzői, megoszlása a nemek között. (3 pont)
3. A zöldhályog és a szürkehályog oka, a látásra gyakorolt hatásuk. (4 pont)
IX.B. Természetvédelem – esszé 10 pont
1. Értelmezze a kihalási küszöb fogalmát, kapcsolatát a genetikai sodródással és a beltenyészet következtében föllépő leromlással! Magyarázza el, hogy mely természetvédelmi intézkedésekkel csökkenthető ennek veszélye! (4 pont)
2. Hozzon két példát a természetvédelem jogi lehetőségeire a fajok védelme érdekében, valamint két példát az aktív természetvédelem fajmegőrző szerepére! (4 pont)
3. Mutassa be egy példán keresztül a tájidegen fajok behurcolásának, bejutásának veszélyeit! (2 pont)
Javítási-értékelési útmutató
Szembetegségek – esszé 10 pont
1.
- A rövid- / közellátó szemben az éles kép a retina előtt keletkezik / a szemtengely hosszabb, mint az éles képalkotáshoz szükséges / a szem fénytörő képessége a szükségesnél nagyobb 1 pont
- ezért a távoli tárgy képe homályos lesz. 1 pont
- Szóró / homorú / konkáv lencsével korrigálható. 1 pont
2.
- A vörös-zöld színtévesztés recesszíven, 1 pont
- X-kromoszómához kötötten öröklődik 1 pont
- ezért / (a hemizigóta öröklés miatt) a férfiakban gyakoribb. 1 pont
3.
- A zöldhályog esetében csarnokvíz felszaporodik / a magas nyomás miatt változik a szaruhártya fénytörése, 1 pont
- (a látóideg pusztulása miatt) látásromlás / látótérkiesés / vakság alakulhat ki. 1 pont
- A szürkehályog a szemlencse fényáteresztő-képességének csökkenése, 1 pont
- emiatt elhomályosul a látás, csökken a szem fényérzékenysége. 1 pont
Természetvédelem – esszé 10 pont
1.
- A kihalási küszöb alatti populációlétszám nem teszi lehetővé a faj tartós fennmaradását (az adott területen). 1 pont
- Oka: a véletlen változások, így a genetikai sodródás változatosságot csökkentő hatása, 1 pont
- a kis populációkban föllépő beltenyészet miatt nő a recesszíven öröklődő betegségek / rendellenességek megjelenésének kockázata. 1 pont
- Természetvédelmi vonatkozások: megfelelő nagyságú terület / megfelelő élőhelyi változatosság stb. fenntartása ahhoz, hogy a védendő faj létszáma ne essen a kihalási küszöb alá. Más konkrét példa, pl. ökológiai folyosók fenntartása is jó válasz. 1 pont
2.
- A védett fajok gyűjtése / vadászata tilos, 1 pont
- a belőle készült termékek kereskedelme is tiltott: 1 pont
- valamely példa említése, pl: szőrmék, elefántcsont, kaktuszgyűjtés stb. 1 pont
- Aktív védelem: pl. sérült példányok gyógyítása / tenyésztés-szabadon engedés / magbankok / invazív, behurcolt fajok visszaszorítása. 1 pont
Más faj és a védelem másféle módjának bemutatása is elfogadható.
3.
- A behurcolt fajok kiszoríthatják az őshonosakat / betegségeket terjeszthetnek / mezőgazdasági károkat okozhatnak 1 pont
- Valamely példa említése, pl: parlagfű – allergia / tigrisszúnyog – vírusos betegségek. Más veszélyforrás és példafaj említése is megfelelő. 1 pont
2022 május
X.A. A vörösvértestek – esszé 10 pont
Mutassa be a vörösvértestek felépítését és szerepét! Fogalmazásában térjen ki az alábbi vázlatpontok mindegyikére!
1. A vörösvértestek felépítésének jellegzetességei (színének magyarázata, alakja). (2 pont)
2. A membránjukon található jellegzetes antigének megnevezése, jelentőségük a vérátömlesztésben. (2 pont)
3. A vörösvértestek feladata, magashegyi edzőtáborok hatása a vörösvértestek számára, hatása a sportteljesítményre, indoklással. (3 pont)
4. A vérszegénység három, elvileg különböző, lehetséges oka. (3 pont)
X.B. Erdők – esszé 10 pont
Leírásában hasonlítsa össze a cseres-tölgyes és a bükkös életközösségeket az alábbi szempontok alapján:
1. Zonális elhelyezkedésük, a különbség oka. A bükkösök visszaszorulásának éghajlati magyarázata. (4 pont)
2. Szerkezetük: gyepszint, különbségek a tavaszi és a nyári aszpektusokban, a változás oka; a cserjeszint jellemzői, magyarázata, lombkoronaszint. (6 pont)
Javítási-értékelési útmutató
A vörösvértestek – esszé 10 pont
1.
- Vörös színüket a sejtplazmában elhelyezkedő hemoglobin adja, 1 pont
- alakjuk fánk/ korong / súlyzó. 1 pont
2.
- Membránjukon vércsoportantigének / A, B, D, 0 antigének helyezkedhetnek el. 1 pont
- Vérátömlesztésnél/transzfúziónál ügyelni kell a csoportazonos vér adására. 1 pont
3.
- A vörösvértestek feladata a légzési gázok/ oxigén és szén-dioxid szállítása. 1 pont
- A magas hegységekben (az oxigén alacsony parciális nyomására) a szervezet fokozott mennyiségű vörösvértest-előállítással válaszol, 1 pont
- az alacsonyabb szintre visszaérkezve a megemelkedett vörösvértest-szám nagyobb teljesítményt tesz lehetővé/ hatékonyabb oxigénkötést biztosít, ezért tartanak edzőtáborokat a hegységekben. 1 pont
4.
- A vérszegénység oka lehet vashiány - a vas elégtelen bevitele vagy felszívódása/ vitaminhiány / B12 vitamin/folsav hiánya, / leukémia következménye nagyfokú vérveszteség / krónikus betegség / a hemoglobin örökletes betegségei / vérképzés zavara. / Rézanyagcsere zavara
1-1 pont, 3 különböző ok említése összesen: 3 pont
Erdők – esszé 10 pont
1. Zonális elhelyezkedés:
- bükkös: 600 m felett 1 pont
- cseres-tölgyes: 400m alatt 1 pont
- A különbség oka: a bükk alacsonyabb átlaghőmérsékletet / több csapadékot igényel, mint a csertölgy. 1 pont
- Visszaszorulásuk magyarázata: csökkenő csapadékátlag / növekvő átlaghőmérséklet 1 pont
2. Az erdő szerkezete
- bükkös gyepszint: kora tavasszal nyíló hagymások, gumósok/ koratavaszi geofita aszpektus 1 pont
- Nyáron szegényes 1 pont
- a lombkoronaszint tavasz végi záródása miatt. 1 pont
- cserjeszint, bükkös: alig/csak a fák magoncai/ fiatal fák 1 pont
- cseres-tölgyes: egész évben gazdag gyep- és cserjeszint 1 pont
- a laza lombkorona / jó megvilágítás miatt. 1 pont
2021 október
IX.A. A női nemi ciklus második szakasza – esszé 10 pont
Mutassa be a női nemi ciklus második szakaszának eseményeit! Esszéjében az alábbi pontokra térjen ki:
1. Melyik hipofízis (agyalapi mirigy) hormon melyik női nemi hormon termelését kezdi jelentősen fokozni ebben a szakaszban? Mi termeli ezt a hormont a petefészekben és mi a fő hatása a méh állapotára? (4 pont)
2. Írja le a negatív visszacsatolási folyamatot és az általa beindított eseménysort, amely a ciklus befejezéséhez vezet, ha a petesejt nem termékenyül meg! (4 pont)
3. Mely hormonok teszik lehetővé, illetve jelzik a várandós állapotot az embrió / csíra beágyazódása esetén? (2 pont)
IX.B. Életmódváltás – esszé 10 pont
Írjon esszét a klímaváltozás okairól az alábbi szempontok alapján:
1. Mit értünk fosszilis és megújuló energiahordozó alatt? Írjon példákat is! (3 pont)
2. Az amazóniai erdőégetés miért és hogyan befolyásolja az üvegházhatást? (2 pont)
3. Írjon két példát a biomassza energetikai felhasználására, illetve egy-egy érvet használata ellen és mellett! (3 pont)
4. Adjon egy-egy javaslatot a városi, illetve a távolsági közlekedés káros hatásának csökkentésére! (2 pont)
Javítási-értékelési útmutató
A női nemi ciklus második szakasza – esszé 10 pont
1.
- A hipofízisben termelődő sárgatestserkentő (LH) hormon 1 pont
- hatására a sárgatestben termelt 1 pont
- progeszteron 1 pont
- fenntartja a méh nyálkahártyájának megvastagodott állapotát. 1 pont
2.
- A magas progeszteronszint gátló / negatív visszacsatoló hatása gátolja az LH termelését, 1 pont
- ami a sárgatest elsorvadásához vezet, melynek hatására 1 pont
- a progeszteronszint lecsökken, 1 pont
- ami kiváltja a menstruációt. 1 pont
3.
- Beágyazódás esetén a progeszteronszint magas marad, 1 pont
- (a vérben / vizeletben az embrió, később a méhlepény által termelt) terhességi hormon / hCG jelenik meg. 1 pont
Életmódváltás – esszé 10 pont
1.
- A fosszilis energiahordozó hosszú idő alatt kialakult biológiai eredetű, széntartalmú energiahordozó 1 pont
- Megújuló: fogyásának és keletkezésének sebessége kb. azonos 1 pont
- Fosszilis: kőszén / kőolaj / földgáz, megújuló: víz / nap / szél. Bármely két jó példa 1 pont
2.
- Az égés során nő a levegő CO2-tartalma 1 pont
- A termőterület csökkenése / erózió miatti CO2-szint növekedés. / csökken a CO2 megkötése. 1 pont
3.
- Biodízel / bioetanol / fatüzelés – 2 példa 1 pont
- Ellene: az égési folyamatok során ugyanannyi káros anyag keletkezik, mint más égési folyamatoknál / termőterületet vesz el az előállítása / csökkentheti a biodiverzitást / előállítása is energiaigényes/ Más jó érv is elfogadható. 1 pont
- Mellette: Megújuló energiaforrás. / Nem növeli érdemben a fosszilis eredetű CO2 mennyiségét. 1 pont
4.
- Városi közlekedés: közösségi közlekedés / kerékpár / gyalogos közlekedés / forgalomszervezési megoldások / Más jó megoldás is elfogadható. 1 pont
- Távolsági közlekedés: Vasút használata közút helyett / kevesebb utazás / Más jó megoldás is elfogadható. 1 pont
2021 május
IX.A. A víz távozása a szervezetből – esszé 10 pont
Írjon fogalmazást arról, milyen folyamatok eredményeképpen távozik víz a testünkből!
Esszéjében az alábbiakra térjen ki!
1. A szűrlet fogalma, keletkezésének magyarázata (a nyomáskülönbség oka) és helye a nefronban. (3 pont)
2. A vízvisszaszívást biztosító fizikai-kémiai transzportmechanizmus. Mely anyagok koncentrációkülönbsége biztosítja az ehhez szükséges hajtóerőt? (3 pont)
3. A vizelet útja a veséktől a külvilágig. (1 pont)
4. A vizelet napi térfogatának növekedését, illetve csökkenését okozó élettani helyzetek egészséges emberben. (1-1 példa) (2 pont)
5. Két példa olyan egészséges folyamatra, amely a vizeletürítésen kívül jelentősebb mennyiségű vizet távolít el a szervezetünkből. (1 pont)
IX.B. A biodiverzitás szintjei - esszé 10 pont
Mutassa be, milyen szinteken értelmezhető a biodiverzitás és mi a megőrzésének jelentősége!
Esszéjében az alábbi szempontokat érintse:
1. Magyarázza meg, hogy a populációk szintjén értelmezett (faj-egyed) biodiverzitás értékének megadásához mit kell figyelembe venni! Nevezze meg az életközösség egy olyan tulajdonságát, mely összefügghet a biodiverzitás értékével, és magyarázza a kapcsolatot! (3 pont)
2. Magyarázza meg, hogy a genetikai diverzitás értelmezéséhez mit kell figyelembe venni!
Értelmezze, hogy ennek az alkalmazkodás folyamatában mi a jelentősége! Értelmezze a sodródás fogalmát és következményét ebből a szempontból! (5 pont)
3. Magyarázza meg, hogy a biodiverzitás védelme mi okból jelent mást, többet, mint az egyes fajok védelme! (2 pont)
Javítási-értékelési útmutató
A víz távozása a szervezetből – esszé 10 pont
1.
- A szűrlet fehérjementes vérplazma 1 pont
- a vesetestecskében / Malpighi-testecskében keletkezik, 1 pont
- a vérnyomáskülönbség hatására: utalás a bemenő és kilépő artériák átmérőjének különbségére. 1 pont
2.
- a víz diffúzióval / ozmózissal / passzív transzporttal szívódik vissza (a közeli elsődleges kanyarulatos csatornát kísérő hajszálerekben) a szűrletképzést követően 1 pont
- (relatíve) kevés víz, de (csaknem) változatlan anyagmennyiségű fehérje marad, emiatt magas a vér (kolloid) ozmózisnyomása, 1 pont
- (a Henle-kacs területén) aktív transzport / pumpamechanizmusok / ionpumpák növelik meg a vér Na+-ion koncentrációját / ozmózisnyomását 1 pont
3.
- (vesemedence/vesemedencék) – húgyvezeték(ek) – húgyhólyag – húgycső (együtt) 1pont
4.
- növeli: a szükségesnél nagyobb mennyiségű folyadék fogyasztása/vízhajtó hatású italok (kávé, tea, alkohol stb.) 1 pont
- csökkenti: a szükségesnél kisebb mennyiségű folyadék fogyasztása / fokozott verejtékezés / menstruáció előtti vízvisszatartás / erősen sós táplálék fogyasztása 1 pont
5.
- verejtékezés / levegő kilégzése / székletürítés (bármely kettő) 1 pont
A biodiverzitás szintjei – esszé 10 pont
1.
- A faj / faj-egyed diverzitást az egy területen élő fajok száma 1 pont
- és azok egyedszám-aránya / eloszlása szabja meg. 1 pont
- A faj / (egyed) diverzitás összefügg a társulás stabilitásával / biológiai produkciójával / a populációs kölcsönhatások sokféleségével / táplálkozási hálózatok tagjainak számával, mert fajgazdagabb / egyenletesebb eloszlású életközösségben többféle kapcsolat valósulhat meg. Más tulajdonság is elfogadható, ésszerű indoklással. 1 pont
2.
- A genetikai diverzitás a populációban előforduló allélok számától 1 pont
- és gyakoriságuk arányától / eloszlásuktól függ. 1 pont
- Magas genetikai diverzitás esetén a populáció alkalmazkodóképesebb, mert lehetőséget ad a szelekcióra / a populáció tagjainak együttműködésére. 1 pont
- A sodródás allélok véletlenszerű kiesését okozhatja, 1 pont
- ezért a genetikai diverzitást csökkenti. (palacknyak- / üvegnyak-hatás) 1 pont
3.
- A diverzitást a fajok kölcsönhatása tartja fenn. 1 pont
- ezért megőrzése az egyes fajok fennmaradást biztosítja (pl. táplálkozási hálózatok / szimbiózisok). 1pont
Másként is megfogalmazható. A megfogalmazás utaljon a társulás fajainak kölcsönös függésére.
2020 október
IX.A. A menstruációs ciklus – esszé 10 pont
Ismertesse a menstruációs ciklus történéseit! Esszéjében az alábbi szempontokat kövesse:
1. Az agyalapi mirigy hormonjai, amelyek a menstruációs ciklust befolyásolják – fő hatásukkal. (5 pont)
2. A tüszőérés és a sárgatest kialakulása, szerepe a ciklus során. (2 pont)
3. A méhnyálkahártya változásai a női nemi ciklus során. a menstruáció hormonális oka. A petefészek hormonjainak hatása a méhnyálkahártya állapotára. (3 pont)
IX.B. Ökológiai kölcsönhatások – esszé 10 pont
Mutassa be az élővilág nagy országainak néhány képviselője közt fellépő populációs kölcsönhatásokat! A következőkre térjen ki:
1. Az antibiózis és az antibiotikumok fogalma és hasznosítása. (3 pont)
2. Az ember és a szervezetében élő baktériumok közötti szimbiózis, asztalközösség (kommenzalizmus) és parazitizmus (a baktériumcsoportokat anyagcseréjük, illetve a szervezetben betöltött szerepük vagy hatásuk alapján nevezze meg). (3 pont)
3. A szintezettség és az aszpektusok soksejtű növények természetes életközösségében (egy jellemző életközösség megnevezésével, a jelenség meghatározása és bemutatása példákon keresztül) (4 pont)
Javítási-értékelési útmutató
A menstruációs ciklus 10 pont
1.
- Az agyalapi mirigy tüszőserkentő hormonja megindítja a tüszőérést. 1 pont
- A sárgatestserkentő hormon(nal együtt) kiváltja a tüszőrepedést. 1 pont
- A ciklus közepén bekövetkezik az ovuláció / tüszőrepedés, ekkor 1 pont
- a petesejt kiszabadul a petefészekből és elindul a petevezetőben. 1 pont
2.
- A visszamaradó tüsző sárgatestté alakul, 1 pont
- amely progeszteron hormont / sárgatest-hormont termel a ciklus második felében. 1 pont
3.
- A progeszteron szint csökkenése váltja ki a menstruációt. 1 pont
- A méh nyálkahártyája a menstruációkor vérzés kíséretében leválik. 1 pont
- Ösztrogén hatására a ciklus első felében újraképződik. 1 pont
- A ciklus második felében progeszteron hatására tovább vastagodik, mirigyekben gazdagodik. 1 pont
Ökológiai kölcsönhatások 10 pont
1.
- Az egyik fajnak közömbös, a másiknak előnytelen kölcsönhatás 1 pont
- Az antibiotikumok (többnyire) gombák által termelt, baktériumok szaporodását gátló anyagok 1 pont
- Felhasználásuk: baktériumos eredetű betegségek kezelésére. 1 pont
2.
- A vastagbélben élő baktériumok egy része szimbionta, mert vitaminokat / átvivőanyagokat termel / mert gátolja más baktériumok vagy gombák elszaporodását, 1 pont
- más részük kommenzalista, mert a vastagbélbe került emésztetlen szerves anyagok lebontását végzi, 1 pont
- bekerülhetnek szervezetünkbe élősködő / betegségokozó fajok is (bármely baktériumos eredetű megbetegedés megnevezése) 1 pont
Más helyszín és kölcsönhatás is leírható, pl. a bőrön vagy a hüvelyben élő baktériumok szerepe.
3.
- Szintezettség: a társulások vertikális szerkezete. Például a gyertyános-tölgyesek lombkorona- és mohaszintjének elkülönülése, 1 pont
- az eltérő fényigényű / páraigényű fajok térbeli elkülönülése miatt. 1 pont
- Aszpektusok: az életközösség évszakoktól függő változásai, például a bükkösök tavaszi / nyári / téli aszpektusa, 1 pont
- tavaszi aszpektus: a lombokorona záródása előtt virágzó lágyszárúak tömeges megjelenése a megvilágítás évszakos változásával magyarázható. 1 pont
Más példák és ezek magyarázata is elfogadható.
2020 május
IX.A. A baktériumtranszformáció és a gyógyítás – esszé 10 pont
Esszéjében írja le, hogy Griffith és Avery kísérletében (baktériumtranszformáció) milyen eredményre jutottak, és felfedezésüknek mi a jelentősége a gyógyításban!
Esszéjében az alábbiakra térjen ki:
1. Milyen módszerrel mit igazolt Griffith, és mit bizonyított Avery kutatócsoportja? Mit igazolt a két kutató kísérletsorozata a prokarióták információátadásáról? (4 pont)
2. Mit értünk a génmódosított (GMO) élőlények létrehozásán? Hozzon egy példát a genetikailag módosított élőlények hasznosítására a gyógyászatban! (2 pont)
3. Mutassa be egy bakteriális eredetű, gyulladásos betegség jellemző tüneteit! Induljon ki a gyulladás általános jellemzőiből és alkalmazza ezt a tüdőre vonatkozóan! (2 ok – 2 következmény) (4 pont)
IX.B. Az élő bolygó – esszé 10 pont
Fogalmazza meg James Lovelock Gaia-elméletének lényegét! Esszéjében a következőkre térjen ki!
1. Fogalmazza meg Gaia szabályozottságának lényegét és funkcióját az elmélet alapján! (2 pont)
2. Írjon példákat arra, hogy az élővilág hogyan hat a környezeti tényezőkre az alábbi esetekben: (4 pont)
- a vízgyűjtő területen fekvő erdők hatása a folyók vízjárására (az erdőtlen területekhez képest),
- pillangósvirágú növények hatása a talaj termőképességére,
- felnövekvő erdőrészletek hatása a légkör pára- és oxigéntartalmára.
3. Mutassa be, hogy milyen emberi tevékenység okozhatja a talajvíz nitrátosodását és a csapadék savasodását! Mutassa be ezek hatását biológiai rendszerekre, illetve az ember egészségére! (4 pont)
Javítási-értékelési útmutató
1.
- Módszer: A kutatók tokképző, illetve tok nélküli / (S és R változatú) (tüdőgyulladás) baktériumokkal (Pneumococcusokkal) fertőztek egereket: a tokképző volt a halálos. A hővel elölt tokképző nem okozott betegséget, de a hővel elölt tokképző és a tok nélküli keverékben létrejöttek a tokos változatok (baktériumtranszformáció). 1 pont
- A tokképződésre vonatkozó információ átjuthat egyik sejtről a másikra (Griffith kísérletei) 1 pont
- Módszer: A keverékhez adott fehérje- vagy RNS-bontó enzimek nem gátolták a transzformációt, a DNS-bontó enzimek azonban igen. 1 pont
- Bizonyítás: tehát a tokképzésre vonatkozó információt a DNS vitte át a kísérlet során. (Averyék eredménye) 1 pont
2.
- A genetikai módosítás egy vagy több gén (transzgén) szándékos / célzott bejuttatása egy másik faj egyedeibe / gyakran eukarióta és prokarióta között megy végbe. 1 pont
- Génmódosított baktériummal pl. lehetséges inzulint előállítani, ami lehetővé teszi a cukorbetegség kezelését. 1 pont Más gyógyászati célú példa is elfogadható.
 |
Az élő bolygó – esszé 10 pont
1.
- A Földön zajló (globális) anyagáramlási folyamatok szabályozottságuk miatt / visszacsatolások révén sok anyag koncentrációját határértékek között tartják, 1 pont
- ennek funkciója a földi élet tartós fenntartásának biztosítása. 1 pont
Másképp is megfogalmazható. A bemutatás jelezze a szabályozottság lényegét és funkcióját.
2.
- Az ilyen erdők egyenletesebbé teszik a folyók vízjárását, 1 pont
- A pillangósvirágúak / gyökérgümőiben élő nitrogénfixáló baktériumok segítségével növelik a talaj termőképességét, 1 pont
- a növekvő fák oxigént termelnek, 1 pont
- és párássá teszik a levegőt. 1 pont
3.
- A (túlzott) (mű)trágyázás / csatornázatlanság növeli a talajvíz nitráttartalmát, 1 pont
- ami csecsemőkre súlyos veszélyt jelent / methemoglobinémia / redukálódva rákkeltő nitritekké alakulhat (bármelyik veszélyes hatás említése) 1 pont
- A fosszilis energiahordozók (főként: kőszén) égetése savas esőket okoz / csökkenti a csapadék pH-ját, 1 pont
- ami (közvetve vagy közvetlenül) erdőpusztulást okozhat. 1 pont
Más, savasodással vagy növekvő nitráttartalommal összefüggő hatás (pl. eutrofizáció) és ezek lehetséges biológiai következményeinek megemlítése is elfogadható.
2019 október
IX.A. A retina és a látás – esszé 10 pont
Esszéjében mutassa be az éles látás és a távolságbecslés fizikai-biológiai hátterét!
A következőkre térjen ki!
1. Jellemezze a retinán kialakuló kép fizikai tulajdonságait (nagyság, helyzet)! (2 pont)
2. Magyarázza, hogy a retina melyik területén a legélesebb a látás! (2 pont)
3. Mutassa be, hogy mely izom segítségével hogyan alkalmazkodik a szem, amikor távoli tárgyról közelire pillantunk! (3 pont)
4. Mutasson be három észlelési módot, ami a látás során lehetőséget ad a megfigyelt tárgyak távolságának becslésére, érzékelésére! (3 pont)
IX.B. Gazdálkodás a nitrogénnel - esszé 10 pont
Esszéjében foglalja össze a nitrogén körforgásával kapcsolatos legfontosabb környezeti és gazdálkodási kérdéseket! Az alábbiakra térjen ki!
1. Mely ionok formájában veszi fel a legtöbb növény a nitrogént? (1 pont)
2. Milyen természetes és mesterséges folyamatok pótolhatják a mezőgazdasági talaj elvesztett nitrogéntartalmát? Mit jelent a minimumelv (Liebig-elv), és milyen gazdálkodási szabály fogalmazható meg ennek figyelembevételével? (4 pont)
3. Milyen következményekkel jár, ha az ivóvízbe sok nitrogéntartalmú ion kerül? (3 pont)
4. Milyen kémiai-biológiai problémákat okoz, ha a levegőbe sok nitrogén-oxid kerül? (2 pont)
Javítási-értékelési útmutató
A retina és a látás – esszé 10 pont
1.
- A retinára kicsinyített, 1 pont
- fordított állású kép vetül. 1 pont
2.
- A sárgafoltban a legélesebb a látás, 1 pont
- mert a csapok itt állnak a legsűrűbben / itt csak csapok vannak. 1 pont
3.
Közelre nézéskor
- a sugártest izmai megfeszülnek, a sugártest bedomborodik, 1 pont
- a lencsefüggesztő rostok ellazulnak, 1 pont
- a szemlencse domborúbbá válik / fókusztávolsága csökken/fénytörése nő. 1 pont
4.
A távolságról információt nyújt
- a szemtengelyek állása / a külső szemmozgató izmok / a tárgyak takarása (a közelebbi takarja a távolabbit), / az ismert nagyságú tárgyak retinára vetülő képének nagysága / perspektivikus rövidülés / mozgás során az elmozdulás látható gyorsasága /a távolabbi tárgyak színének módosulása.
Bármelyik három leírása: 1+1+1 = 3 pont
Gazdálkodás a nitrogénnel - esszé 10 pont
1.
- Nitrát /NO3 – ionok formájában. (Elfogadható az ammónium-ion is.) 1 pont
2.
- Természetes úton a nitrogéngyűjtő baktériumok / a villámlás során keletkező nitrogénvegyületek. 1 pont
- Mesterségesen: trágyázás / műtrágyázás útján. 1 pont
- A minimum-elv szerint csak a legkisebb koncentrációban hozzáférhető ion arányában képes hasznosítani a növény a többit, 1 pont
- ezért csak annyi nitrogén(mű)trágyát szabad használni, amennyinek a hasznosítását a többi felvehető ion lehetővé teszi. 1 pont
3.
- Az ammónium-ion mérgező / undort keltő, / a nitrit-ion rákkeltő/ egészségkárosító. / A nitrát-ion nitrit-ionná redukálódhat. Bármelyik. 1 pont
- A nitrát-ionok csecsemőkben fulladást okoznak / methemoglobinémia, mert a (csecsemőkori) hemoglobinhoz kapcsolódva gátolják annak oxigénszállítását. 1+1 = 2 pont
4.
- A nitrogén-oxidok savas esőt okoznak, / mely növényzetpusztulást / korróziót okoz.
- A (fotokémiai) szmog / füstköd alkotóiként egészségkárosítók. 1+1 = 2 pont
2019 május
IX.A. Az agyalapi mirigy – esszé 10 pont
Ismertesse az agyalapi mirigy működésének megváltozását egy benne képződő hormontermelő daganat következtében! Az alábbiakra térjen ki:
1. Hogyan befolyásolja a daganat a pajzsmirigy működését? Milyen hatása van a beteg anyagcseréjére, testhőmérsékletére, testsúlyára és idegrendszeri működésére? (5 pont)
2. Milyen hatása van az elváltozásnak a mellékvese működésére? (3 pont)
3. Milyen következményekkel járhat együtt a növekedési hormon túltermelése fiatal- és felnőtt korban? (2 pont)
IX.B. Folyadékáramlás az erdőkben – esszé 10 pont
Mutassa be a folyadékáramlás okát és következményeit a fákban, illetve hazai erdeinkben!
Az alábbiakra térjen ki:
1. Mely tényezők biztosítják a víz áramlását a gyökértől a levelek felé? Mely szövetelemekben zajlik ez a folyamat? (4 pont)
2. Milyen következményekkel járhat a (már megindult anyagcseréjű) fatörzsben a hirtelen tavaszi fagy? (1 pont)
3. Hogyan befolyásolja az erdei életközösség életműködéseivel a folyók vízjárását, a talaj és a légkör összetételét? (5 pont)
Javítási-értékelési útmutató
Az agyalapi mirigy – esszé 10 pont
1.
- A daganat hatására fokozódik a pajzsmirigyben a hormontermelés / a tiroxin elválasztása / leadása, 1 pont
- aminek hatására az anyagcsere felgyorsul / a lebontó anyagcsere aránya nő, 1 pont
- a (normál) testhőmérséklet megemelkedik, 1 pont
- az idegrendszer aktivitása fokozódik / a betegre idegesség / alvászavar jellemző / ingerküszöbe csökken 1 pont
- (éhségérzet / sok táplálék ellenére) testsúlya csökken. 1 pont
2.
- A mellékvesekéreg-serkentő hormon termelésének fokozódása miatt
- a mellékvese kéregállományának működése is fokozódik,
- elsődlegesen a szénhidrátanyagcserére ható hormonok / glükokortikoidok elválasztásának fokozása
- a szénhidrátanyagcsere zavarát / a glikogénraktárak kiürülését okozza / megemelkedő vércukorszintet okoz / befolyásolja (gátolja) az immunrendszert.
- Részben a mineralokortikoidok / szexuálszteroidok elválasztása is fokozódik.
Bármely három következmény megfogalmazása 1+1+1 = 3 pont
Ha a vizsgázó hibás előfeltevésből (a gátlásból) indult ki, de a továbbiakban az oksági összefüggéseket helyesen elemezte, a gondolatmenet első 1-1 pontját leszámítva értékelhető a válasza, ami maximálisan 4+2 = 6 pont.
3.
A növekedési hormon túltermelése
- fiatal korban óriásnövést 1 pont
- felnőttkorban a csúcsi szervek / fül, orr nyelv megnagyobbodását / akromegáliát okoz. 1 pont
Folyadékáramlás az erdőkben – esszé 10 pont
1.
- A víz a gyökérnyomás, 1 pont
- a kapillaritás / kohézió 1 pont
- és a levelek párologtatása (következtében fellépő szívóerő/ ozmózisnyomás) miatt áramlik 1 pont
- a farész vízszállító csövein / (részben vízszállító sejtjein) át. 1 pont
2.
- A hirtelen fagy a működő, vízzel telt szállítószövetet szétrepesztheti (fagykár).1 pont
3.
- A fák (párologtatásuk miatt) párássá, 1 pont
- fotoszintézisük miatt oxigéngazdaggá teszik a légkört, 1 pont
- csökkentik a szén-dioxid szintet / szennyező anyagot kötnek meg. 1 pont
- Az erdei életközösség a talajt tápanyagban / humuszban gazdaggá teszi / az eróziót gátolja 1 pont
- Emiatt a folyók vízjárását egyenletesebbé teszik / az árvízveszélyt csökkentik. 1 pont
2018 október
IX.A. Az agyalapi mirigy hormontermelése – esszé 10 pont
Írjon rövid esszét az agyalapi mirigy hormontermeléséről! Fogalmazásában térjen ki az alábbi szempontokra:
1. Az agynak pontosan melyik részével áll közvetlen működési kapcsolatban az agyalapi mirigy? Röviden írja le működési kapcsolataik lényegét! (2 pont)
2. A hátsó lebenyhez köthető hormonok neve, hatásai. (4 pont)
3. Melyek a növekedési hormon hiányának és túltermelődésének következményei? (2 pont)
4. A pajzsmirigy hormontermelését közvetlenül szabályozó hormon működésében mely szabályozási mechanizmus hogyan valósul meg? (2 pont)
IX.B. A növények párologtatása – esszé 10 pont
Írjon esszét a növények párologtatásáról! Térjen ki a következőkre:
1. Nevezze meg a növényi párologtatás helyszínét (szerv és szövettani részlet) valamint mechanizmusát! (3 pont)
2. Írja le, hogyan segíti a párologtatás a vízáramlást a növényben! (3 pont)
3. Mely környezeti tényezők befolyásolják a párologtatás mértékét! Említsen kettőt! (2 pont)
4. Száraz élőhelyeken honos növények milyen módon csökkentik párologtatásukat? Írjon két példát a testszerveződésben megnyilvánuló alkalmazkodásra! (2 pont)
Javítási-értékelési útmutató
Az agyalapi mirigy hormontermelése – esszé 10 pont
1.
- Az agyalapi mirigy a hipotalamuszhoz kapcsolódik. 1 pont
- A hipotalamusz termeli a hátsó lebeny hormonjait és szabályozza az elülső lebeny hormontermelését. 1 pont
2.
- A hátsó lebenyből ürül az oxitocin, 1 pont
- ami simaizom-összehúzódást okoz a méhfalban / emlőmirigyekben, 1 pont
- illetve a vazopresszin / ADH, 1 pont
- ami fokozza a víz visszaszívását a szűrletből. 1 pont
(A mechanizmusra vonatkozó megfogalmazás is helyes: víz számára átjárhatóvá teszi a nefron / elvezető csatorna / gyűjtőcsatorna falát.)
3.
- Hiányában arányos törpenövés, 1 pont
- magas szintje esetén óriásnövés /gigantizmus / akromegália alakul ki. 1 pont
4.
- A (pajzsmirigyserkentő) hormon termelését a tiroxin gátolja, ami a tiroxinszintjének beállítását eredményezi. (Másként is megfogalmazható.) 1 pont
- Ez negatív visszacsatolás 1 pont
A növények párologtatása – esszé 10 pont
1.
- A párologtatás szerve a levél.
- A gázcserenyílásokon keresztül
- diffúzióval a levegőbe kerül a vízgőz. 3 pont
2.
- A párologtatás hozza létre a levelek szívóhatását
- a vízgőz távozása miatt a levél sejtjeiben nő a koncentráció / ozmózisnyomás
- így a párologtatás a folyadék áramlását biztosítja a nagyobb ozmózisnyomású / koncentrációjú hely felé 3 pont
3.
- Pl. hőmérséklet / szél / levegő páratartalma /a vízutánpótlás / talaj víztartalma (Bármilyen 2 jó megoldás esetén.) 2 pont
4.
- Pl. kis felületű levelek / besüllyedt gázcserenyílások / szőrözött levél / viasz (Bármilyen 2 jó megoldás esetén.) 2 pont
2018 május
IX.A. A szőlőcukor útja a szervezetben – esszé 10 pont
Egy gyermek kiflit majszol. A kifli keményítőtartalma glükózzá emésztődik a tápcsatornájában.
Ismertesse a glükóz útját a gyermek szervezetében az alábbiak szerint:
1. A tápcsatorna melyik szakaszában, hová szívódik fel a szőlőcukor? (2 pont)
2. Mely hormon segíti raktározását poliszacharid formában? (1 pont)
3. Melyik biokémiai folyamatban bontják le a glükózt energianyerés céljából a sejtek oxigén jelenlétében és oxigén hiányában? (A folyamatok megnevezése, sejten belüli helyszíne.) Nevezze meg e lebontó folyamatok széntartalmú végtermékeit, és ismertesse azok további útját a szervezetben! (7 pont)
IX.B. A szén-dioxid útja a növényekben – esszé 10 pont
A gulya szerves anyagai eredendően a puszta növényeinek szerves anyagaiból jönnek létre. Az is igaz, hogy a szarvasmarhák által leadott szén is visszakerülhet a körforgásba. Mutassa be, hogy milyen utat járnak be a gulya által kilélegzett szén-dioxid szénatomjai a növényi szervezetben! Esszéjében az alábbi gondolatokra térjen ki!
1. A biológia oxidáció mely folyamatai során keletkezik a szarvasmarhák és a növények sejtjeiben CO2? (2 pont)
2. Kövesse végig a CO2 szénatomjának útját a növényi szerves anyaggá alakulásig! Nevezze meg a felvétel mechanizmusát és helyét (szövettípus, és azon belül a pontos helyszín)! Írja le, mely biokémiai folyamat során hasznosul a szén-dioxid és mely anyagok biztosítják ehhez az energiát, illetve a folyamathoz szükséges hidrogént! (6 pont)
3. Írja le, hogy mely két poliszacharid keletkezik a növényekben a fotoszintézis végtermékéből! (2 pont)
Javítási-értékelési útmutató
A szőlőcukor útja a szervezetben – esszé 10 pont
1.
- A glükóz a vékonybélből 1 pont
- a véráramba / kapillárisokba szívódik fel. 1 pont
2.
- Az inzulin serkenti a raktározást. / Elfogadható a glükokortikoidok hatásának leírása is. 1 pont
3.
- Oxigén jelenlétében a biológiai / terminális oxidációban 1 pont
- a mitokondriumokban, 1 pont
- oxigén hiányában tejsavas erjedésben. 1 pont
- a sejtplazmában bomlik. 1 pont
- Az oxidációban a glükóz széntartalma szén-dioxiddá alakul, 1 pont
- amely a keringés útján a tüdő léghólyagocskáiba / a kilégzés folyamatában a külvilágba kerül. 1 pont
- A tejsav a vérárammal a májba jut. 1 pont
A szén-dioxid útja a növényekben – esszé 10 pont
1.
- A szarvasmarhák / és a növények szervezetében / a piroszőlősav átalakulása / a glikolízis zárólépése során / 1 pont
- és a citromsavciklus folyamatában keletkezik CO2. 1pont
2.
- A légköri CO2 a hajtás bőrszövetének 1 pont
- gázcserenyílásain át 1 pont
- diffúzióval jut a növénybe. 1 pont
- A szén-dioxidot a fotoszintézis sötét szakaszában / Calvin-ciklusban 1 pont
- A NADPH(2) / hidrogénszállító molekula hidrogénjeivel 1 pont
- az ATP energiájának felhasználásával redukálja szőlőcukorrá / szerves anyaggá a növényi sejt. 1 pont
3.
- A glükózból keményítő 1 pont
- és cellulóz képződik. 1 pont
2017 október
X.A. A szinapszisok – esszé 10 pont
Esszéjében fogalmazza meg a kémiai szinapszisok működésének néhány jellegzetességét!
A következőkre térjen ki:
1. A serkentő és a gátló szinapszis hatása a szinapszis utáni idegsejt polaritásának mértékére, az ezt létrehozó mechanizmusok. (3 pont)
2. A működés energiaigényének, egyirányúságának és az ingerületátadás időigényének magyarázata. (3 pont)
3. A drogok lehetséges támadáspontja, hatásuk lehetséges módja (két példa). A drogtolerancia (hozzászokás) jelensége és lehetséges magyarázata. (4 pont)
X.B. Veszélyforrások - esszé 10 pont
Foglalja össze a hazai sziklagyepeinket fenyegető veszélyforrásokat, a védelem lehetőségeit!
Az alábbiakra térjen ki:
1. Mely nemzeti parkjaink vagy hegységeink területén találhatunk sziklagyepeket (egy példa), természetvédelmi jelentőségük indoklása. (2 pont)
2. A tájidegen fajok, például feketefenyő telepítésének lehetséges hatása, a taposás, útépítések, külszíni bányászat következménye, a természetesnél sűrűbb vadállomány fenntartása.(4 pont)
3. A több kisebb védett területre szakítás következtében fellépő beltenyésztés és sodródás (genetikai) lényege, oka, ezek lehetséges hatása a populációk túlélésére. (4 pont)
Javítási-értékelési útmutató
A szinapszisok – esszé 10 pont
1.
- A serkentő szinapszis működése a szinapszis utáni sejtmembrán polaritását csökkenti,
- a gátló szinapszis a polaritást fokozza / a sejtet hiperpolarizálja.
- A serkentést, ill. gátlást ioncsatornák nyitása váltja ki / a membrán ionáteresztő képessége változik. 3 pont
2.
- Az energiaigényt az átvivőanyagok szintézisének / visszavételének energiaigénye magyarázza.
- Az egyirányúság oka: az átvivőanyag a szinapszis előtti sejtből szabadul fel / sok szinapszisban az átvivőanyag-receptorok csak a szinapszis egyik oldalán találhatók.
- A szinaptikus késés oka a szinaptikus résen át történő diffúzió időigénye. 3 pont
3.
- A drogok lehetséges hatása: a jelátvivő anyag működésének / felszabadulásának fokozása/ visszavételének gátlása / receptormódosítás (Bármely két példa, vagy annak konkrét kifejtése: 2 pont).
- Drogtolerancia: ugyanazon drog azonos mennyisége egyre kisebb hatást vált ki / azonos hatás eléréséhez egyre nagyobb drogmennyiség szükséges. 1 pont
- Lehetséges ok: a receptorok számának / a szinapszisok méretének / számának módosulása. 1 pont
Veszélyforrások – esszé 10 pont
1.
- Sziklagyepek a Bükki / Aggteleki / Duna-Ipoly / Balaton-felvidéki nemzeti parkok /Dunántúli- és Északi-középhegység / Mecsek területén (bármelyik példa jó) fordulnak elő. 1 pont
- Meleg- és hidegkedvelő maradványfajok / bennszülött növények / életközösségek lelőhelyei. (Másképpen is megfogalmazható.) 1 pont
2.
- A feketefenyő árnyékolása / lassan bomló avarrétege miatt kiszorítja az itt őshonos fajokat/ egyéb fajok kompetíciója hasonló hatású lehet 1 pont
- a taposás eróziót / degradációt / gyomosodást indít meg, 1 pont
- a külszíni bányászat csökkenti a termőterületet, 1 pont
- a túltartott vadállomány túrásával / rágásával károsít. 1 pont
3.
- A beltenyésztés a rokon egyedek egymás közti szaporodása, 1 pont
- ami az öröklődő recesszív rendellenességek megjelenésének kockázatát növeli, 1 pont
- A sodródás a kis populációk genetikai változatosságát véletlenszerűen csökkenti, 1 pont
- mindkettő csökkenti a populációk túlélési esélyét. 1 pont
2017 május
IX.A. A reflexek – esszé 10 pont
Mutassa be az emberi szervezet reflexeit az alábbi szempontok figyelembevételével!
1. A feltétlen és a feltételes reflex különbsége. Pavlov vizsgálati módszerének lényege. (3 pont)
2. A vegetatív és a szomatikus reflexívek felépítésének különbsége, az eltérő hatású vegetatív dúcok lehetséges helyei. (4 pont)
3. A vegetatív reflex bemutatása a testhőmérséklet szabályozásának példáján: a szabályozó központ helye (1 pont), két változás, ami a külső hőmérséklet csökkenésekor a szabályozás során végbemegy. (2 pont)
IX.B. Az erdő mintázatai – esszé 10 pont
Mutassa be egy hazai erdő tér-és időbeli mintázatait a következő példák segítségével:
1. Mik az aszpektusok, az aszpektusváltások oka és hatásai két példán bemutatva. (4 pont)
2. A szintezettség két lehetséges okának megnevezése egy erdőn belül. (2 pont)
3. A talaj humusztartalmának eredete, a növények életében játszott szerepe, a lebontásában részt vevő élőlények és szerepük. (4 pont)
Javítási-értékelési útmutató
A reflexek – esszé 10 pont
1.
- A feltétlen reflexek öröklöttek / nem tanultak, 1 pont
- a feltételes reflex egy feltétlen és egy korábban közömbös inger hatásának összekapcsolása / ingertársítás / két reflex összekapcsolása útján jön létre. 1 pont
- Pavlov módszerének leírása (ingertársítás). 1 pont
2.
- A vegetatív reflexek mozgató / végrehajtó idegsejtje a vegetatív dúcban, 1 pont
- a szomatikus reflexeké a központi idegrendszer / gerincvelő szürkeállományában van. 1 pont
(VAGY:
- A vegetatív reflexek simaizomra hatnak. 1 pont
- A szomatikus reflexek harántcsíkolt izomra / vázizomra hatnak. 1 pont)
- A szimpatikus dúcok a (háti-ágyéki) gerincvelő mellett, 1 pont
- a paraszimpatikusok a célszerv közelében (vagy a célszervben) vannak. 1 pont
3.
- A testhőmérsékletet szabályozó központ a hipotalamuszban van. 1 pont
- Lehűléskor e központ fokozza a biológiai oxidációt/ csökkenti a bőr vérellátását/ 1 pont
- csökkenti a verejtékezést / fokozza az izomaktivitást (didergés) /a szőrmerevítő izmok működését („lúdbőr”). 1 pont Bármely két jó hatás leírása.
Az erdő mintázatai – esszé 10 pont
1.
- Az aszpektusok a társulás ciklikusan / rendszeresen visszatérő állapotai, 1 pont
- a váltások fő oka az évszakok váltakozása. 1 pont
- Például ősszel a lombhullató fák leveleinek lehullása, 1 pont
- tavasszal a lombkorona záródása előtti gumós lágyszárúak virágzása (kora tavaszi aszpektus pl. a gyertyános-tölgyesekben). 1 pont
(Bármely két jó példa alkalmas.)
2.
- A szintezettség a fény / a páratartalom / az ökológiai kölcsönhatásban részt vevő partnerek / a szél / a hőmérséklet függőleges irányú egyenlőtlen eloszlásának következménye. 2 pont
(Bármely két jó környezeti tényező megnevezése.)
3.
- A humusz az elpusztult élőlények/maradványok szerves anyagaiból képződik,1 pont
- szemcséi / részecskéi felszínén víz és ásványi sók kötődnek meg, / lebontása után tápanyagforrás 1 pont
- a talajt morzsalékossá, levegőssé teszi. 1 pont
- A lebontók, pl. gombák, baktériumok alakítják vissza szervetlen anyaggá. 1 pont
Témakörök szerinti esszégyűjtemény
AZ EMBERI SZERVEZET
4.3. A mozgás
2025 október
XI.A. Mozgásérzékelés – esszé 10 pont
Esszéjét összefüggő és logikus mondatokban a következő szempontok alapján építse fel!
1. Mutassa be a belső fül szerepét a testhelyzet és a testmozgás érzékelésében! (4 pont)
2. Ismertesse az izomorsók funkcióját, a térdreflexben részt vevő idegsejtek kapcsolódását, működését! Magyarázza meg, hogy miért van szerepe ennek a reflexnek a testérző rendszer működésében, ha alapvetően ez egy gerincvelői reflex! (6 pont)
Javítási-értékelési útmutató
Mozgásérzékelés – esszé 10 pont
Minden helyes válasz 1 pont.
1.
- A félkörös ívjáratokban a folyadék elmozdulása
- a fej gyorsuló mozgását / szöggyorsulását,
- a tömlőcskékben és a zsákocskákban levő receptorok (szőrsejtek) segítségével
- a fej statikus helyzetét / egyenletes lineáris gyorsulását érzékeli a szervezet.
2.
- Az izomorsók a harántcsíkolt (váz)izmok megnyúlását érzékelik.
- A combfeszítő izom megnyúlásakor az izomorsó ingerületi állapota átterjed az érzőidegsejtre.
- A gerincvelőben az érzőidegsejt axonja kettéágazik. Egyik nyúlványa a mozgatóidegsejten keresztül összehúzódásra készteti a combfeszítő izmot,
- másik nyúlványa a combhajlító izom összehúzódását gátolja.
- A reflexív egyik leágazása a testrész helyzetéről információt szállít
- a nagyagyi érzőkéregbe / a nagyagyba / az (elsődleges) érzőkéregbe.
4.4. A táplálkozás
2024 október
IX.A. A máj sokrétű funkciói – esszé 10 pont
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra!
1. Az epesavak mely kémiai jellemzője biztosítja az emulgeáló funkciót, és mely egyéb feladata van még az epesavaknak? Miért előnyös az emésztés szempontjából az emulgeálás? (3 pont)
2. Mely, a vizeletbe kerülő vegyületet képes előállítani a máj az aminosavak lebontása során? Mely aminosavakat nem képes más aminosavakból előállítani? (2 pont)
3. Mely szénhidrát formájában raktározza a máj a glükóz feleslegét? Elsősorban mely hormon szabályozza a szintézis mértékét? (2 pont)
4. A hemoglobin lebontását is a máj végzi. Hova kerülnek és hogyan ürülnek a szervezetből a hemoglobin lebontási termékei abban az esetben, ha a máj egészséges, és mi történik velük, ha a máj súlyosan károsodott? Mely látható tünet jelzi ezt? (3 pont)
Javítási-értékelési útmutató
A máj sokrétű funkciói – esszé 10 pont
1.
- Az epesavak kettős oldékonyságú (amfipatikus) molekulák / képesek poláris és apoláris anyagokban is oldódni, így tudják emulgeálni a zsírokat. 1 pont
- További funkciójuk, hogy aktiválják a lipázokat (zsírbontó enzimek) / stabilizálják a lipidemulziót. 1 pont
- Az emulgeálás révén a lipázok nagyobb felületen tudnak hozzáférni a lipidekhez, így hatékonyabban tudják azokat lebontani. 1 pont
2.
- A máj az aminosavak lebontása során karbamidot hoz létre, 1 pont
- az esszenciális aminosavat nem tudja állítani (megfelelő mennyiségben). 1 pont
3.
- A glükózt glikogén formájában raktározza 1 pont
- ennek mértékét elsősorban az inzulin / glukagon szintje szabályozza. 1 pont
4.
- A hemoglobin lebontási termékeit (biliverdin, bilirubin) a máj bejuttatja az epébe / vékonybélbe epefestékként, azok a széklettel / vizelettel ürülnek. 1 pont
- Ha a máj súlyosan károsodott, akkor ezek a lebontási termékek felhalmozódnak a vérben / bejutnak a bőrbe és a szem ínhártyájába, 1 pont
- emiatt a májelégtelenség sárgasághoz vezet. (Hasi ödémát is okozhat.) 1 pont
A májelégtelenség más helytálló következménye is elfogadható.
4.5. A légzés
4.6. Az anyagszállítás
2022 május
X.A. A vörösvértestek – esszé 10 pont
Mutassa be a vörösvértestek felépítését és szerepét! Fogalmazásában térjen ki az alábbi vázlatpontok mindegyikére!
1. A vörösvértestek felépítésének jellegzetességei (színének magyarázata, alakja). (2 pont)
2. A membránjukon található jellegzetes antigének megnevezése, jelentőségük a vérátömlesztésben. (2 pont)
3. A vörösvértestek feladata, magashegyi edzőtáborok hatása a vörösvértestek számára, hatása a sportteljesítményre, indoklással. (3 pont)
4. A vérszegénység három, elvileg különböző, lehetséges oka. (3 pont)
Javítási-értékelési útmutató
A vörösvértestek – esszé 10 pont
1.
- Vörös színüket a sejtplazmában elhelyezkedő hemoglobin adja, 1 pont
- alakjuk fánk/ korong / súlyzó. 1 pont
2.
- Membránjukon vércsoportantigének / A, B, D, 0 antigének helyezkedhetnek el. 1 pont
- Vérátömlesztésnél/transzfúziónál ügyelni kell a csoportazonos vér adására. 1 pont
3.
- A vörösvértestek feladata a légzési gázok/ oxigén és szén-dioxid szállítása. 1 pont
- A magas hegységekben (az oxigén alacsony parciális nyomására) a szervezet fokozott mennyiségű vörösvértest-előállítással válaszol, 1 pont
- az alacsonyabb szintre visszaérkezve a megemelkedett vörösvértest-szám nagyobb teljesítményt tesz lehetővé/ hatékonyabb oxigénkötést biztosít, ezért tartanak edzőtáborokat a hegységekben. 1 pont
4.
- A vérszegénység oka lehet vashiány - a vas elégtelen bevitele vagy felszívódása/ vitaminhiány / B12 vitamin/folsav hiánya, / leukémia következménye nagyfokú vérveszteség / krónikus betegség / a hemoglobin örökletes betegségei / vérképzés zavara. / Rézanyagcsere zavara
1-1 pont, 3 különböző ok említése összesen: 3 pont
4.7. A kiválasztás
2021 május
IX.A. A víz távozása a szervezetből – esszé 10 pont
Írjon fogalmazást arról, milyen folyamatok eredményeképpen távozik víz a testünkből!
Esszéjében az alábbiakra térjen ki!
1. A szűrlet fogalma, keletkezésének magyarázata (a nyomáskülönbség oka) és helye a nefronban. (3 pont)
2. A vízvisszaszívást biztosító fizikai-kémiai transzportmechanizmus. Mely anyagok koncentrációkülönbsége biztosítja az ehhez szükséges hajtóerőt? (3 pont)
3. A vizelet útja a veséktől a külvilágig. (1 pont)
4. A vizelet napi térfogatának növekedését, illetve csökkenését okozó élettani helyzetek egészséges emberben. (1-1 példa) (2 pont)
5. Két példa olyan egészséges folyamatra, amely a vizeletürítésen kívül jelentősebb mennyiségű vizet távolít el a szervezetünkből. (1 pont)
Javítási-értékelési útmutató
A víz távozása a szervezetből – esszé 10 pont
1.
- A szűrlet fehérjementes vérplazma 1 pont
- a vesetestecskében / Malpighi-testecskében keletkezik, 1 pont
- a vérnyomáskülönbség hatására: utalás a bemenő és kilépő artériák átmérőjének különbségére. 1 pont
2.
- a víz diffúzióval / ozmózissal / passzív transzporttal szívódik vissza (a közeli elsődleges kanyarulatos csatornát kísérő hajszálerekben) a szűrletképzést követően 1 pont
- (relatíve) kevés víz, de (csaknem) változatlan anyagmennyiségű fehérje marad, emiatt magas a vér (kolloid) ozmózisnyomása, 1 pont
- (a Henle-kacs területén) aktív transzport / pumpamechanizmusok / ionpumpák növelik meg a vér Na+-ion koncentrációját / ozmózisnyomását 1 pont
3.
- (vesemedence/vesemedencék) – húgyvezeték(ek) – húgyhólyag – húgycső (együtt) 1pont
4.
- növeli: a szükségesnél nagyobb mennyiségű folyadék fogyasztása/vízhajtó hatású italok (kávé, tea, alkohol stb.) 1 pont
- csökkenti: a szükségesnél kisebb mennyiségű folyadék fogyasztása / fokozott verejtékezés / menstruáció előtti vízvisszatartás / erősen sós táplálék fogyasztása 1 pont
5.
- verejtékezés / levegő kilégzése / székletürítés (bármely kettő) 1 pont
4.8. A szabályozás, érzékszervek
2025 május
IX.A. Elemi idegjelenségek – esszé 10 pont
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra! Esszéjét a 19. oldalon írhatja meg!
1. Ismertesse, hogy a ligandfüggő és a feszültségfüggő ioncsatornák az idegsejt sejthártyájának jellemzően melyik részén találhatók meg! Adja meg, hogy mi váltja ki szerkezetük megváltozását, és működésüknek köszönhetően hogyan változhat meg a membránfeszültség! (4 pont)
2. Ismertesse, hogy mely ionok milyen irányú transzportjának köszönhető a membránpotenciál növekedése, illetve csökkenése! (2 pont)
3. Foglalja össze, hogy az ingerületvezetés az idegsejt melyik területén történik analóg módon, melyik részén digitálisan, és mi módon kódolja ez a kétféle jel az inger erősségét! (4 pont)
Javítási-értékelési útmutató
Elemi idegjelenségek – esszé 10 pont
1.
- A ligandfüggő ioncsatornák a dendrit / sejttest sejthártyáján / a posztszinaptikus membránon helyezkednek el. 1 pont
- A feszültségfüggő ioncsatornák az axon eredési dombján / az axon sejthártyáján / az idegrost befűződésén / az axon végbunkóján találhatók meg. 1 pont
- A ligandfüggő ioncsatornák szerkezete a (fehérjemolekulához) kapcsolódó ligand / ingerület átvivő anyag hatására változik meg, és ennek következtében helyi áram / hipo-, illetve hiperpolarizáció alakul ki. 1 pont
- A feszültségfüggő ioncsatornák szerkezete egy adott membránpotenciál / küszöbpotenciál kialakulása esetén változik meg, és ennek hatására (tovaterjedő) akciós potenciál alakul ki. 1 pont
2.
- A membránpotenciál csökkenése / hipopolarizációja a Na+ felvétele miatt alakul ki. 1 pont
- A membránpotenciál növekedése / hiperpolarizációja a Cl– felvétele / K+ leadása miatt alakul ki. 1 pont
3.
- Analóg módon a sejttesten / dendriteken / csupasz axonokon 1 pont
- digitálisan az (velőshüvelyes) axonon / idegroston történik az ingerületvezetés / kémiai szinpaszisban (a „minden vagy semmi” törvény alapján). 1 pont
- Az analóg jel a de- / hipo- / hiperpolarizáció / a membránpotenciálváltozás nagyságán keresztül kódolja az inger erősségét / nagyságát. 1 pont
- A digitális jel az akciós potenciálok frekvenciáján / időbeli gyakoriságán keresztül kódolja az inger erősségét / nagyságát. 1 pont
2022 október
IX.A. Szembetegségek – esszé 10 pont
Írjon rövid esszét a látást veszélyeztető felsorolt megbetegedésekről! Fogalmazásában térjen ki az alábbi szempontokra:
1. A rövid- (közel)látás fizikai oka, következménye, a nem műtéti kezelési lehetősége. (3 pont)
2. A vörös-zöld színtévesztés öröklődésének jellemzői, megoszlása a nemek között. (3 pont)
3. A zöldhályog és a szürkehályog oka, a látásra gyakorolt hatásuk. (4 pont)
Javítási-értékelési útmutató
Szembetegségek – esszé 10 pont
1.
- A rövid- / közellátó szemben az éles kép a retina előtt keletkezik / a szemtengely hosszabb, mint az éles képalkotáshoz szükséges / a szem fénytörő képessége a szükségesnél nagyobb 1 pont
- ezért a távoli tárgy képe homályos lesz. 1 pont
- Szóró / homorú / konkáv lencsével korrigálható. 1 pont
2.
- A vörös-zöld színtévesztés recesszíven, 1 pont
- X-kromoszómához kötötten öröklődik 1 pont
- ezért / (a hemizigóta öröklés miatt) a férfiakban gyakoribb. 1 pont
3.
- A zöldhályog esetében csarnokvíz felszaporodik / a magas nyomás miatt változik a szaruhártya fénytörése, 1 pont
- (a látóideg pusztulása miatt) látásromlás / látótérkiesés / vakság alakulhat ki. 1 pont
- A szürkehályog a szemlencse fényáteresztő-képességének csökkenése, 1 pont
- emiatt elhomályosul a látás, csökken a szem fényérzékenysége. 1 pont
2019 október
IX.A. A retina és a látás – esszé 10 pont
Esszéjében mutassa be az éles látás és a távolságbecslés fizikai-biológiai hátterét!
A következőkre térjen ki!
1. Jellemezze a retinán kialakuló kép fizikai tulajdonságait (nagyság, helyzet)! (2 pont)
2. Magyarázza, hogy a retina melyik területén a legélesebb a látás! (2 pont)
3. Mutassa be, hogy mely izom segítségével hogyan alkalmazkodik a szem, amikor távoli tárgyról közelire pillantunk! (3 pont)
4. Mutasson be három észlelési módot, ami a látás során lehetőséget ad a megfigyelt tárgyak távolságának becslésére, érzékelésére! (3 pont)
Javítási-értékelési útmutató
A retina és a látás – esszé 10 pont
1.
- A retinára kicsinyített, 1 pont
- fordított állású kép vetül. 1 pont
2.
- A sárgafoltban a legélesebb a látás, 1 pont
- mert a csapok itt állnak a legsűrűbben / itt csak csapok vannak. 1 pont
3.
Közelre nézéskor
- a sugártest izmai megfeszülnek, a sugártest bedomborodik, 1 pont
- a lencsefüggesztő rostok ellazulnak, 1 pont
- a szemlencse domborúbbá válik / fókusztávolsága csökken/fénytörése nő. 1 pont
4.
A távolságról információt nyújt
- a szemtengelyek állása / a külső szemmozgató izmok / a tárgyak takarása (a közelebbi takarja a távolabbit), / az ismert nagyságú tárgyak retinára vetülő képének nagysága / perspektivikus rövidülés / mozgás során az elmozdulás látható gyorsasága /a távolabbi tárgyak színének módosulása.
Bármelyik három leírása: 1+1+1 = 3 pont
2019 május
IX.A. Az agyalapi mirigy – esszé 10 pont
Ismertesse az agyalapi mirigy működésének megváltozását egy benne képződő hormontermelő daganat következtében! Az alábbiakra térjen ki:
1. Hogyan befolyásolja a daganat a pajzsmirigy működését? Milyen hatása van a beteg anyagcseréjére, testhőmérsékletére, testsúlyára és idegrendszeri működésére? (5 pont)
2. Milyen hatása van az elváltozásnak a mellékvese működésére? (3 pont)
3. Milyen következményekkel járhat együtt a növekedési hormon túltermelése fiatal- és felnőtt korban? (2 pont)
Javítási-értékelési útmutató
Az agyalapi mirigy – esszé 10 pont
1.
- A daganat hatására fokozódik a pajzsmirigyben a hormontermelés / a tiroxin elválasztása / leadása, 1 pont
- aminek hatására az anyagcsere felgyorsul / a lebontó anyagcsere aránya nő, 1 pont
- a (normál) testhőmérséklet megemelkedik, 1 pont
- az idegrendszer aktivitása fokozódik / a betegre idegesség / alvászavar jellemző / ingerküszöbe csökken 1 pont
- (éhségérzet / sok táplálék ellenére) testsúlya csökken. 1 pont
2.
- A mellékvesekéreg-serkentő hormon termelésének fokozódása miatt
- a mellékvese kéregállományának működése is fokozódik,
- elsődlegesen a szénhidrátanyagcserére ható hormonok / glükokortikoidok elválasztásának fokozása
- a szénhidrátanyagcsere zavarát / a glikogénraktárak kiürülését okozza / megemelkedő vércukorszintet okoz / befolyásolja (gátolja) az immunrendszert.
- Részben a mineralokortikoidok / szexuálszteroidok elválasztása is fokozódik.
Bármely három következmény megfogalmazása 1+1+1 = 3 pont
Ha a vizsgázó hibás előfeltevésből (a gátlásból) indult ki, de a továbbiakban az oksági összefüggéseket helyesen elemezte, a gondolatmenet első 1-1 pontját leszámítva értékelhető a válasza, ami maximálisan 4+2 = 6 pont.
3.
A növekedési hormon túltermelése
- fiatal korban óriásnövést 1 pont
- felnőttkorban a csúcsi szervek / fül, orr nyelv megnagyobbodását / akromegáliát okoz. 1 pont
2018 október
IX.A. Az agyalapi mirigy hormontermelése – esszé 10 pont
Írjon rövid esszét az agyalapi mirigy hormontermeléséről! Fogalmazásában térjen ki az alábbi szempontokra:
1. Az agynak pontosan melyik részével áll közvetlen működési kapcsolatban az agyalapi mirigy? Röviden írja le működési kapcsolataik lényegét! (2 pont)
2. A hátsó lebenyhez köthető hormonok neve, hatásai. (4 pont)
3. Melyek a növekedési hormon hiányának és túltermelődésének következményei? (2 pont)
4. A pajzsmirigy hormontermelését közvetlenül szabályozó hormon működésében mely szabályozási mechanizmus hogyan valósul meg? (2 pont)
Javítási-értékelési útmutató
Az agyalapi mirigy hormontermelése – esszé 10 pont
1.
- Az agyalapi mirigy a hipotalamuszhoz kapcsolódik. 1 pont
- A hipotalamusz termeli a hátsó lebeny hormonjait és szabályozza az elülső lebeny hormontermelését. 1 pont
2.
- A hátsó lebenyből ürül az oxitocin, 1 pont
- ami simaizom-összehúzódást okoz a méhfalban / emlőmirigyekben, 1 pont
- illetve a vazopresszin / ADH, 1 pont
- ami fokozza a víz visszaszívását a szűrletből. 1 pont
(A mechanizmusra vonatkozó megfogalmazás is helyes: víz számára átjárhatóvá teszi a nefron / elvezető csatorna / gyűjtőcsatorna falát.)
3.
- Hiányában arányos törpenövés, 1 pont
- magas szintje esetén óriásnövés /gigantizmus / akromegália alakul ki. 1 pont
4.
- A (pajzsmirigyserkentő) hormon termelését a tiroxin gátolja, ami a tiroxinszintjének beállítását eredményezi. (Másként is megfogalmazható.) 1 pont
- Ez negatív visszacsatolás 1 pont
2017 október
X.A. A szinapszisok – esszé 10 pont
Esszéjében fogalmazza meg a kémiai szinapszisok működésének néhány jellegzetességét!
A következőkre térjen ki:
1. A serkentő és a gátló szinapszis hatása a szinapszis utáni idegsejt polaritásának mértékére, az ezt létrehozó mechanizmusok. (3 pont)
2. A működés energiaigényének, egyirányúságának és az ingerületátadás időigényének magyarázata. (3 pont)
3. A drogok lehetséges támadáspontja, hatásuk lehetséges módja (két példa). A drogtolerancia (hozzászokás) jelensége és lehetséges magyarázata. (4 pont)
Javítási-értékelési útmutató
A szinapszisok – esszé 10 pont
1.
- A serkentő szinapszis működése a szinapszis utáni sejtmembrán polaritását csökkenti,
- a gátló szinapszis a polaritást fokozza / a sejtet hiperpolarizálja.
- A serkentést, ill. gátlást ioncsatornák nyitása váltja ki / a membrán ionáteresztő képessége változik. 3 pont
2.
- Az energiaigényt az átvivőanyagok szintézisének / visszavételének energiaigénye magyarázza.
- Az egyirányúság oka: az átvivőanyag a szinapszis előtti sejtből szabadul fel / sok szinapszisban az átvivőanyag-receptorok csak a szinapszis egyik oldalán találhatók.
- A szinaptikus késés oka a szinaptikus résen át történő diffúzió időigénye. 3 pont
3.
- A drogok lehetséges hatása: a jelátvivő anyag működésének / felszabadulásának fokozása/ visszavételének gátlása / receptormódosítás (Bármely két példa, vagy annak konkrét kifejtése: 2 pont).
- Drogtolerancia: ugyanazon drog azonos mennyisége egyre kisebb hatást vált ki / azonos hatás eléréséhez egyre nagyobb drogmennyiség szükséges. 1 pont
- Lehetséges ok: a receptorok számának / a szinapszisok méretének / számának módosulása. 1 pont
2017 május
IX.A. A reflexek – esszé 10 pont
Mutassa be az emberi szervezet reflexeit az alábbi szempontok figyelembevételével!
1. A feltétlen és a feltételes reflex különbsége. Pavlov vizsgálati módszerének lényege. (3 pont)
2. A vegetatív és a szomatikus reflexívek felépítésének különbsége, az eltérő hatású vegetatív dúcok lehetséges helyei. (4 pont)
3. A vegetatív reflex bemutatása a testhőmérséklet szabályozásának példáján: a szabályozó központ helye (1 pont), két változás, ami a külső hőmérséklet csökkenésekor a szabályozás során végbemegy. (2 pont)
Javítási-értékelési útmutató
A reflexek – esszé 10 pont
1.
- A feltétlen reflexek öröklöttek / nem tanultak, 1 pont
- a feltételes reflex egy feltétlen és egy korábban közömbös inger hatásának összekapcsolása / ingertársítás / két reflex összekapcsolása útján jön létre. 1 pont
- Pavlov módszerének leírása (ingertársítás). 1 pont
2.
- A vegetatív reflexek mozgató / végrehajtó idegsejtje a vegetatív dúcban, 1 pont
- a szomatikus reflexeké a központi idegrendszer / gerincvelő szürkeállományában van. 1 pont
(VAGY:
- A vegetatív reflexek simaizomra hatnak. 1 pont
- A szomatikus reflexek harántcsíkolt izomra / vázizomra hatnak. 1 pont)
- A szimpatikus dúcok a (háti-ágyéki) gerincvelő mellett, 1 pont
- a paraszimpatikusok a célszerv közelében (vagy a célszervben) vannak. 1 pont
3.
- A testhőmérsékletet szabályozó központ a hipotalamuszban van. 1 pont
- Lehűléskor e központ fokozza a biológiai oxidációt/ csökkenti a bőr vérellátását/ 1 pont
- csökkenti a verejtékezést / fokozza az izomaktivitást (didergés) /a szőrmerevítő izmok működését („lúdbőr”). 1 pont Bármely két jó hatás leírása.
4.9. Szaporodás és egyedfejlődés
2024 május
IX.A. Kétivarúság és várandósság – esszé 10 pont
Írjon esszét a nemi jellegekért felelős hormonokról, a megtermékenyítésről és az ikerterhességről az alábbi vázlatpontok alapján!
1. A kromoszómális nem definiálása. (1 pont)
2. A másodlagos női, illetve hím ivari jellegek kialakításért felelős hormonok neve, termelődésük helye és időbeli jellege. Mely életszakaszban (életkorban/mettől meddig) termelődnek a női, illetve a hím ivari hormonok a születés utáni élet során? (4 pont)
3. A megtermékenyítés helye és lehetséges időpontja a női nemi ciklus során. (2 pont)
4. Az egypetéjű és kétpetéjű ikrek kialakulása, genetikai információtartalmuk összehasonlítása egymással. (3 pont)
Javítási-értékelési útmutató
Kétivarúság és várandósság – esszé 10 pont
1.
- A kromoszomális nem a testi sejtek ivari kromoszómakészlete, XX: nő, XY: férfi. 1 pont
2.
- Ösztrogén – petefészkek / tüszők termelik, 1 pont
- tesztoszteron – herék termelik. 1 pont
- Az ösztrogén ciklikusan, a tesztoszteron folyamatosan termelődik. 1 pont
- Az ösztrogén az ivaréréstől a menopauzáig, a tesztoszteron ivaréréstől élethossziglan termelődik. 1 pont
3.
- A megtermékenyítés a petevezetékben / a petevezeték felső harmadában, 1 pont
- a ciklus 14–15. napján/ az ovulációt követő 12–24 órában megy végbe. 1 pont
4.
- Egypetéjű ikrek: az embrió néhány sejtes állapotban kettéválik. 1 pont
- Kétpetéjű ikrek: egyszerre /időben egymáshoz közel két petesejt termékenyül meg. 1 pont
- Az egypetéjű ikrek genetikai információja azonos (ha eltekintünk a mutációktól),
a kétpetéjűeké csak annyira hasonlít, mint a különböző időpontban született testvéreké./ átlagosan felerészben hasonló. 1 pont
Más hasonló tartalmú megfogalmazás is elfogadható.
2021 október
IX.A. A női nemi ciklus második szakasza – esszé 10 pont
Mutassa be a női nemi ciklus második szakaszának eseményeit! Esszéjében az alábbi pontokra térjen ki:
1. Melyik hipofízis (agyalapi mirigy) hormon melyik női nemi hormon termelését kezdi jelentősen fokozni ebben a szakaszban? Mi termeli ezt a hormont a petefészekben és mi a fő hatása a méh állapotára? (4 pont)
2. Írja le a negatív visszacsatolási folyamatot és az általa beindított eseménysort, amely a ciklus befejezéséhez vezet, ha a petesejt nem termékenyül meg! (4 pont)
3. Mely hormonok teszik lehetővé, illetve jelzik a várandós állapotot az embrió / csíra beágyazódása esetén? (2 pont)
Javítási-értékelési útmutató
A női nemi ciklus második szakasza – esszé 10 pont
1.
- A hipofízisben termelődő sárgatestserkentő (LH) hormon 1 pont
- hatására a sárgatestben termelt 1 pont
- progeszteron 1 pont
- fenntartja a méh nyálkahártyájának megvastagodott állapotát. 1 pont
2.
- A magas progeszteronszint gátló / negatív visszacsatoló hatása gátolja az LH termelését, 1 pont
- ami a sárgatest elsorvadásához vezet, melynek hatására 1 pont
- a progeszteronszint lecsökken, 1 pont
- ami kiváltja a menstruációt. 1 pont
3.
- Beágyazódás esetén a progeszteronszint magas marad, 1 pont
- (a vérben / vizeletben az embrió, később a méhlepény által termelt) terhességi hormon / hCG jelenik meg. 1 pont
2020 október
IX.A. A menstruációs ciklus – esszé 10 pont
Ismertesse a menstruációs ciklus történéseit! Esszéjében az alábbi szempontokat kövesse:
1. Az agyalapi mirigy hormonjai, amelyek a menstruációs ciklust befolyásolják – fő hatásukkal. (5 pont)
2. A tüszőérés és a sárgatest kialakulása, szerepe a ciklus során. (2 pont)
3. A méhnyálkahártya változásai a női nemi ciklus során. a menstruáció hormonális oka. A petefészek hormonjainak hatása a méhnyálkahártya állapotára. (3 pont)
Javítási-értékelési útmutató
A menstruációs ciklus 10 pont
1.
- Az agyalapi mirigy tüszőserkentő hormonja megindítja a tüszőérést. 1 pont
- A sárgatestserkentő hormon(nal együtt) kiváltja a tüszőrepedést. 1 pont
- A ciklus közepén bekövetkezik az ovuláció / tüszőrepedés, ekkor 1 pont
- a petesejt kiszabadul a petefészekből és elindul a petevezetőben. 1 pont
2.
- A visszamaradó tüsző sárgatestté alakul, 1 pont
- amely progeszteron hormont / sárgatest-hormont termel a ciklus második felében. 1 pont
3.
- A progeszteron szint csökkenése váltja ki a menstruációt. 1 pont
- A méh nyálkahártyája a menstruációkor vérzés kíséretében leválik. 1 pont
- Ösztrogén hatására a ciklus első felében újraképződik. 1 pont
- A ciklus második felében progeszteron hatására tovább vastagodik, mirigyekben gazdagodik. 1 pont
Egyéb
2023 október
IX.A. Fehérjefunkciók az emberben – esszé 10 pont
Mutassa be a fehérjemolekulák különféle funkcióit az emberi szervezetben, a következő fehérjemolekulák példáján:
1. aktin és miozin;
2. amiláz;
3. keratin;
4. fibrinogén;
5. vazopresszin/antidiuretikus hormon.
Minden egyes fehérje esetében térjen ki arra, hogy mely szerv(ek)ben termelődik a szervezetben, és mi a funkciója!
Javítási-értékelési útmutató
Fehérjefunkciók az emberben 10 pont
1. aktin és miozin:
- Az izomszövetekben található filamentumok, 1 pont
- az izomösszehúzódást teszik lehetővé. 1 pont
2.amiláz:
- Termelik a nyálmirigyek és a hasnyálmirigy, 1 pont
- keményítőt / poliszacharidokat / oligoszacharidokat emészt a tápcsatornában. 1 pont
3. keratin:
- A bőrben, 1 pont
- mechanikai / kiszáradás elleni védelem. 1 pont
4. fibrinogén:
- A máj termeli, 1 pont
- a véralvadás fehérjéje / fibrinné alakul, majd hálót képez. 1 pont
5. vazopresszin/antidiuretikus hormon:
- A hipotalamusz termeli, 1 pont
- a víz visszaszívását serkenti a vesében. 1 pont
2023 május
IX.A. A kalcium szerepe az ember szervezetében – esszé (10 pont)
Az esszében térjen ki az alábbiakra!
1. A vázizmok működésében nélkülözhetetlenek a Ca2+-ionok. Írja le a harántcsíkolt izmok működésének folyamatát az alábbi szempontok figyelembevételével!
A Ca2+-ionok kötődése, ennek hatása a folyamatban résztvevő fehérjékre, az energiafelhasználás módja, a sötét és világos sávok szélességének változása és ennek magyarázata. (4 pont)
2. A Ca2+-ionok nélkülözhetetlenek a véralvadás folyamatában is. Írja le azokat a protrombinból kiinduló eseményeket, amelyek fibrinháló kialakulásához vezetnek! (2 pont)
3. A kalciumnak a csontok felépítésében is fontos szerepe van. A csontszövet mely alkotórészében, mely sók formájában (egy példa elég) fordulnak elő a Ca2+-ionok, s milyen tulajdonságát adják a csontoknak? (3 pont)
4. Nevezze meg azt a vitamint, amely elősegíti a Ca2+-ionok felszívódását! (1 pont)
Javítási-értékelési útmutató
A kalcium szerepe az ember szervezetében – esszé 10 pont
1.
- a Ca2+-ionok hatására a miozin az aktinszálhoz (troponinhoz) kötődik 1 pont
- az aktinszálak egymás felé / a miozinfonalak közé csúsznak 1 pont
- ennek következtében a világos és sötét sávok (egymáshoz viszonyított) aránya változik 1 pont
- a folyamat ATP-igényes / a kalcium-ionok visszajutása is energiaigényes 1 pont
2.
- protrombinból trombin lesz 1 pont
- a trombin katalizálja azt a folyamatot, amelynek során az oldott állapotú fibrinogénből oldhatatlan fibrinszálak lesznek (majd ezekből alakul ki a fibrinháló) 1 pont
3.
- sejtközötti állományban 1 pont
- Ca-foszfát / karbonát / fluorid / (hidroxiapatit) 1 pont
- szilárdságát / keménységét / színét 1 pont
4.
- D-vitamin 1 pont
2020 május
IX.A. A baktériumtranszformáció és a gyógyítás – esszé 10 pont
Esszéjében írja le, hogy Griffith és Avery kísérletében (baktériumtranszformáció) milyen eredményre jutottak, és felfedezésüknek mi a jelentősége a gyógyításban!
Esszéjében az alábbiakra térjen ki:
1. Milyen módszerrel mit igazolt Griffith, és mit bizonyított Avery kutatócsoportja? Mit igazolt a két kutató kísérletsorozata a prokarióták információátadásáról? (4 pont)
2. Mit értünk a génmódosított (GMO) élőlények létrehozásán? Hozzon egy példát a genetikailag módosított élőlények hasznosítására a gyógyászatban! (2 pont)
3. Mutassa be egy bakteriális eredetű, gyulladásos betegség jellemző tüneteit! Induljon ki a gyulladás általános jellemzőiből és alkalmazza ezt a tüdőre vonatkozóan! (2 ok – 2 következmény) (4 pont)
Javítási-értékelési útmutató
A baktériumtranszformáció és a gyógyítás – esszé 10 pont
1.
- Módszer: A kutatók tokképző, illetve tok nélküli / (S és R változatú) (tüdőgyulladás) baktériumokkal (Pneumococcusokkal) fertőztek egereket: a tokképző volt a halálos. A hővel elölt tokképző nem okozott betegséget, de a hővel elölt tokképző és a tok nélküli keverékben létrejöttek a tokos változatok (baktériumtranszformáció). 1 pont
- A tokképződésre vonatkozó információ átjuthat egyik sejtről a másikra (Griffith kísérletei) 1 pont
- Módszer: A keverékhez adott fehérje- vagy RNS-bontó enzimek nem gátolták a transzformációt, a DNS-bontó enzimek azonban igen. 1 pont
- Bizonyítás: tehát a tokképzésre vonatkozó információt a DNS vitte át a kísérlet során. (Averyék eredménye) 1 pont
2.
- A genetikai módosítás egy vagy több gén (transzgén) szándékos / célzott bejuttatása egy másik faj egyedeibe / gyakran eukarióta és prokarióta között megy végbe. 1 pont
- Génmódosított baktériummal pl. lehetséges inzulint előállítani, ami lehetővé teszi a cukorbetegség kezelését. 1 pont Más gyógyászati célú példa is elfogadható.
|
2018 május
IX.A. A szőlőcukor útja a szervezetben – esszé 10 pont
Egy gyermek kiflit majszol. A kifli keményítőtartalma glükózzá emésztődik a tápcsatornájában.
Ismertesse a glükóz útját a gyermek szervezetében az alábbiak szerint:
1. A tápcsatorna melyik szakaszában, hová szívódik fel a szőlőcukor? (2 pont)
2. Mely hormon segíti raktározását poliszacharid formában? (1 pont)
3. Melyik biokémiai folyamatban bontják le a glükózt energianyerés céljából a sejtek oxigén jelenlétében és oxigén hiányában? (A folyamatok megnevezése, sejten belüli helyszíne.) Nevezze meg e lebontó folyamatok széntartalmú végtermékeit, és ismertesse azok további útját a szervezetben! (7 pont)
Javítási-értékelési útmutató
A szőlőcukor útja a szervezetben – esszé 10 pont
1.
- A glükóz a vékonybélből 1 pont
- a véráramba / kapillárisokba szívódik fel. 1 pont
2.
- Az inzulin serkenti a raktározást. / Elfogadható a glükokortikoidok hatásának leírása is. 1 pont
3.
- Oxigén jelenlétében a biológiai / terminális oxidációban 1 pont
- a mitokondriumokban, 1 pont
- oxigén hiányában tejsavas erjedésben. 1 pont
- a sejtplazmában bomlik. 1 pont
- Az oxidációban a glükóz széntartalma szén-dioxiddá alakul, 1 pont
- amely a keringés útján a tüdő léghólyagocskáiba / a kilégzés folyamatában a külvilágba kerül. 1 pont
- A tejsav a vérárammal a májba jut. 1 pont
ÖKOLÓGIA, EGYED FELETTI SZERVEZŐDÉSI SZINTEK
2025 október
XI.B. Biológiai sokféleség és regeneráció – esszé 10 pont
Fogalmazza meg a biodiverzitással, annak megóvásával és helyreállításával kapcsolatos összefüggéseket!
Esszéjét összefüggő és logikus mondatokban a következő szempontok alapján építse fel!
1. Definiálja a biodiverzitás következő szintjeit: genetikai diverzitás, fajdiverzitás, ökológiai diverzitás! Melyik diverzitási szint értelmezhető a havasi kecskék esetén, és ennek értéke hogyan változhatott meg a B) szövegrészlet alapján? (4 pont)
B) szövegrészlet: „[…] A Délkelet-alaszkai Egyetem, illetve a kanadai Victoria Egyetem vadbiológusai 17 évig tartó kutatásuk során igazolták, hogy a valóságban rengeteg havasi kecske (2. ábra) pusztul el lavina következtében. […] A lavinák komoly hatást gyakorolnak a kecskék populációdinamikájára. Volt olyan alaszkai populáció, amelyben a meghalt állatok 65 százalékát lavina pusztította el […]. Átlagosan évente az állatok nyolc százaléka pusztul el lavinaomlás következtében, de volt olyan különösen rossz év is, amikor a mortalitás 22 százalékos volt. […] A lavinaomlásban az adatok szerint nemcsak a tapasztalatlan és gyenge gidák pusztulnak el, hanem gyakorlatilag a populációk bármely egyede. Az áldozatok között többségben vannak a legtermékenyebb életszakaszukat élő nőstények […].”
2. Tegyen egy-egy természetvédelmi javaslatot, amelyekkel az A) és a B) szövegrészletben szereplő esetekben megőrizhető, megvédhető a biodiverzitás! (2 pont)
A) szövegrészlet: „2020 januárjában, amikor a világ nagy része még mit sem sejtett a közelgő világjárványról, sok millióan figyelték a soha nem látott vadsággal tomboló ausztrál erdőtüzeket. Egész Délkelet-Ausztrália füstbe merült, a tűzoltók hasztalan próbálták megfékezni a lángokat, amelyek felmérhetetlen kiterjedésű erdőt és állatok sokaságát emésztettek el. A 2019–2020-as „fekete nyár”, ahogy elnevezték a hónapokig tartó katasztrófát, ízelítőt nyújtott abból, ami a klímaváltozás következtében vár a Föld nagy erdőségeire, legyenek azok Kaliforniában, Kanadában vagy Dél-Európában. […] A szakirodalomban számos adatot találtak arról, hogy a legkülönfélébb állatok – nemcsak emlősök, de madarak és hüllők is – rendszeresen használják a vombatok járatait erdőtüzek idején. Ezért a Charles Sturt Egyetem ökológusai elhatározták, hogy szisztematikusabban megvizsgálják a vombatüregek szerepét az erdei ökoszisztémák tűzzel szembeni ellenálló képességében. A terepi vizsgálatokat a Woomargama Nemzeti Parkban végezték, amelynek 311 négyzetkilométernyi területéből nagyjából 180 km2 megsemmisült a 2019–2020-as erdőtüzekben. Amikor egy évvel később az ökológusok elkezdték a terepi felméréseket, azt tapasztalták, hogy a növényzet már látványosan regenerálódni kezdett. […] A vombatok (1. ábra) növényevők, és bár időnként agresszívek lehetnek a territoriális fenyegetést jelentő fajtársaikkal és más állatokkal szemben, még mindig sokkal biztonságosabb a közelségükben átvészelni a tüzet, mint egy ragadozó társaságában. […]”
3. Az A) szövegrészletben leírt erdőtüzeket követően a területek fokozatos regenerációja figyelhető meg. Fogalmazza meg, hogyan változik a regenerálódás során az életközösségekben a K-stratégiájú populációk aránya, a biomassza értéke, a populációs kölcsönhatások száma és a gyomnövényfajok száma! Indokolja, hogy miért adott irányba változnak ezek a tényezők! (4 pont)
Javítási-értékelési útmutató
Biológiai sokféleség és regeneráció – esszé 10 pont
1.
- A genetikai diverzitás egy adott populációban jelenlévő allélok számával és azok gyakoriságával / az allélösszetétel változatosságával jellemezhető. 1 pont
- A fajdiverzitás azt írja le, hogy egy adott életközösségben hány faj él, és a jelenlévő fajoknak mekkora a relatív gyakorisága (egyedszámarányosan vizsgálja a fajszámot). 1 pont
- Az ökológiai diverzitás megmutatja az adott életközösségben fellelhető ökológiai szerepek / populációs kapcsolatok sokszínűségét / összetettségét. 1 pont
- A havasi kecske populációi esetén a genetikai diverzitás értelmezhető csak, melynek értéke csökken a lavinák miatti egyedpusztulás miatt. 1 pont
Másként megfogalmazott definíciók is elfogadhatók.
2.
- Az A) szövegrészlet alapján: fajok visszatelepítésével / mesterségesen létrehozott föld alatti üregekkel / a talaj tápanyagtartalmának helyreállításával segíthető a helyreállás. 1 pont
- A B) szövegrészlet alapján: akár helybeni védelemmel, például lavina elleni védettséget biztosító mesterséges búvóhelyek létrehozásával / akár egyedek védett helyekre, rezervátumokba telepítésével megvédhető a populáció a kihalástól. 1 pont
Bármely más helyes természetvédelmi módszer elfogadható.
3.
- A regeneráció során a K-stratégisták aránya növekszik, mivel a környezethez stabil létszámban alkalmazkodó populációk jönnek létre a szukcesszió során (a klimaxtársulás felé haladva). 1 pont
- A biomassza értéke szintén növekvő tendenciát mutat, mivel egyre nő az egyedszám (ezáltal az adott területen jelenlévő élővilág össztömege is). 1 pont
- A populációs kölcsönhatások száma szintén növekszik a szukcesszió előrehaladtával, mivel a megjelenő új fajok miatt egyre komplexebb kapcsolatrendszer jön létre. 1 pont
- A gyomnövényfajok száma viszont csökkenni fog, ugyanis a gyomok rosszul viselik a kompetíciót (a náluk erősebb versengők kiszorítják őket a területről). 1 pont
Más helyes indoklás, érvelés is elfogadható.
2025 május
XI. B. Hegységeink természetvédelme – esszé
Esszéjében összefüggő és logikus mondatokban fejtse ki válaszát, kitérve a következő szempontokra!
1. Említsen a hegyvidéki élőhelyeket átalakító, befolyásoló természetvédelmi szempontból káros vagy előnyös emberi hatások közül négyet, és kapcsolja össze ezeket az ivóvízellátással, a bányászattal, az árvízvédelemmel és a turizmussal! (8 pont)
2. Miért előnyösebb az élőhelyek jogi védelme az egyes fajok védelmével szemben? (1 pont)
3. A klímaváltozás mely lehetséges hatása és hogyan befolyásolja a medvefül kankalin elterjedését? (1 pont)
Javítási-értékelési útmutató
Hegységeink természetvédelme – esszé 10 pont
1. Négy fontos emberi hatás megfogalmazása, pl.:
- erdőirtások, fakitermelés: irtásrétek, kaszálók, települések megjelenése 1 pont
- erdészeti kezelés: kisebb diverzitású, egységesen kezelt állományok megjelenése 1 pont
- sziklagyepek helyén: települések, nyaralók, bányák külszíni fejtése, tájidegen fajok megjelenése 1 pont
- A bányászat (külszíni fejtések) főként szilagyepek helyén 1 pont
Más jó példák is elfogadhatók. A vizsgázó abban az esetben kapja meg az 1+1+1+1 pontot, ha mindegyik megadott szempontot figyelembe véve elemezte a problémát.
A vizsgázó által megadott négy (előnyös és/vagy káros) hatás elemzése a megadott szempontok szerint, pl.:
- Az erdős területek csökkenése fokozódó eróziót, árvízveszélyt jelent, / de az újraerdősítés / környezetkímélő gazdálkodás csökkenti ezt a veszélyt. 1 pont
- A települések, bányák, szeméttelepek stb. növelik a (karszt)vízszennyezés kockázatát, / de a csatornázás stb. csökkenti ezt a veszélyt. 1 pont
- A turizmus fokozhatja a környezeti terhelést (taposás, hegyi motorozás), / de a falusi turizmus előmozdíthatja a természetközeli gazdálkodást. 1 pont
- A bányászat csökkenti a fajok termőterületét / degradálja élőhelyét 1 pont
Négy összefüggés elemzése, mely a megadott témákat érinti.
2. Az élőhelyek / területek védelme jobban biztosítja az egyes fajok fennmaradását, mert a létükhöz szükséges környezeti tényezőket / kölcsönhatásokat fenntartja. 1 pont
3. A felmelegedés / egyenetlenné váló csapadékeloszlás / csökkenő páratartalom veszélyeztetheti a hűvös klímát kedvelő (reliktum) fajok, így a medvefül kankalin fennmaradását is. 1 pont
2024 október
IX.B. Tűrőképesség és biológiai indikáció – esszé
Esszéjében az alábbi szempontokra térjen ki!
1. Határozza meg a tűrőképesség fogalmát! (2 pont)
2. Egy tűrőképességi görbe rajzát elemezve adja meg annak lehetséges dimenzióit és jellemző szakaszait! (5 pont)
3. Értelmezze az indikátor szervezet fogalmát, hozzon példát indikációra, megadva azt, hogy a vizsgált példában mi, milyen módon s mit indikál! (3 pont)
Javítási-értékelési útmutató
Tűrőképesség és biológiai indikáció - esszé 10 pont
1.
- Tűrőképesség a környezeti tényezők értékeinek azon tartománya, 1 pont
- amelyen belül az adott faj populációja előfordulhat / amit a faj populációja elvisel 1 pont
2.
Egy toleranciagörbe felrajzolása után annak jellemzése:
- Az x / vízszintes tengelyen a vizsgált környezeti tényező értékeit, pl. hőmérséklet, páratartalom stb. (Bármely jó példa) 1 pont
- Az y / függőleges tengelyen a faj életképességét jellemző valamilyen mennyiséget pl. szaporodóképességet / egyedszámot / növekedési ütemet stb. tüntetjük fel. 1 pont
(Bármely jó példa)
- - minimum: éppen elviselhető alsó határ 1 pont
- - maximum: éppen elviselhető felső határ 1 pont
- - optimum: a legkedvezőbb értéktartomány. 1 pont
3.
- Indikátor szervezet: élőlény, amely jelenlétével vagy hiányával / egyedszámával / viselkedésével / gyarapodásával jelzi a (élettelen) környezet valamely tényezőjének értéktartományát. 1 pont
- Az indikátor fajok jellemzően szűktűrésűek (egy vagy néhány tényezővel szemben) 1 pont
- Pl. bizonyos zuzmók a kén-dioxid szennyezésre, ebben az esetben a zuzmók hiánya (indikáció) jelzi a kén-dioxid magas szintjét, 1 pont
Más példa is elfogadható, ha egyértelmű, hogy mely faj mi módon és mit indikál.
2022 október
IX.B. Természetvédelem – esszé 10 pont
1. Értelmezze a kihalási küszöb fogalmát, kapcsolatát a genetikai sodródással és a beltenyészet következtében föllépő leromlással! Magyarázza el, hogy mely természetvédelmi intézkedésekkel csökkenthető ennek veszélye! (4 pont)
2. Hozzon két példát a természetvédelem jogi lehetőségeire a fajok védelme érdekében, valamint két példát az aktív természetvédelem fajmegőrző szerepére! (4 pont)
3. Mutassa be egy példán keresztül a tájidegen fajok behurcolásának, bejutásának veszélyeit! (2 pont)
Javítási-értékelési útmutató
Természetvédelem – esszé 10 pont
1.
- A kihalási küszöb alatti populációlétszám nem teszi lehetővé a faj tartós fennmaradását (az adott területen). 1 pont
- Oka: a véletlen változások, így a genetikai sodródás változatosságot csökkentő hatása, 1 pont
- a kis populációkban föllépő beltenyészet miatt nő a recesszíven öröklődő betegségek / rendellenességek megjelenésének kockázata. 1 pont
- Természetvédelmi vonatkozások: megfelelő nagyságú terület / megfelelő élőhelyi változatosság stb. fenntartása ahhoz, hogy a védendő faj létszáma ne essen a kihalási küszöb alá. Más konkrét példa, pl. ökológiai folyosók fenntartása is jó válasz. 1 pont
2.
- A védett fajok gyűjtése / vadászata tilos, 1 pont
- a belőle készült termékek kereskedelme is tiltott: 1 pont
- valamely példa említése, pl: szőrmék, elefántcsont, kaktuszgyűjtés stb. 1 pont
- Aktív védelem: pl. sérült példányok gyógyítása / tenyésztés-szabadon engedés / magbankok / invazív, behurcolt fajok visszaszorítása. 1 pont
Más faj és a védelem másféle módjának bemutatása is elfogadható.
3.
- A behurcolt fajok kiszoríthatják az őshonosakat / betegségeket terjeszthetnek / mezőgazdasági károkat okozhatnak 1 pont
- Valamely példa említése, pl: parlagfű – allergia / tigrisszúnyog – vírusos betegségek. Más veszélyforrás és példafaj említése is megfelelő. 1 pont
2022 május
X.B. Erdők – esszé 10 pont
Leírásában hasonlítsa össze a cseres-tölgyes és a bükkös életközösségeket az alábbi szempontok alapján:
1. Zonális elhelyezkedésük, a különbség oka. A bükkösök visszaszorulásának éghajlati magyarázata. (4 pont)
2. Szerkezetük: gyepszint, különbségek a tavaszi és a nyári aszpektusokban, a változás oka; a cserjeszint jellemzői, magyarázata, lombkoronaszint. (6 pont)
Javítási-értékelési útmutató
Erdők – esszé 10 pont
1. Zonális elhelyezkedés:
- bükkös: 600 m felett 1 pont
- cseres-tölgyes: 400m alatt 1 pont
- A különbség oka: a bükk alacsonyabb átlaghőmérsékletet / több csapadékot igényel, mint a csertölgy. 1 pont
- Visszaszorulásuk magyarázata: csökkenő csapadékátlag / növekvő átlaghőmérséklet 1 pont
2. Az erdő szerkezete
- bükkös gyepszint: kora tavasszal nyíló hagymások, gumósok/ koratavaszi geofita aszpektus 1 pont
- Nyáron szegényes 1 pont
- a lombkoronaszint tavasz végi záródása miatt. 1 pont
- cserjeszint, bükkös: alig/csak a fák magoncai/ fiatal fák 1 pont
- cseres-tölgyes: egész évben gazdag gyep- és cserjeszint 1 pont
- a laza lombkorona / jó megvilágítás miatt. 1 pont
2021 október
IX.B. Életmódváltás – esszé 10 pont
Írjon esszét a klímaváltozás okairól az alábbi szempontok alapján:
1. Mit értünk fosszilis és megújuló energiahordozó alatt? Írjon példákat is! (3 pont)
2. Az amazóniai erdőégetés miért és hogyan befolyásolja az üvegházhatást? (2 pont)
3. Írjon két példát a biomassza energetikai felhasználására, illetve egy-egy érvet használata ellen és mellett! (3 pont)
4. Adjon egy-egy javaslatot a városi, illetve a távolsági közlekedés káros hatásának csökkentésére! (2 pont)
Javítási-értékelési útmutató
Életmódváltás – esszé 10 pont
1.
- A fosszilis energiahordozó hosszú idő alatt kialakult biológiai eredetű, széntartalmú energiahordozó 1 pont
- Megújuló: fogyásának és keletkezésének sebessége kb. azonos 1 pont
- Fosszilis: kőszén / kőolaj / földgáz, megújuló: víz / nap / szél. Bármely két jó példa 1 pont
2.
- Az égés során nő a levegő CO2-tartalma 1 pont
- A termőterület csökkenése / erózió miatti CO2-szint növekedés. / csökken a CO2 megkötése. 1 pont
3.
- Biodízel / bioetanol / fatüzelés – 2 példa 1 pont
- Ellene: az égési folyamatok során ugyanannyi káros anyag keletkezik, mint más égési folyamatoknál / termőterületet vesz el az előállítása / csökkentheti a biodiverzitást / előállítása is energiaigényes/ Más jó érv is elfogadható. 1 pont
- Mellette: Megújuló energiaforrás. / Nem növeli érdemben a fosszilis eredetű CO2 mennyiségét. 1 pont
4.
- Városi közlekedés: közösségi közlekedés / kerékpár / gyalogos közlekedés / forgalomszervezési megoldások / Más jó megoldás is elfogadható. 1 pont
- Távolsági közlekedés: Vasút használata közút helyett / kevesebb utazás / Más jó megoldás is elfogadható. 1 pont
2021 május
IX.B. A biodiverzitás szintjei - esszé 10 pont
Mutassa be, milyen szinteken értelmezhető a biodiverzitás és mi a megőrzésének jelentősége!
Esszéjében az alábbi szempontokat érintse:
1. Magyarázza meg, hogy a populációk szintjén értelmezett (faj-egyed) biodiverzitás értékének megadásához mit kell figyelembe venni! Nevezze meg az életközösség egy olyan tulajdonságát, mely összefügghet a biodiverzitás értékével, és magyarázza a kapcsolatot! (3 pont)
2. Magyarázza meg, hogy a genetikai diverzitás értelmezéséhez mit kell figyelembe venni!
Értelmezze, hogy ennek az alkalmazkodás folyamatában mi a jelentősége! Értelmezze a sodródás fogalmát és következményét ebből a szempontból! (5 pont)
3. Magyarázza meg, hogy a biodiverzitás védelme mi okból jelent mást, többet, mint az egyes fajok védelme! (2 pont)
Javítási-értékelési útmutató
A biodiverzitás szintjei – esszé 10 pont
1.
- A faj / faj-egyed diverzitást az egy területen élő fajok száma 1 pont
- és azok egyedszám-aránya / eloszlása szabja meg. 1 pont
- A faj / (egyed) diverzitás összefügg a társulás stabilitásával / biológiai produkciójával / a populációs kölcsönhatások sokféleségével / táplálkozási hálózatok tagjainak számával, mert fajgazdagabb / egyenletesebb eloszlású életközösségben többféle kapcsolat valósulhat meg. Más tulajdonság is elfogadható, ésszerű indoklással. 1 pont
2.
- A genetikai diverzitás a populációban előforduló allélok számától 1 pont
- és gyakoriságuk arányától / eloszlásuktól függ. 1 pont
- Magas genetikai diverzitás esetén a populáció alkalmazkodóképesebb, mert lehetőséget ad a szelekcióra / a populáció tagjainak együttműködésére. 1 pont
- A sodródás allélok véletlenszerű kiesését okozhatja, 1 pont
- ezért a genetikai diverzitást csökkenti. (palacknyak- / üvegnyak-hatás) 1 pont
3.
- A diverzitást a fajok kölcsönhatása tartja fenn. 1 pont
- ezért megőrzése az egyes fajok fennmaradást biztosítja (pl. táplálkozási hálózatok / szimbiózisok). 1pont
Másként is megfogalmazható. A megfogalmazás utaljon a társulás fajainak kölcsönös függésére.
2020 október
IX.B. Ökológiai kölcsönhatások – esszé 10 pont
Mutassa be az élővilág nagy országainak néhány képviselője közt fellépő populációs kölcsönhatásokat! A következőkre térjen ki:
1. Az antibiózis és az antibiotikumok fogalma és hasznosítása. (3 pont)
2. Az ember és a szervezetében élő baktériumok közötti szimbiózis, asztalközösség (kommenzalizmus) és parazitizmus (a baktériumcsoportokat anyagcseréjük, illetve a szervezetben betöltött szerepük vagy hatásuk alapján nevezze meg). (3 pont)
3. A szintezettség és az aszpektusok soksejtű növények természetes életközösségében (egy jellemző életközösség megnevezésével, a jelenség meghatározása és bemutatása példákon keresztül) (4 pont)
Javítási-értékelési útmutató
Ökológiai kölcsönhatások 10 pont
1.
- Az egyik fajnak közömbös, a másiknak előnytelen kölcsönhatás 1 pont
- Az antibiotikumok (többnyire) gombák által termelt, baktériumok szaporodását gátló anyagok 1 pont
- Felhasználásuk: baktériumos eredetű betegségek kezelésére. 1 pont
2.
- A vastagbélben élő baktériumok egy része szimbionta, mert vitaminokat / átvivőanyagokat termel / mert gátolja más baktériumok vagy gombák elszaporodását, 1 pont
- más részük kommenzalista, mert a vastagbélbe került emésztetlen szerves anyagok lebontását végzi, 1 pont
- bekerülhetnek szervezetünkbe élősködő / betegségokozó fajok is (bármely baktériumos eredetű megbetegedés megnevezése) 1 pont
Más helyszín és kölcsönhatás is leírható, pl. a bőrön vagy a hüvelyben élő baktériumok szerepe.
3.
- Szintezettség: a társulások vertikális szerkezete. Például a gyertyános-tölgyesek lombkorona- és mohaszintjének elkülönülése, 1 pont
- az eltérő fényigényű / páraigényű fajok térbeli elkülönülése miatt. 1 pont
- Aszpektusok: az életközösség évszakoktól függő változásai, például a bükkösök tavaszi / nyári / téli aszpektusa, 1 pont
- tavaszi aszpektus: a lombokorona záródása előtt virágzó lágyszárúak tömeges megjelenése a megvilágítás évszakos változásával magyarázható. 1 pont
Más példák és ezek magyarázata is elfogadható.
2020 május
IX.B. Az élő bolygó – esszé 10 pont
Fogalmazza meg James Lovelock Gaia-elméletének lényegét! Esszéjében a következőkre térjen ki!
1. Fogalmazza meg Gaia szabályozottságának lényegét és funkcióját az elmélet alapján! (2 pont)
2. Írjon példákat arra, hogy az élővilág hogyan hat a környezeti tényezőkre az alábbi esetekben: (4 pont)
- a vízgyűjtő területen fekvő erdők hatása a folyók vízjárására (az erdőtlen területekhez képest),
- pillangósvirágú növények hatása a talaj termőképességére,
- felnövekvő erdőrészletek hatása a légkör pára- és oxigéntartalmára.
3. Mutassa be, hogy milyen emberi tevékenység okozhatja a talajvíz nitrátosodását és a csapadék savasodását! Mutassa be ezek hatását biológiai rendszerekre, illetve az ember egészségére! (4 pont)
Javítási-értékelési útmutató
Az élő bolygó – esszé 10 pont
1.
- A Földön zajló (globális) anyagáramlási folyamatok szabályozottságuk miatt / visszacsatolások révén sok anyag koncentrációját határértékek között tartják, 1 pont
- ennek funkciója a földi élet tartós fenntartásának biztosítása. 1 pont
Másképp is megfogalmazható. A bemutatás jelezze a szabályozottság lényegét és funkcióját.
2.
- Az ilyen erdők egyenletesebbé teszik a folyók vízjárását, 1 pont
- A pillangósvirágúak / gyökérgümőiben élő nitrogénfixáló baktériumok segítségével növelik a talaj termőképességét, 1 pont
- a növekvő fák oxigént termelnek, 1 pont
- és párássá teszik a levegőt. 1 pont
3.
- A (túlzott) (mű)trágyázás / csatornázatlanság növeli a talajvíz nitráttartalmát, 1 pont
- ami csecsemőkre súlyos veszélyt jelent / methemoglobinémia / redukálódva rákkeltő nitritekké alakulhat (bármelyik veszélyes hatás említése) 1 pont
- A fosszilis energiahordozók (főként: kőszén) égetése savas esőket okoz / csökkenti a csapadék pH-ját, 1 pont
- ami (közvetve vagy közvetlenül) erdőpusztulást okozhat. 1 pont
Más, savasodással vagy növekvő nitráttartalommal összefüggő hatás (pl. eutrofizáció) és ezek lehetséges biológiai következményeinek megemlítése is elfogadható.
2019 október
IX.B. Gazdálkodás a nitrogénnel - esszé 10 pont
Esszéjében foglalja össze a nitrogén körforgásával kapcsolatos legfontosabb környezeti és gazdálkodási kérdéseket! Az alábbiakra térjen ki!
1. Mely ionok formájában veszi fel a legtöbb növény a nitrogént? (1 pont)
2. Milyen természetes és mesterséges folyamatok pótolhatják a mezőgazdasági talaj elvesztett nitrogéntartalmát? Mit jelent a minimumelv (Liebig-elv), és milyen gazdálkodási szabály fogalmazható meg ennek figyelembevételével? (4 pont)
3. Milyen következményekkel jár, ha az ivóvízbe sok nitrogéntartalmú ion kerül? (3 pont)
4. Milyen kémiai-biológiai problémákat okoz, ha a levegőbe sok nitrogén-oxid kerül? (2 pont)
Javítási-értékelési útmutató
Gazdálkodás a nitrogénnel - esszé 10 pont
1.
- Nitrát /NO3 – ionok formájában. (Elfogadható az ammónium-ion is.) 1 pont
2.
- Természetes úton a nitrogéngyűjtő baktériumok / a villámlás során keletkező nitrogénvegyületek. 1 pont
- Mesterségesen: trágyázás / műtrágyázás útján. 1 pont
- A minimum-elv szerint csak a legkisebb koncentrációban hozzáférhető ion arányában képes hasznosítani a növény a többit, 1 pont
- ezért csak annyi nitrogén(mű)trágyát szabad használni, amennyinek a hasznosítását a többi felvehető ion lehetővé teszi. 1 pont
3.
- Az ammónium-ion mérgező / undort keltő, / a nitrit-ion rákkeltő/ egészségkárosító. / A nitrát-ion nitrit-ionná redukálódhat. Bármelyik. 1 pont
- A nitrát-ionok csecsemőkben fulladást okoznak / methemoglobinémia, mert a (csecsemőkori) hemoglobinhoz kapcsolódva gátolják annak oxigénszállítását. 1+1 = 2 pont
4.
- A nitrogén-oxidok savas esőt okoznak, / mely növényzetpusztulást / korróziót okoz.
- A (fotokémiai) szmog / füstköd alkotóiként egészségkárosítók. 1+1 = 2 pont
2017 október
X.B. Veszélyforrások - esszé 10 pont
Foglalja össze a hazai sziklagyepeinket fenyegető veszélyforrásokat, a védelem lehetőségeit!
Az alábbiakra térjen ki:
1. Mely nemzeti parkjaink vagy hegységeink területén találhatunk sziklagyepeket (egy példa), természetvédelmi jelentőségük indoklása. (2 pont)
2. A tájidegen fajok, például feketefenyő telepítésének lehetséges hatása, a taposás, útépítések, külszíni bányászat következménye, a természetesnél sűrűbb vadállomány fenntartása.(4 pont)
3. A több kisebb védett területre szakítás következtében fellépő beltenyésztés és sodródás (genetikai) lényege, oka, ezek lehetséges hatása a populációk túlélésére. (4 pont)
Javítási-értékelési útmutató
Veszélyforrások – esszé 10 pont
1.
- Sziklagyepek a Bükki / Aggteleki / Duna-Ipoly / Balaton-felvidéki nemzeti parkok /Dunántúli- és Északi-középhegység / Mecsek területén (bármelyik példa jó) fordulnak elő. 1 pont
- Meleg- és hidegkedvelő maradványfajok / bennszülött növények / életközösségek lelőhelyei. (Másképpen is megfogalmazható.) 1 pont
2.
- A feketefenyő árnyékolása / lassan bomló avarrétege miatt kiszorítja az itt őshonos fajokat/ egyéb fajok kompetíciója hasonló hatású lehet 1 pont
- a taposás eróziót / degradációt / gyomosodást indít meg, 1 pont
- a külszíni bányászat csökkenti a termőterületet, 1 pont
- a túltartott vadállomány túrásával / rágásával károsít. 1 pont
3.
- A beltenyésztés a rokon egyedek egymás közti szaporodása, 1 pont
- ami az öröklődő recesszív rendellenességek megjelenésének kockázatát növeli, 1 pont
- A sodródás a kis populációk genetikai változatosságát véletlenszerűen csökkenti, 1 pont
- mindkettő csökkenti a populációk túlélési esélyét. 1 pont
2017 május
IX.B. Az erdő mintázatai – esszé 10 pont
Mutassa be egy hazai erdő tér-és időbeli mintázatait a következő példák segítségével:
1. Mik az aszpektusok, az aszpektusváltások oka és hatásai két példán bemutatva. (4 pont)
2. A szintezettség két lehetséges okának megnevezése egy erdőn belül. (2 pont)
3. A talaj humusztartalmának eredete, a növények életében játszott szerepe, a lebontásában részt vevő élőlények és szerepük. (4 pont)
Javítási-értékelési útmutató
Az erdő mintázatai – esszé 10 pont
1.
- Az aszpektusok a társulás ciklikusan / rendszeresen visszatérő állapotai, 1 pont
- a váltások fő oka az évszakok váltakozása. 1 pont
- Például ősszel a lombhullató fák leveleinek lehullása, 1 pont
- tavasszal a lombkorona záródása előtti gumós lágyszárúak virágzása (kora tavaszi aszpektus pl. a gyertyános-tölgyesekben). 1 pont
(Bármely két jó példa alkalmas.)
2.
- A szintezettség a fény / a páratartalom / az ökológiai kölcsönhatásban részt vevő partnerek / a szél / a hőmérséklet függőleges irányú egyenlőtlen eloszlásának következménye. 2 pont
(Bármely két jó környezeti tényező megnevezése.)
3.
- A humusz az elpusztult élőlények/maradványok szerves anyagaiból képződik,1 pont
- szemcséi / részecskéi felszínén víz és ásványi sók kötődnek meg, / lebontása után tápanyagforrás 1 pont
- a talajt morzsalékossá, levegőssé teszi. 1 pont
- A lebontók, pl. gombák, baktériumok alakítják vissza szervetlen anyaggá. 1 pont
NÖVÉNYTAN
2024 május
IX.B. A zárvatermők szaporodása – esszé 10 pont
Mutassa be a zárvatermők szaporodását az alábbi szempontok alapján!
1. Az ivaros és az ivartalan szaporítás lényege, lehetséges célja és az ivartalan szaporítás két módjának megnevezése. (3 pont)
2. A termés kialakulása az ivarsejtek kialakulásától kezdődően. A virág és a termés részeinek kapcsolata. (7 pont)
Javítási-értékelési útmutató
A zárvatermők szaporodása – esszé 10 pont
1.
- Az ivaros szaporítás lényege a genetikai változatosság biztosítása / két egyed genetikai állományának keveredése / nemesítés, 1 pont
- míg az ivartalan szaporítás célja, hogy a kiválasztott génállományú növényből minél többet hozzunk létre / pl. hibridek esetében az adott jelleg ne változzon. 1 pont
- két ivartalan szaporítási mód említése, pl. tőosztás / bujtás / szemzés / dugványozás / oltás 1 pont
2.
- A megtermékenyítés során az egyik hímivarsejt a petesejttel egyesülve zigótát, 1 pont
- majd csíranövényt / embriót alakít ki. 1 pont
- A másik hímivarsejt a központi sejttel egyesülve 1 pont
- a mag táplálószövetét hozza létre. 1 pont
- Ezzel párhuzamosan a magkezdemény burkából létrejön a maghéj, 1 pont
- a magkezdeményből a mag, 1 pont
- a magházból / termőből pedig termés lesz. 1 pont
2023 október
IX.B. A zárvatermők ivaros szaporodása – esszé 10 pont
Esszéjében ismertesse a megtermékenyítésre kész embriózsák felépítését és a kettős megtermékenyítés folyamatát az alábbi szempontok alapján!
1. Mely sejtek találhatók a megtermékenyítésre váró embriózsákban (nevük, számuk, ploidiatípusuk- azaz melyek a haploid, diploid és triploid sejtek)?
2. Hogyan játszódik le a kettős megtermékenyítés (a lépések sorrendje, eredménye, milyen ploidiatípusú sejte(ke)t eredményeznek)?
Javítási-értékelési útmutató
A zárvatermők ivaros szaporodása – esszé 10 pont
1.
- három ellenlábas sejt, 1 pont
- két segítő (kísérő) sejt 1 pont
- egy petesejt 1 pont
- ezek haploidok 1 pont
- egy központi sejt 1 pont
- ez diploid / két haploid sejt összeolvadásából jött létre 1 pont
2.
- vegetatív sejt – pollentömlőt hajt 1 pont
- generatív sejt (számtartó osztódás) – két hímivarsejt alakul ki 1 pont
- egyik hímivarsejt összeolvad a petesejttel: diploid zigóta 1 pont
- másik hímivarsejt összeolvad a központi sejttel: triploid sejt 1 pont
2023 május
IX.B. Megvilágítás és virágzás – esszé (10 pont)
A mikulásvirág virágoztatása sok esetben nehézségbe ütközik hazánkban, mivel virágzását befolyásolja a napi megvilágítás hossza. Mutassa be, hogyan befolyásolja a nappalhosszúság a növények virágzását. Esszéjében az alábbi szempontokra térjen ki!
1. A rövid- és a hosszúnappalos növény fogalma, származása. Magyarázza a különbséget a származási hely földrajzi jellemzőivel! (5 pont)
2. Hazánkban melyik évszakban jellemző a rövid- és a hosszúnappalos fajok virágzása? (2 pont)
3. Melyik kategóriába tartozik a mikulásvirág? Többnyire miért marad el az ablakpárkányon nevelt, szobai világításnak is kitett példányok virágzása? Javasoljon egy lehetséges módszert, amivel el lehet érni, hogy a mikulásvirág nálunk is virágozzon! (3 pont)
Javítási-értékelési útmutató
Megvilágítás és virágzás – esszé 10 pont
1.
- A rövidnappalos növények kb. azonos hosszúságú nappal és éjszaka / 10-12 óra megvilágítást követő min. 12 órás folyamatos sötétperiódus esetén virágoznak. 1 pont
- A hosszúnappalos növények az éjszaka tartamát meghaladó megvilágítás / 14-16 óra megvilágítás(t követő 8-10 órás sötétperiódus) esetén virágoznak. 1 pont
- A rövidnappalos növények trópusi / szubtrópusi területekről származnak, 1 pont
- a hosszúnappalos növények mérsékelt (vagy hideg) övi területekről származnak. 1 pont
- A különbség oka, hogy a trópusokon nincs jelentős évszaktól függő eltérés a nappalok és éjszakák hossza között, a mérsékelt övben a hosszú / hosszabbodó nappalok / rövidülő éjszakák a nyár közeledtét jelzik. 1 pont
2.
- A rövidnappalos növények hazánkban nyár végén / ősszel / koratavasszal virágoznak, 1 pont
- a hosszúnappalos növények hazánkban tavasszal / nyáron virágoznak. 1 pont
3.
- A mikulásvirág rövidnappalos növény, 1 pont
- virágzásának beindítását a mesterséges fény is gátolja. 1 pont
- Virágzásához hosszabb időre / legalább 12-14 órára teljes sötétségbe kell tenni a növényt. 1 pont
Más hasonló, a tartós sötétségre utaló válasz is elfogadható.
2019 május
IX.B. Folyadékáramlás az erdőkben – esszé 10 pont
Mutassa be a folyadékáramlás okát és következményeit a fákban, illetve hazai erdeinkben!
Az alábbiakra térjen ki:
1. Mely tényezők biztosítják a víz áramlását a gyökértől a levelek felé? Mely szövetelemekben zajlik ez a folyamat? (4 pont)
2. Milyen következményekkel járhat a (már megindult anyagcseréjű) fatörzsben a hirtelen tavaszi fagy? (1 pont)
3. Hogyan befolyásolja az erdei életközösség életműködéseivel a folyók vízjárását, a talaj és a légkör összetételét? (5 pont)
Javítási-értékelési útmutató
Folyadékáramlás az erdőkben – esszé 10 pont
1.
- A víz a gyökérnyomás, 1 pont
- a kapillaritás / kohézió 1 pont
- és a levelek párologtatása (következtében fellépő szívóerő/ ozmózisnyomás) miatt áramlik 1 pont
- a farész vízszállító csövein / (részben vízszállító sejtjein) át. 1 pont
2.
- A hirtelen fagy a működő, vízzel telt szállítószövetet szétrepesztheti (fagykár).1 pont
3.
- A fák (párologtatásuk miatt) párássá, 1 pont
- fotoszintézisük miatt oxigéngazdaggá teszik a légkört, 1 pont
- csökkentik a szén-dioxid szintet / szennyező anyagot kötnek meg. 1 pont
- Az erdei életközösség a talajt tápanyagban / humuszban gazdaggá teszi / az eróziót gátolja 1 pont
- Emiatt a folyók vízjárását egyenletesebbé teszik / az árvízveszélyt csökkentik. 1 pont
2018 október
IX.B. A növények párologtatása – esszé 10 pont
Írjon esszét a növények párologtatásáról! Térjen ki a következőkre:
1. Nevezze meg a növényi párologtatás helyszínét (szerv és szövettani részlet) valamint mechanizmusát! (3 pont)
2. Írja le, hogyan segíti a párologtatás a vízáramlást a növényben! (3 pont)
3. Mely környezeti tényezők befolyásolják a párologtatás mértékét! Említsen kettőt! (2 pont)
4. Száraz élőhelyeken honos növények milyen módon csökkentik párologtatásukat? Írjon két példát a testszerveződésben megnyilvánuló alkalmazkodásra! (2 pont)
Javítási-értékelési útmutató
A növények párologtatása – esszé 10 pont
1.
- A párologtatás szerve a levél.
- A gázcserenyílásokon keresztül
- diffúzióval a levegőbe kerül a vízgőz. 3 pont
2.
- A párologtatás hozza létre a levelek szívóhatását
- a vízgőz távozása miatt a levél sejtjeiben nő a koncentráció / ozmózisnyomás
- így a párologtatás a folyadék áramlását biztosítja a nagyobb ozmózisnyomású / koncentrációjú hely felé 3 pont
3.
- Pl. hőmérséklet / szél / levegő páratartalma /a vízutánpótlás / talaj víztartalma (Bármilyen 2 jó megoldás esetén.) 2 pont
4.
- Pl. kis felületű levelek / besüllyedt gázcserenyílások / szőrözött levél / viasz (Bármilyen 2 jó megoldás esetén.) 2 pont
2018 május
IX.B. A szén-dioxid útja a növényekben – esszé 10 pont
A gulya szerves anyagai eredendően a puszta növényeinek szerves anyagaiból jönnek létre. Az is igaz, hogy a szarvasmarhák által leadott szén is visszakerülhet a körforgásba. Mutassa be, hogy milyen utat járnak be a gulya által kilélegzett szén-dioxid szénatomjai a növényi szervezetben! Esszéjében az alábbi gondolatokra térjen ki!
1. A biológia oxidáció mely folyamatai során keletkezik a szarvasmarhák és a növények sejtjeiben CO2? (2 pont)
2. Kövesse végig a CO2 szénatomjának útját a növényi szerves anyaggá alakulásig! Nevezze meg a felvétel mechanizmusát és helyét (szövettípus, és azon belül a pontos helyszín)! Írja le, mely biokémiai folyamat során hasznosul a szén-dioxid és mely anyagok biztosítják ehhez az energiát, illetve a folyamathoz szükséges hidrogént! (6 pont)
3. Írja le, hogy mely két poliszacharid keletkezik a növényekben a fotoszintézis végtermékéből! (2 pont)
Javítási-értékelési útmutató
A szén-dioxid útja a növényekben – esszé 10 pont
1.
- A szarvasmarhák / és a növények szervezetében / a piroszőlősav átalakulása / a glikolízis zárólépése során / 1 pont
- és a citromsavciklus folyamatában keletkezik CO2. 1pont
2.
- A légköri CO2 a hajtás bőrszövetének 1 pont
- gázcserenyílásain át 1 pont
- diffúzióval jut a növénybe. 1 pont
- A szén-dioxidot a fotoszintézis sötét szakaszában / Calvin-ciklusban 1 pont
- A NADPH(2) / hidrogénszállító molekula hidrogénjeivel 1 pont
- az ATP energiájának felhasználásával redukálja szőlőcukorrá / szerves anyaggá a növényi sejt. 1 pont
3.
- A glükózból keményítő 1 pont
- és cellulóz képződik. 1 pont
Emelt szintű jegyzetek
Emelt szintű jegyzetek megjelenítése / eltüntetése
Az alábbi emelt szintű jegyzetek teljes mértékben a 2020-as NAT által meghatározott, 2022 január 1-től alkalmazandó, 2024-től pedig kizárólagosan érvénybe lépő, jelenleg is aktuális érettségi követelményrendszer alapján készültek.
Minden jegyzet elején az adott fejezethez tartozó követelmények vannak feltüntetve, melyek egyben az adott fejezetek tartalomjegyzékei. A fehér betűs részek a középszintű, a sárga színnel kiemelt szövegrészek az emelt szintű követelményeket jelzik. Az emelt szintű követelmények magukba foglalják a középszintű ismeretanyagot is. Dőlt betűs részek középszinten a szóbeli B tételek lehetséges elvégzendő vizsgálatait, emelt szinten a szóbeli B tételek lehetséges problémafeladatait tüntetik fel.
A jegyzetekben a tananyag vastag betűs, ill. a kék kiemeléssel jelölt szövegrészei a követelmények által elvárt fontosabb fogalmakat, ismereteket tartalmazzák, ugyanakkor az apró, szürke betűs részek nem szükségesek, csupán kiegészítő jellegűek. A jegyzetek alatt megtalálhatók az adott fejezetekhez tartozó korábbi középszintű és emelt szintű érettségi feladatok tematikus gyűjteményei, melyek Gőz József tanár úr (BioTéka) nagyszerű összeállításai. A jegyzetekben, ill. a tematikus gyűjteményekben megtalálható érettségi feladatok az Oktatási Hivatal és annak jogelődjei szellemi tulajdonát képezik. Továbbá köszönettel tartozom fakultációs diákjaimnak, különösen Szepesi Máténak a hasznos észrevételekért.
Az egyes fejezetek folyamatosan frissülnek, érdemes a jegyzetek megnyitása előtt újratölteni az oldalt. Androidos készülékek használata esetén javasolt a Firefox böngésző használata.
Jó tanulást, sikeres felkészülést kívánok!
A jegyzetek a készítő nevének feltüntetése mellett nem kereskedelmi céllal szabadon másolhatók, nyomtathatók, terjeszthetők, előadhatók. A jegyzetek anyagának bármilyen további felhasználása kizárólag a készítő előzetes engedélyéhez kötött.
Tekintve, hogy az oldal minden tanulni vágyó diák számára költségmentesen elérhető, a jegyzetek anyagának bármilyen kereskedelmi célű felhasználásához semmilyen módon nem járulok hozzá.
Vizkievicz András
- A biológia vizsgatárgy 2024. május-júniusi vizsgaidőszaktól érvényes vizsgakövetelményei(2020-as NAT-ra épülő vizsgakövetelmények)
- Tájékoztató az emelt szintű biológia szóbeli vizsgához, az A) feladatok címei
1. BEVEZETÉS A BIOLÓGIÁBA - 1.1. A biológia tudománya
- 1.2. Fizikai, kémiai alapismeretek
2. EGYED ALATTI SZERVEZŐDÉSI SZINT - 2.1. Szervetlen és szerves alkotóelemek
- 2.3. Sejtalkotók az eukarióta sejtben
- 2.3.1. Eukarióta sejtalkotók, 2.3.2. Elhatárolás és összeköttetés, 2.3.3. Mozgás, 2.3.4. Anyagcsere, 2.3.5. Osztódás, 2.3.6. A sejtműködések szabályozása és a sejtek közötti kommunikáció
- Anyagcsere, sejtbiológia, molekuláris genetika fejezetek középszintű és emelt szintű érettségi feladatai
- Középszintű és emelt szintű érettségi feladatok megoldásai
3. AZ EGYED SZERVEZŐDÉSI SZINTJE - 3.1. Nem sejtes rendszerek
- 3.2. Sejtes rendszerek
- 3.3. Többsejtű eukarióták
- 3.4. Szövetek, szervek, szervrendszerek, testtájak
- 3.4.1. A növényvilág főbb csoportjai a szervi differenciálódás szempontjából, 3.4.2. A növények szövetei, szervei, életműködései
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 3.4.3. Az állatvilág főbb csoportjai a szervi differenciálódás szempontjából
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 3.4.4. Az állatok szövetei, szaporodása, viselkedése
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
4. AZ EMBERI SZERVEZET - 4.1. Homeosztázis, rendszerszemlélet
- 4.2. Kültakaró
- 4.3. A mozgás
- 4.4. A táplálkozás
- 4.5. A légzés
- 4.6. Az anyagszállítás
- 4.8. A szabályozás
- 4.8.1. Idegrendszer és érzékszervek, 4.8.1.1. Idegrendszer, 4.8.1.2. Sejtszintű folyamatok, 4.8.1.3. Szinapszis
- 4.8.1.4. Az idegrendszer általános jellemzése, 4.8.1.5. A gerincvelő
- 4.8.1.6. Az agy, 4.8.1.7. Testérző rendszerek, 4.8.1.8. Érzekelés
- 4.8.1.9. Látás
- 4.8.1.10. Hallás és egyensúlyérzés, 4.8.1.11. Kémiai érzékelés
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 4.8.1.12. Testmozgató rendszerek, 4.8.1.13. Vegetatív érző- és mozgatórendszerek
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 4.8.2. Az emberi magatartás biológiai-pszichológiai alapjai, 4.8.2.1.Kognitív folyamatok, 4.8.2.2. A magatartás elemei, 4.8.2.2.1. Öröklött elemek, 4.8.2.2.2. Tanult elemek, 4.8.2.3. Emlékezés, 4.8.2.4. Pszichés fejlődés, 4.8.3. Az idegrendszer egészségtana, 4.8.3.1. Drogok
- 4.8.4. A hormonrendszer, 4.8.4.1. Hormonális működések, 4.8.4.2. Belső elválasztású mirigyek, 4.8.4.3. A hormonrendszer egészségtana
- Hormonok összefoglaló táblázata
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 4.8.5. Az immunrendszer, 4.8.5.1. Immunitás, 4.8.5.2. Vércsoportok, 4.8.5.3. Az immunrendszer egészségtana
- 4.9. Szaporodás és egyedfejlődés
5. EGYED FELETTI SZERVEZŐDÉSI SZINTEK - 5.1. Populáció
- 5.2. Életközösségek
- 5.4. Ökoszisztéma
- 5.3. Bioszféra 5.5. Környezet- és természetvédelem 5.6. Fenntarthatóság
6. ÖRÖKLŐDÉS, VÁLTOZÉKONYSÁG, EVOLÚCIÓ - 6.1. Molekuláris genetika
- 6.2. Mendeli genetika
- 6.2.1. Minőségi jellegek, Mendel szabályai, mendeli genetika kiterjesztése, monogénes autoszomális recesszív (AR), autoszomális domináns (AD), X-hez kötött recesszív (XR), X-hez kötött domináns (XD) jellegek, letális allélok, génkölcsönhatások, extranukleáris öröklődés, kapcsolt öröklődés 6.2.2. Mennyiségi jellegek
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- Gyakorló feladatok I
- Gyakorló feladatok II
- 6.3. Evolúció
- 6.3.1. Evolúciós folyamatok, Darwin evolúciós elmélete, evolúciós változások mechanizmusai, Hardy-Weinberg modell 6.3.2. Fajképződés és az evolúció bizonyítékai
- Populációgenetika feladatok
- Középszintű és emelt szintű érettségi feladatok
- Középszintű és emelt szintű érettségi feladatok megoldásai
- 6.3.3. Biotechnológia, 6.3.4. Bioetika, 6.3.5. Bioinformatika
Kapcsolat
Üzenetküldés
Van bármilyen kérdésed vagy javaslatod számomra? Bátran küldd el
az alábbi űrlapon.